飲用水處理工藝中VBNC 狀態(tài)細(xì)菌研究現(xiàn)狀
2024-05-06 06:29:52雍紹文王金山羅曉峰
凈水技術(shù)
2024年4期
關(guān)鍵詞:檢測(cè)
蘭 童,雍紹文,王金山,羅曉峰,錢(qián) 江,劉 成,*,陳 衛(wèi)
(1.河海大學(xué)淺水湖泊綜合治理與資源開(kāi)發(fā)教育部重點(diǎn)實(shí)驗(yàn)室,江蘇南京 210098;2.河海大學(xué)環(huán)境學(xué)院,江蘇南京 210098;3.寧夏水投銀川水務(wù)有限公司,寧夏銀川 750201)
1982 年,Xu 等[1]首次發(fā)現(xiàn)并提出了細(xì)菌的“活的非可培養(yǎng)” 狀態(tài)(viable but non-culturable,VBNC),明確了其基本概念:細(xì)菌在不良外界環(huán)境中,無(wú)法在常規(guī)固體培養(yǎng)基上生長(zhǎng)繁殖形成菌落,但仍然具有代謝活性的一種特殊休眠狀態(tài),在合適條件下可以復(fù)蘇并恢復(fù)可培養(yǎng)性[2]。
VBNC 狀態(tài)的存在影響了基于細(xì)胞可培養(yǎng)性的傳統(tǒng)檢測(cè)方法的準(zhǔn)確性,易導(dǎo)致微生物泄漏問(wèn)題。包秋華等[3]通過(guò)研究1994 年—2021 年Web of Science 數(shù)據(jù)庫(kù)中836 篇關(guān)于VBNC 的文獻(xiàn)發(fā)現(xiàn),細(xì)菌VBNC 研究方向主要集中在微生物學(xué)、傳染病學(xué)、生物化學(xué)分子生物學(xué)等領(lǐng)域,高頻關(guān)鍵詞也反映了目前的研究熱點(diǎn)包括檢測(cè)手段、誘導(dǎo)條件、形成機(jī)制等方面,具體如圖1 所示。 目前對(duì)于飲用水過(guò)程中細(xì)菌的VBNC 狀態(tài)研究相對(duì)較少,部分研究結(jié)果表明一些致病菌能夠在飲用水處理過(guò)程中呈現(xiàn)VBNC狀態(tài),并在輸配水環(huán)節(jié)復(fù)蘇,恢復(fù)致病性,威脅飲用水安全。 然而人們對(duì)該轉(zhuǎn)變過(guò)程的影響因素和作用機(jī)制尚不明確。
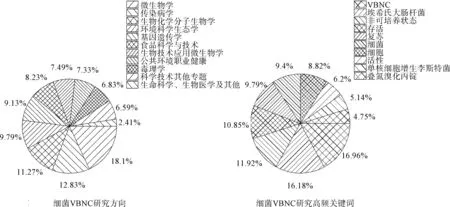
圖1 VBNC 細(xì)菌研究方向和高頻關(guān)鍵詞[3]Fig.1 Research Directions and High Frequency Keywords of VBNC Bacterial[3]
因此,本文從VBNC 狀態(tài)水源性致病菌種類(lèi)、生物學(xué)特性變化、檢測(cè)技術(shù)的發(fā)展以及誘導(dǎo)因素對(duì)目前細(xì)菌VBNC 狀態(tài)的研究進(jìn)行論述,并結(jié)合飲用水處理過(guò)程中的工藝環(huán)節(jié)和水質(zhì)條件,分析細(xì)菌VBNC 狀態(tài)的形成和復(fù)蘇機(jī)制,重點(diǎn)探討了臭氧-生物活性炭(O3-BAC)單元對(duì)細(xì)菌實(shí)現(xiàn)VBNC 狀態(tài)轉(zhuǎn)變的影響。……
登錄APP查看全文
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
