蘋果芽變品種珍富果實品質和花青苷合成基因表達分析
2024-04-30 14:57:10汪少麗車曉志石潔陳敏張紫然陳平張新忠劉保友
果樹學報 2024年3期
汪少麗 車曉志 石潔 陳敏 張紫然 陳平 張新忠 劉保友

摘? ? 要:【目的】測定2001富士及其芽變品種珍富之間果實品質及花青苷合成相關基因表達水平差異,明確珍富品種特性,為其示范推廣提供參考依據,也為研究果皮顏色芽變機制提供理論支撐。【方法】以芽變品種珍富及2001富士果實為試材,對其采收期及貯藏14 d后果實外觀、內在品質、花青苷含量和合成相關基因表達水平進行測定。【結果】與2001富士相比,芽變品種珍富果實采收日期可以提前6 d,盛花期至采收期提前6 d;采收期及貯藏14 d后,外在品質方面,珍富果皮亮度、飽和度、花青苷總量及3種不同花青苷的含量均顯著高于2001富士,但二者單果質量及果形指數無顯著差異;果實內在品質方面,采收期及貯藏14 d后,珍富果實的可溶性固形物、維生素C含量及固酸比都顯著高于2001富士,但珍富的可滴定酸含量顯著低于2001富士;二者果肉硬度、破裂力在采收期及貯藏期均無顯著差異;10個花青苷合成關鍵結構基因及5個轉錄因子類調控基因都有不同程度的提高,關鍵結構基因MdC4H、MdANS、MdUFGT及轉錄因子基因MdMYB10、MdMYB11、MdERF3表達量顯著提高。【結論】采收期及貯藏14 d后,紅色芽變品種珍富果皮亮度L*值及果皮顏色飽和度C*值均顯著高于2001富士;珍富果實可溶性固形物、維生素C含量及固酸比顯著提高,可滴定酸含量顯著降低;珍富果皮中花青苷總量及3種不同花青苷含量都顯著提高,花青苷合成相關結構基因和轉錄調控基因也顯著上調表達。
關鍵詞:蘋果;芽變品種;果實品質;花青苷;基因表達
中圖分類號:S661.1 文獻標志碼:A 文章編號:1009-9980(2024)03-0426-10
Analysis of fruit quality and expression of genes related to anthocyanins synthesis in apple bud sport Zhenfu
WANG Shaoli1, CHE Xiaozhi2#, SHI Jie1, CHEN Min1, ZHANG Ziran1, CHEN Ping1, ZHANG Xinzhong3*, LIU Baoyou1*
(1Yantai Academy of Agricultural Sciences, Yantai 265500, Shandong, China; 2Agricultural Comprehensive Service Center, Longwangzhuang Subdistrict Office, Laiyang 265500, Shandong, China; 3China Agricultural University, Beijing 100083, China)
Abstract: 【Objective】 Fuji 2001 apple has been widely planted since it was introduced into China from Japan in 1993. Zhenfu is a red bud sport of Fuji 2001, and it passed variety registration on August 24, 2021. The peel of Zhenfu shows more obvious red color than that of Fuji 2001. This mirable advantage in peel color has attracted the attention of apple growers. However, whether there are differences in fruit quality, anthocyanin compositions and contents, and expression of anthocyanin synthesis-related genes need to be further clarified. These results will provide reference for production extension and a research basis for disclosing the peel pigment mechanism of Zhenfu. 【Methods】 Traits of fruits, internal quality, composition of anthocyanin compounds and expression level of the genes related to pigment synthesis of Fuji 2001 apple and its bud sport Zhenfu fruits were measured and assessed. The maturity of apple was determined by the starch-iodine staining method. The vertical and horizontal diameters of each fruit were measured by the syntek electronic digital vernier caliper. The single fruit weight of each fruit was measured by an electronic balance. The flesh firmness and rupture force were measured by TMS-PRO texture analyzer. The content of soluble solids was measured by PR-101α refractometer, and peel brightness (L*) and saturation (C*) were measured by CR-400 colorimeter. Titratable acid was determined by acid-base titration. The contents of vitamin C and soluble solids were determined by 2, 6-dichloroindophenol titration and PAL-1 digital refractometer, respectively. Solid/acid ratio was expressed as the ratio of soluble solids to total acid. Peel sample of 1.00 g was grinded into homogenate in liquid nitrogen, dissolved in 5 mL of HCl-methanol (0.5∶99.5, v/v) solution, extracted for 24 h at 4 ℃ under dark conditions, centrifuged at 12 000 r·min-1 for 10 min at 4 ℃, and filtered through 0.22 μm organic phase membrane. 1.5 mL supernatant was transferred to an automatic injection bottle and detected by Waters HPLC high performance liquid chromatography to determine anthocyanin contents. The total RNA of fruit peel was extracted by trizol extraction kit. RNA was reverse transcribed into cDNA using the first-strand cDNA synthesis kit. The first-strand cDNA was applied to analyze gene expression levels by step-one fluorescence quantitative PCR. 【Results】 The fruit harvest date of Zhenfu was 6 days earlier than Fuji 2001. In terms of external qualities, both apple peels set red stripes, but Zhenfu peel showed more obvious red color. At the day of harvest and 14th day after storage, L* value of peel brightness and C* value of peel color saturation of Zhenfu were significantly higher than those of Fuji 2001. Three kinds of anthocyanin compounds, including cyanidin-3-galactoside, cyanidin-3-O-glucoside and anthocyanin rhamnoside, were detected in the peels of both Fuji 2001 and Zhenfu. Among them, the content of cyanidin-3-galactoside was the highest, and was the main anthocyanin component in the peel of Fuji 2001 and Zhenfu. At the day of harvest, the contents of cyanidin-3-galactoside, cyanidin-3-O-glucoside and anthocyanin rhamnoside in Zhenfu peel were 2.21, 1.98 and 1.60 times higher than those of Fuji 2001 peel. The total contents of anthocyanins in Zhenfu were 2.18 times higher than those of Fuji 2001. After 14-day storage, the contents of 3 kinds of anthocyanins and the total amount of anthocyanins in Zhenfu were still higher than those in Fuji 2001. There was no significant difference in fruit quality and fruit shape index between them. In terms of internal quality, the contents of soluble solids, vitamin C and solid acid of Zhenfu were significantly higher than those of Fuji 2001. The titratable acid content of Zhenfu was significantly lower than that of Fuji 2001. However, there were no significant differences in flesh firmness and rupture force between the two varieties at the day of harvest and 14th day after storage. Some key structural genes and positively regulating transcription factor genes in anthocyanin synthesis pathway showed higher expression levels in Zhenfu than those in Fuji 2001, especially MdC4H, MdANS, MdUFGT, MdMYB10, MdMYB11 and MdERF3. 【Conclusion】 At harvest and 14th day after storage, peel brightness L* value and peel color saturation C* value of Zhenfu were significantly higher than those of Fuji 2001. The titratable acid content of Zhenfu was significantly lower than that of Fuji 2001, and the soluble solids content, vitamin C content and solid/acid ratio were significantly higher than those of Fuji 2001. In addition, the total amount of anthocyanins in the peel of Zhenfu was significantly higher than that of Fuji 2001. The key structural genes in the anthocyanin synthesis pathway of Zhenfu and the related transcriptional regulatory factors were significantly up-regulated in the peel of Zhenfu.
Key words: Apple; Bud sport; Fruit quality; Anthocyanins; Gene expression
果實色澤是蘋果重要的外觀品質,極大程度地決定了果實的商品價值。蘋果果皮色澤與果皮花青苷含量密切相關,同時花青苷也是果實重要的功能營養成分,具有預防心腦血管疾病、抗氧化、延緩衰老等重要的保健功能[1-3]。因此,研究蘋果果皮色澤的遺傳特性及調控機制,改善果實色澤和提高果皮花青苷含量是蘋果育種及相關領域研究關注的熱點。
花青苷通過苯丙氨酸代謝途徑,在多種結構基因及轉錄因子類調控基因的參與下合成。結構基因主要包括PAL、CHS、CHI、F3H、DFR、LDOX、ANS和UFGT[4-5]。其中,CHS、CHI以及F3H為早期結構基因,而DFR、LDOX、ANS和UFGT為晚期結構基因,都在花青苷合成途徑中起正向調控作用[6-7]。花青苷的生物合成還受到MYB、bHLH和WD40蛋白復合體(MYB-bHLH-WD40)的調控,其中MYB轉錄因子發揮至關重要的作用[8-10]。MdMYB1、MdMYB10被認為是蘋果花青苷生物合成的正向調控因子,直接激活MdDFR和MdUF3GT基因的表達,促進花青苷的生物合成[11-13]。報道還發現,茉莉酸甲酯處理嘎拉蘋果幼苗后,MdMYB11基因表達量提高,MdMYB11基因轉入蘋果愈傷組織后,通過結合MdANS等結構基因啟動子調控花青苷的合成,積累大量的花青苷與原花青苷[14]。除了MYB之外,其他家族轉錄因子也參與了花青苷的代謝。MdbZIP44在激素ABA誘導下,與MdMYB1相互作用,增強了MdMYB1與下游靶基因的互作從而促進了花青苷的合成[15]。另有研究發現,在擬南芥及蘋果葉片中轉入MdERF3基因后,花青苷及原花青苷含量顯著提高[16]。
2001富士品種自1993年由日本引入中國以來,以其豐產、質優的顯著特點受到果農和消費者的普遍歡迎和認可[17]。珍富是2001富士的紅色芽變,發現于山東省棲霞市,并于2021年8月24日獲得品種登記證書。珍富與2001富士果皮顏色都為條紅類型,但珍富果皮表現出了更明顯的紅色,這種果皮色澤上的明顯優勢受到蘋果種植戶的重視,并在一定區域內得到了快速發展。然而珍富與2001富士相比,除了明顯的果皮顏色差異外,在果實品質、花青苷類物質組分及含量、花青苷合成相關基因的表達方面是否存在差異仍需進一步明確,為珍富蘋果的示范推廣提供數據支撐,也為進一步解析其果皮著色機制提供基礎。
1 材料和方法
1.1 材料
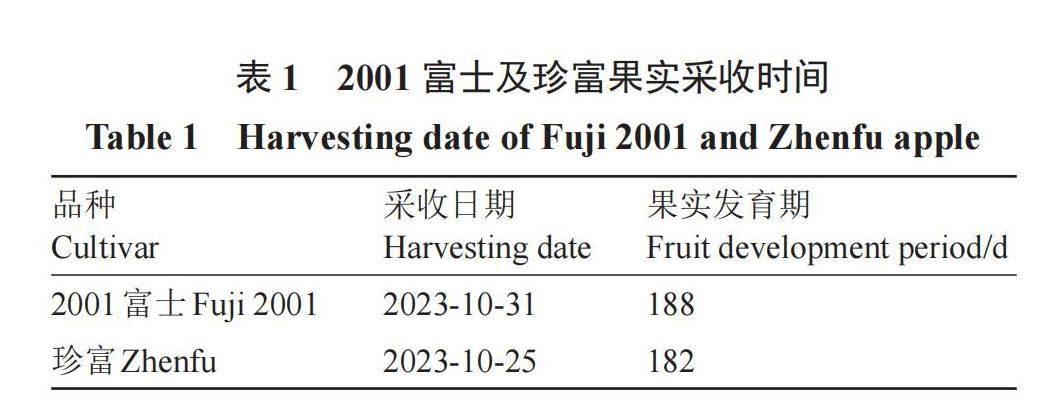
2001富士和珍富芽變蘋果果實試材均采自山東省棲霞市莊園街道謝家溝村,兩品種的砧木(八棱海棠)、栽培管理措施、果袋類型、套袋及摘袋時間均保持一致,2001富士及珍富果實采收時間見表1。
1.2 方法
1.2.1 果實采摘 選取長勢良好、樹齡一致的蘋果樹,在樹冠外圍和內膛不同方向均勻采收。每個品種選擇大小一致、無病蟲害及磕碰傷的果實,隨機分為3組,每組30個果實。第1組果實于采摘當天進行果實性狀相關指標測定;第2組果實在(25±1)℃、相對濕度85%~90%環境條件下貯藏14 d后再進行果實相關指標測定。第3組果實在田間將果皮用液氮速凍后裝入液氮罐中,于-80 ℃超低溫冰箱中保存,測定時液氮冷凍研磨成粉末,用于花青苷含量及基因表達分析。
1.2.2 果實性狀測定 采用淀粉-碘染色法[18]測定蘋果成熟度;采用syntek電子數顯游標卡尺測定每個果實的縱橫徑,計算為果形指數;日本島津TXB222L型電子天平測定每個果實單果質量;使用TMS-PRO質構儀(圓盤探頭直徑為75 mm)測定果實破裂力和硬度;使用PR-101α折光儀(日本ATAGO)測定可溶性固形物含量,采用CR-400色差計測定果皮顏色,所用光源為D65,分別測定果皮的亮度(L*)和飽和度(C*);可滴定酸含量采用酸堿滴定法測定,所用儀器為瑞士萬通808電位滴定儀;維生素C含量采用2,6-二氯靛酚滴定法測定,所用儀器為瑞士萬通808電位滴定儀;采用PAL-1數顯折光儀測定果實汁液中的可溶性固形物含量;固酸比以可溶性固形物含量和總酸含量的比例表示。以上每個指標測定30個果實。
1.2.3 花青苷提取及含量測定 取果皮樣品1.00 g,經液氮研磨成勻漿,溶解于5 mL的HCl-甲醇(0.5∶99.5,j)溶液中,4 ℃及黑暗條件下提取24 h,然后在4 ℃條件下,12 000 r·min-1離心10 min,再過0.22 ?m有機相濾膜,將1.5 mL上清液轉移至自動進樣瓶中,用美國Waters HPLC高效液相色譜儀檢測。檢測波長530 nm,柱溫35 ℃,流速1.0 mL·min-1,進樣5 ?L,梯度洗脫,溶液A(甲醇)和溶液B(10%甲酸水溶液)。洗脫條件如下:0 min,溶液A 17%,溶液B 83%;1 min,溶液A 17%,溶液B 83%;8 min,溶液A 35%,溶液B 65%;25 min,溶液A 37%,溶液B 63%;30 min,溶液A 17%,溶液B 83%。
1.2.4 RNA提取和cDNA第一條鏈合成 采用Trizol提取試劑盒提取試驗蘋果果皮的總RNA。以2種試材的RNA為模板,使用第一鏈cDNA合成試劑盒(RevertAid Premium Reverse Transcriptase)(Thermo Scientific? EP0733)將RNA反轉錄成cDNA,用于后續研究。
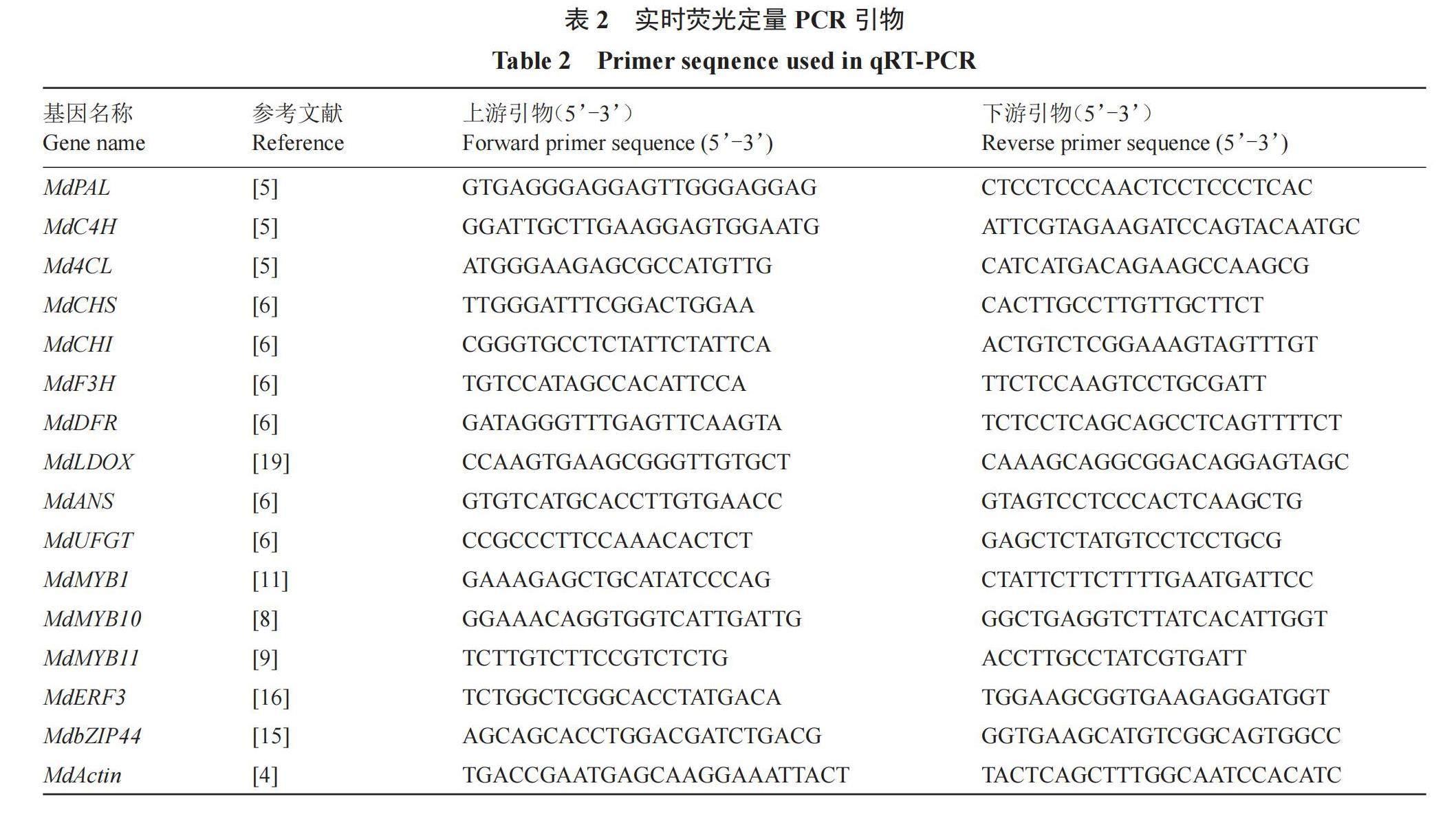
1.2.5 引物設計及qRT-PCR 采用Step-one型熒光定量PCR儀(ABI),反應體系為SYBR Green qPCR Master Mix 10 ?L,cDNA模板2 ?L,上下游引物各0.4 ?L,ddH2O 7.2 ?L;擴增程序為95 ℃預變性3 min;95 ℃變性7 s,57 ℃退火10 s,72 ℃延伸15 s,共40個循環。蘋果花青素合成相關基因MdPAL、MdC4H、Md4CL、MdCHS、MdCHI、MdF3H、MdDFR、MdLDOX、MdANS、MdUFGT、MdMYB1、MdMYB10、MdMYB11、MdERF3、MdbZIP44、MdActin的定量PCR引物序列見表2。
1.2.6 數據分析 采用Excel 2010進行數據處理,采用DPS 18.10進行差異顯著性分析,各圖表中不同小寫字母分別表示各處理在5%水平上的差異顯著性。
2 結果與分析
2.1 2001富士和珍富芽變蘋果果實成熟期及外觀比較
2001富士和珍富芽變蘋果均屬于晚熟品種,在所有栽培措施一致的情況下,果實完全成熟時確定為采收日期,與2001富士相比,珍富果實采收日期提前了6 d,這表明果實發育期縮短了6 d(表1)。2001富士和珍富芽變蘋果均為“條紅”型蘋果,但珍富果皮呈現更加明顯的紅色(圖1)。采收期,珍富果皮亮度L*值及果皮顏色飽和度C*值均顯著高于2001富士的果皮。貯藏14 d后,兩種蘋果果皮L*值及C*值均呈上升趨勢,證實果皮顏色能夠在貯藏過程中發生變化,且珍富果皮亮度L*值及果皮顏色飽和度C*值仍顯著高于2001富士,這表明貯藏期珍富的色澤特性優于2001富士(圖2)。2001富士和珍富果實的單果質量及縱橫徑比方面無顯著差異,二者均呈圓形或近圓形,縱橫徑比即果形指數分別為0.90、0.88(圖3)。
2.2 2001富士和珍富果皮花青苷類物質含量比較
蘋果果皮色澤與果皮花青苷含量密切相關,進一步測定果皮花青素含量發現,采收期在2001富士和珍富果皮中都檢測到了3種花青苷類物質,分別為矢車菊素-3-半乳糖苷、矢車菊素-3-O-葡萄糖苷及花色素鼠李糖苷。其中,矢車菊素-3-半乳糖苷含量最高,是2001富士和珍富果皮中主要的花青苷組成成分。珍富果皮中矢車菊素-3-半乳糖苷、矢車菊素-3-O-葡萄糖苷及花色素鼠李糖苷含量分別是2001富士果皮中的2.21倍、1.98倍、1.60倍,花青苷總量是2001富士果皮中的2.18倍。貯藏14 d后,2001富士和珍富果皮中花青苷含量均呈上升趨勢。珍富果皮中3種花青苷含量及花青苷總量仍高于2001富士(表3)。
2.3 2001富士和珍富果實內在品質比較
如圖4所示,采收期珍富果實的可溶性固形物含量在5%水平上顯著高于2001富士,而其可滴定酸含量也顯著低于2001富士;珍富果實的維生素C含量及固酸比分別是2001富士的1.21、1.20倍;果肉硬度、破裂力方面二者無顯著差異。貯藏14 d后,二者可溶性固形物含量分別降低8.06%、6.46%,但珍富果實的可溶性固形物含量顯著高于2001富士,且其可滴定酸含量仍顯著低于2001富士;二者維生素C含量都顯著降低,但珍富仍是2001富士的1.41倍;二者固酸比分別提高了6.66%、6.63%,珍富是2001富士的1.20倍;二者果肉硬度、破裂力均降低,且無顯著差異。
2.4 2001富士和珍富果皮花青苷類物質合成途徑相關基因表達量比較
花青苷合成途徑中的關鍵結構基因在2001富士和珍富中出現了不同的表達模式。其中,珍富果皮中MdPAL、Md4CL、MdCHS、MdCHI、MdF3H、MdDFR、MdLDOX表達量提高幅度較小,分別是2001富士的2.34、2.24、2.80、3.35、2.78、2.85、2.45倍,MdC4H、MdANS、MdUFGT表達量提高幅度更顯著,分別是2001富士的5.07倍、7.83倍、12.65倍(圖4)。
除了以上關鍵的結構基因外,轉錄因子也是影響花青苷合成的關鍵因素。因此,筆者挑選了部分已報道的正向調控花青苷合成的轉錄因子進行了表達量分析,發現MdMYB10、MdMYB1、MdMYB11、MdERF3、MdbZIP44轉錄因子基因表達量在珍富果皮中均顯著提高。其中,珍富果皮中MdMYB10、MdMYB11、MdERF3表達量提高更顯著,分別是2001富士的3.61倍、6.37倍、10.00倍(圖5)。
3 討 論
紅富士在我國蘋果產業獨占鰲頭(占比約70%),以紅富士為主的品種結構使我國成為世界上最大的蘋果生產國和消費國[20]。富士系蘋果是經芽變選種的育種手段培育的系列品種,芽變選種的突出特點是優中選優,經此育種手段選育的長富2號、富士3號、富士8號、龍富等富士系蘋果品種,占據蘋果市場的很大比例,一代又一代新的紅色優質芽變品種的出現有效推動了蘋果產業的發展[21],探究芽變品種果實品質的形成機制對改善蘋果品質、推動蘋果產業發展具有重要意義。
2001富士屬于紅富士的紅色和短枝芽變品種,其晚熟和耐貯這2個性狀沒有改變,滿足了我國的市場需求[22-23]。筆者在本研究中選擇的芽變品種珍富的果實采收日期及盛花期至采收期時間較2001富士均明顯縮短,證實珍富較2001富士提早成熟,而這種提早成熟可能與乙烯正調控因子MdERF3有關,據報道ERF家族轉錄因子在植物生長中起著重要作用,參與調節植物對激素、脅迫、果實成熟的反應并調控花青素合成[24]。在本研究中,珍富果實中MdERF3轉錄因子基因相較于2001富士上調表達10倍,因此筆者推測珍富的提早成熟可能與MdERF3基因的較高表達有關。
在內在品質方面,采收期珍富的可溶性固形物、維生素C含量及固酸比均顯著高于2001富士,而可滴定酸含量顯著低于2001富士,果肉硬度、破裂力均無顯著差異;在外在品質方面,珍富果皮亮度L*值及果皮顏色飽和度C*值均顯著高于2001富士的果皮,二者的單果質量、果形指數均無顯著差異。以上結果證實,采收期不論是內在還是外在品質珍富都要優于2001富士;且兩種蘋果分別貯藏14 d后,上述差異仍存在,但二者除了固酸比升高外,可溶性固形物含量、硬度等其他指標都有所降低,這也證實蘋果在采收后仍進行著一系列的生命活動,即蘋果的后熟軟化,蘋果作為典型的呼吸躍變型果實,其在后熟過程中呼吸作用強度驟然升高,生成大量乙烯,同時可溶性糖等營養物質消耗增多[25];貯藏中構成果實細胞壁的果膠質、纖維素等物質降解加速了細胞壁結構的松弛,也是貯藏期果實硬度下降的原因之一[26]。
果實色澤也是蘋果重要的外在商品性狀,它直接影響蘋果的市場競爭力[27-29]。蘋果果皮紅色主要由花青苷決定,花青苷在人類健康方面也扮演著重要的角色,具有抗氧化、提高記憶力、增進視力、預防肺部疾病和抗腫瘤等多種生理功能[3]。在本研究中,采收期及貯藏14 d時,2001富士和珍富果皮中都檢測到了3種花青苷類物質。其中,含量最高的為矢車菊素-3-半乳糖苷,該結果與其他報道中發現的蘋果果皮中含量最多的花青苷種類是矢車菊-3-半乳糖苷的結果一致[19,30]。
目前關于蘋果果皮花青苷合成基因在不同蘋果果實、不同環境中的表達模式的研究已見諸多報道[29]。筆者在本研究中選取的10個已報道與花青苷合成相關的結構基因,均在珍富果皮上調表達,這也證實上述結構基因的上調表達促進了其花青苷的合成積累,而這些結構基因的上調表達與MYB1、MYB10、MYB11等MYB類轉錄因子的調控相關[12-14]。另有研究證實,ERF家族轉錄因子也可以直接激活結構基因的啟動子調控花青素生物合成,例如MdERF109通過直接結合花青素結構基因MdCHS、MdUFGT以及調節基因MdbHLH3啟動子并激活其轉錄,促進花青素生物合成[31]。ERF家族轉錄因子不僅能夠直接激活結構基因的啟動子,還能夠與MYB類轉錄因子結合,通過MdERF3依賴的途徑促進乙烯的生物合成,從而增強乙烯途徑對激活MdMYB1自身轉錄活性的促進作用,進而加速花青素生物合成[32]。值得注意的是,筆者在本研究中發現,珍富果皮中MdMYB10、MdMYB11表達量分別是2001富士的3.61、6.37倍,這也暗示珍富芽變后也可能存在類似的影響花青苷合成的調控模式,即MdERF3上調表達后與MdMYB10、MdMYB11結合相互作用,進而促進珍富果皮中花青苷合成結構基因的表達及含量的積累,但這種調控模式是否存在及其具體調控花青苷合成作用機制還需進一步解析驗證。
4 結 論
采收期及貯藏14 d后,紅色芽變品種珍富果皮亮度L*值及果皮顏色飽和度C*值均顯著高于2001富士;珍富可溶性固形物、維生素C含量及固酸比顯著高于2001富士,可滴定酸含量顯著低于2001富士,二者果肉硬度及破裂力無顯著差異;珍富果皮中花青苷總量顯著高于2001富士;珍富花青苷合成途徑中的關鍵結構基因及MYB、EFR、bZIP44等轉錄調控基因均顯著上調表達。
參考文獻 References:
[1] OLIVARES-SOTO H,BAST?AS R M,CALDER?N-ORELLANA A,L?PEZ M D. Sunburn control by nets differentially affects the antioxidant properties of fruit peel in ‘Gala and ‘Fuji apples[J]. Horticulture,Environment,and Biotechnology,2020,61(2):241-254.
[2] 張媛雯,趙琳,譚偉. 植物原花青素的應用研究進展[J]. 中國果菜,2023,43(7):24-28.
ZHANG Yuanwen,ZHAO Lin,TAN Wei. Application progress of plant proanthocyanidins[J]. China Fruit & Vegetable,2023,43(7):24-28.
[3] C?MARA J S,LOCATELLI M,PEREIRA J A M,OLIVEIRA H,ARLORIO M,FERNANDES I,PERESTRELO R,FREITAS V,BORDIGA M. Behind the scenes of anthocyanins-from the health benefits to potential applications in food,pharmaceutical and cosmetic fields[J]. Nutrients,2022,14(23):5133.
[4] FANG X,ZHANG L Z,SHANGGUAN L F,WANG L J. MdMYB110a,directly and indirectly,activates the structural genes for the ALA-induced accumulation of anthocyanin in apple[J]. Plant Science,2023,326:111511.
[5] SHI C Y,LIU L,WEI Z F,LIU J W,LI M,YAN Z L,GAO D T. Anthocyanin accumulation and molecular analysis of correlated genes by metabolomics and transcriptomics in sister line apple cultivars[J]. Life,2022,12(8):1246.
[6] 王龍,王芳,湯蕾,童盼盼,張亞若,王江波. 新疆紅肉蘋果不同時期果皮花青苷含量變化及其合成相關基因表達[J]. 江西農業學報,2021,33(8):6-10.
WANG Long,WANG Fang,TANG Lei,TONG Panpan,ZHANG Yaruo,WANG Jiangbo. Changes of anthocyanin content and expression of synthesis-related genes in peel of Xinjiang red flesh apples in different periods[J]. Acta Agriculturae Jiangxi,2021,33(8):6-10.
[7] ZHENG J,LIU L B,TAO H H,AN Y Y,WANG L J. Transcriptomic profiling of apple calli with a focus on the key genes for ALA-induced anthocyanin accumulation[J]. Frontiers in Plant Science,2021,12:640606.
[8] 許海峰. bHLH33與MYB抑制子參與蘋果花青苷生物合成的分子機理[D]. 泰安:山東農業大學,2020.
XU Haifeng. The molecular mechanism underlying anthocyanin biosynthesis in apple using bHLH33 and MYB repressors[D]. Taian:Shandong Agricultural University,2020.
[9] WANG S,ZHANG Z,LI L X,WANG H B,ZHOU H,CHEN X S,FENG S Q. Apple MdMYB306-like inhibits anthocyanin synthesis by directly interacting with MdMYB17 and MdbHLH33[J]. The Plant Journal,2022,110(4):1021-1034.
[10] LI W F,NING G X,ZUO C W,CHU M Y,YANG S J,MA Z H,ZHOU Q,MAO J,CHEN B H. MYB_SH [AL] QKY [RF] transcription factors MdLUX and MdPCL-like promote anthocyanin accumulation through DNA hypomethylation and MdF3H activation in apple[J]. Tree Physiology,2021,41(5):836-848.
[11] 姜生輝. DNA甲基化修飾MdMYB1啟動子調控蘋果花青苷轉運的機理[D]. 泰安:山東農業大學,2020.
JIANG Shenghui. Mechanism of DNA methylation modifies MdMYB1 promoter to regulate anthocyanin transport in apple[D]. Taian:Shandong Agricultural University,2020.
[12] 馬長青. 蘋果MdMYB1啟動子甲基化對套袋果實著色的調控研究[D]. 楊凌:西北農林科技大學,2019.
MA Changqing. Regulation of the methylation of the MdMYB1 promoter on fruit coloration of Malus after bagging treatment[D]. Yangling:Northwest A & F University,2019.
[13] 王美麗. MYB10不同基因型對蘋果果皮顏色的影響[D]. 拉薩:西藏大學,2020.
WANG Meili. Effects of MYB10 different genotypes on the color of apple fruit skin[D]. Lasa:Tibet University,2020.
[14] 安秀紅. 蘋果MdTTG1、MdMYB9與MdMYB11基因調控花青苷合成的作用機制研究[D]. 泰安:山東農業大學,2013.
AN Xiuhong. Studies on the mechanism by which MdTTG1,MdMYB9 and MdMYB11 Regulate anthocyanin biosythesis in apple[D]. Taian:Shandong Agricultural University,2013.
[15] AN J P,YAO J F,XU R R,YOU C X,WANG X F,HAO Y J. Apple bZIP transcription factor MdbZIP44 regulates abscisic acid-promoted anthocyanin accumulation[J]. Plant,Cell & Environment,2018,41(11):2678-2692.
[16] 畢思琦,安建平,王小非,郝玉金,芮麟,李彤,韓月彭,由春香. 蘋果乙烯響應因子MdERF3促進花青苷和原花青苷積累[J]. 園藝學報,2019,46(12):2277-2285.
BI Siqi,AN Jianping,WANG Xiaofei,HAO Yujin,RUI Lin,LI Tong,HAN Yuepeng,YOU Chunxiang. Ethylene response factor MdERF3 promotes anthocyanin and proanthocyanidin accumulation in apple[J]. Acta Horticulturae Sinica,2019,46(12):2277-2285.
[17] 安佰娟,楊小奎,路超. 2001富士蘋果的矮化密植栽培技術要點[J]. 落葉果樹,2020,52(5):79-80.
AN Baijuan,YANG Xiaokui,LU Chao. Key points of dwarfing and close planting techniques of Fuji 2001[J]. Deciduous Fruits,2020,52(5):79-80.
[18] 衛世乾. 不同成熟度蘋果中多酚類物質的含量研究[J]. 許昌學院學報,2019,38(2):99-102.
WEI Shiqian. Study on the contents of polyphenols in apples with different maturity[J]. Journal of Xuchang University,2019,38(2):99-102.
[19] TAKOS A M,UBI B E,ROBINSON S P,WALKER A R. Condensed tannin biosynthesis genes are regulated separately from other flavonoid biosynthesis genes in apple fruit skin[J]. Plant Science,2006,170(3):487-499.
[20] 陳學森,李秀根,毛志泉,王楠,張宗營,馬鋒旺,叢佩華,張玉剛,郭黃萍,王志剛,姜召濤,徐月華. 新種質創造支撐果品產業升級:紅肉蘋果和‘庫爾勒香梨種質資源利用以及‘紅富士芽變選種案例分析[J]. 果樹學報,2021,38(1):128-141.
CHEN Xuesen,LI Xiugen,MAO Zhiquan,WANG Nan,ZHANG Zongying,MA Fengwang,CONG Peihua,ZHANG Yugang,GUO Huangping,WANG Zhigang,JIANG Zhaotao,XU Yuehua. Fruit industry upgrading supported by new germplasm creation:Case study on the utilization of germplasm resources of red-fleshed apple and ‘Kuerlexiangli pear and the sports selection of ‘Red Fuji[J]. Journal of Fruit Science,2021,38(1):128-141.
[21] 陳學森,伊華林,王楠,張敏,姜生輝,徐娟,毛志泉,張宗營,王志剛,姜召濤,徐月華,李建明. 芽變選種推動世界蘋果和柑橘產業優質高效發展案例解讀[J]. 中國農業科學,2022,55(4):755-768.
CHEN Xuesen,YI Hualin,WANG Nan,ZHANG Min,JIANG Shenghui,XU Juan,MAO Zhiquan,ZHANG Zongying,WANG Zhigang,JIANG Zhaotao,XU Yuehua,LI Jianming. Interpretation of the case of bud sports selection to promote the high-quality and efficient development of the worlds apple and Citrus industry[J]. Scientia Agricultura Sinica,2022,55(4):755-768.
[22] 陳學森,毛志泉,王志剛,王楠,張宗營,姜生輝,姜召濤,徐月華,東明學,李建明,隋秀奇. 持續多代芽變選種及其芽變機理揭開‘紅富士在我國蘋果產業獨占鰲頭的謎底[J]. 中國果樹,2020(3):1-5.
CHEN Xuesen,MAO Zhiquan,WANG Zhigang,WANG Nan,ZHANG Zongying,JIANG Shenghui,JIANG Zhaotao,XU Yuehua,DONG Mingxue,LI Jianming,SUI Xiuqi. Continuous multigenerational sports selection and its mechanism reveals the mystery of ‘Red Fuji in Chinas apple industry[J]. China Fruits,2020(3):1-5.
[23] 宋來慶,趙玲玲,劉美英,唐巖,孫燕霞,李元軍,姜中武. 富士蘋果芽變選種特點及影響因素分析[J]. 中國果菜,2016,36(2):54-56.
SONG Laiqing,ZHAO Lingling,LIU Meiying,TANG Yan,SUN Yanxia,LI Yuanjun,JIANG Zhongwu. Analysis of main features and influencing factors of Fuji apple sport selection[J]. China Fruit & Vegetable,2016,36(2):54-56.
[24] AN J P,ZHANG X W,BI S Q,YOU C X,WANG X F,HAO Y J. The ERF transcription factor MdERF38 promotes drought stress-induced anthocyanin biosynthesis in apple[J]. The Plant Journal,2020,101(3):573-589.
[25] LIU M C,PIRRELLO J,CHERVIN C,ROUSTAN J P,BOUZAYEN M. Ethylene control of fruit ripening:Revisiting the complex network of transcriptional regulation[J]. Plant Physiology,2015,169(4):2380-2390.
[26] CORNUAULT V,POS? S,KNOX J P. Disentangling pectic homogalacturonan and rhamnogalacturonan-I polysaccharides:Evidence for sub-populations in fruit parenchyma systems[J]. Food Chemistry,2018,246:275-285.
[27] 鄧秀新,束懷瑞,郝玉金,徐強,韓明玉,張紹鈴,段常青,姜全,易干軍,陳厚彬. 果樹學科百年發展回顧[J]. 農學學報,2018,8(1):24-34.
DENG Xiuxin,SHU Huairui,HAO Yujin,XU Qiang,HAN Mingyu,ZHANG Shaoling,DUAN Changqing,JIANG Quan,YI Ganjun,CHEN Houbin. Review on the centennial development of pomology in China[J]. Journal of Agriculture,2018,8(1):24-34.
[28] 束懷瑞,陳修德. 我國果樹產業發展的時代任務[J]. 中國果樹,2018(2):1-3.
SHU Huairui,CHEN Xiude. The current task of the development of fruits industry in China[J]. China Fruits,2018(2):1-3.
[29] 王金政,毛志泉,叢佩華,呂德國,馬鋒旺,任小林,束懷瑞,李保華,郭玉蓉,郝玉金,姜遠茂,張新忠,楊欣,曹克強,趙政陽,韓振海,霍學喜,魏欽平. 新中國果樹科學研究70年:蘋果[J]. 果樹學報,2019,36(10):1255-1263.
WANG Jinzheng,MAO Zhiquan,CONG Peihua,L? Deguo,MA Fengwang,REN Xiaolin,SHU Huairui,LI Baohua,GUO Yurong,HAO Yujin,JIANG Yuanmao,ZHANG Xinzhong,YANG Xin,CAO Keqiang,ZHAO Zhengyang,HAN Zhenhai,HUO Xuexi,WEI Qinping. Fruit scientific research in New China in the past 70 years:Apple[J]. Journal of Fruit Science,2019,36(10):1255-1263.
[30] MAZZA G,VELIOGLU Y S. Anthocyanins and other phenolic compounds in fruits of red-flesh apples[J]. Food Chemistry,1992,43(2):113-117.
[31] MA H Y,YANG T,LI Y,ZHANG J,WU T,SONG T T,YAO Y C,TIAN J. The long noncoding RNA MdLNC499 bridges MdWRKY1 and MdERF109 function to regulate early-stage light-induced anthocyanin accumulation in apple fruit[J]. The Plant Cell,2021,33(10):3309-3330.
[32] AN J P,WANG X F,LI Y Y,SONG L Q,ZHAO L L,YOU C X,HAO Y J. EIN3-LIKE1,MYB1,and ETHYLENE RESPONSE FACTOR3 act in a regulatory loop that synergistically modulates ethylene biosynthesis and anthocyanin accumulation[J]. Plant Physiology,2018,178(2):808-823.
