一株馬鈴薯干腐病生防菌的鑒定及其抑菌活性研究
2024-06-08 19:54:05吳慧芳蒙耀石暉琴魏琳沈碩
植物保護 2024年3期
吳慧芳 蒙耀 石暉琴 魏琳 沈碩

摘要
鐮孢菌引起的馬鈴薯干腐病是影響馬鈴薯貯存期間產量與品質的重要病害之一。尋找高效低毒的防治馬鈴薯干腐病的生防藥劑對提高馬鈴薯的產量與質量具有重要意義。本研究以腐皮鐮孢Fusarium?solani為病原菌,測定分離自馬鈴薯塊莖的菌株219CJK2的抑菌活性。該菌株發酵液、無菌發酵液、菌懸液、胞內組分以及揮發性有機物對腐皮鐮孢均具有抑菌活性,抑制率分別為93.37%、61.92%、80.82%、44.01%和49.16%。結合形態學、生理生化特征和多基因系統發育樹構建鑒定其為貝萊斯芽胞桿菌Bacillus?velezensis。馬鈴薯切片試驗和馬鈴薯塊莖試驗表明,當菌株發酵液濃度為1×109?cfu/mL時,對病原菌的抑制效果最好。除此之外,菌株可以分泌蛋白酶、淀粉酶、纖維素酶,具有溶磷能力和生物膜形成能力,但不具有溶血性,并且對供試的其他6種病原真菌有較強的抑菌活性。研究表明貝萊斯芽胞桿菌219CJK2抑菌譜廣,具有生物安全性,對腐皮鐮孢有較強的抑菌活性,在植物病害防治中具有一定的應用潛力,可為馬鈴薯干腐病生防制劑的開發奠定基礎。
關鍵詞
馬鈴薯干腐病;?腐皮鐮孢;?貝萊斯芽胞桿菌;?生物防治
中圖分類號:?S?476
文獻標識碼:?A
DOI:?10.16688/j.zwbh.2023408
Identification?of?a?biocontrol?bacterium?against?potato?dry?rot?and?its?antibacterial?activity
WU?Huifang,?MENG?Yao,?SHI?Huiqin,?WEI?Lin,?SHEN?Shuo*
(Key?Laboratory?of?Biotechnology?in?QinghaiTibet?Plateau,?Ministry?of?Education,?Key?Laboratory?of?Potato
Breeding?in?Qinghai?Province,?Engineering?Research?Center?of?Northwest?Potato,?Ministry?of?Education,
Qinghai?Academy?of?Agricultural?and?Forestry?Sciences,?Qinghai?University,?Xining?810016,?China)
Abstract
Potato?dry?rot?caused?by?Fusarium?is?one?of?the?important?diseases?affecting?both?the?yield?and?quality?of?potato?during?storage.?Finding?highly?efficient?and?lowtoxicity?biocontrol?agents?to?control?potato?dry?rot?holds?great?significance?in?improving?the?yield?and?quality?of?potato.?In?this?study,?the?antifungal?activity?of?strain?219CJK2?isolated?from?potato?tubers?against?Fusarium?solani?was?determined.?The?results?revealed?that?various?components?of?the?strain,?including?the?fermentation?broth,?sterile?fermentation?broth,?bacterial?suspension,?intracellular?components,?and?volatile?organic?compounds,?exhibited?notable?antifungal?activity?against?F.solani,?with?an?inhibition?rate?of?93.37%,?61.92%,?80.82%,?44.01%?and?49.16%,?respectively.?A?combination?of?morphological,?physiological,?and?biochemical?characteristics,?along?with?multigene?phylogenetic?analysis,?it?was?identified?as?Bacillus?velezensis.?Antifungal?test?using?potato?slice?and?tubers?showed?that?the?optimal?control?effect?was?achieved?when?the?concentration?of?the?fermentation?broth?was?1×109?cfu/mL.?In?addition,?the?strain?could?secrete?proteases,?amylases,?and?cellulases,?has?along?with?the?ability?to?dissolve?phosphate?and?form?biofilm,?but?it?lacked?hemolysis?activity?and?displayed?strong?antifungal?activity?against?six?other?pathogenic?fungi.?These?results?showed?that?B.velezensis?219CJK2?had?a?broad?antifungal?spectrum,?biosafety,?and?strong?antifungal?activity?against?F.solani.?Our?study?suggests?its?potential?application?in?the?prevention?and?control?of?plant?diseases,?thereby?laying?a?foundation?for?the?development?of?biocontrol?agents?for?potato?dry?rot.
Key?words
potato?dry?rot;?Fusarium?solani;?Bacillus?velezensis;?biological?control
馬鈴薯Solanum?tuberosum?L.為茄科Solanaceae茄屬Solanum的一年生草本植物,栽培范圍廣、產量高,是僅次于小麥、玉米、水稻之外主要的非谷類主糧作物[1]。在馬鈴薯栽培和貯藏期間會發生多種病害,其中由鐮孢菌Fusarium?spp.引起的馬鈴薯干腐病發生普遍,嚴重影響馬鈴薯的產量和品質,并給加工業和消費者帶來重大損失[2]。該病害在馬鈴薯栽培期和貯藏期均可發生,主要危害塊莖。發病初期薯塊表面出現黑褐色凹陷病斑,隨后擴大形成較多輪紋狀褶皺;后期薯塊內部變空,空腔內布滿菌絲,整個薯塊變硬、變輕、干縮,呈灰褐色或深褐色[3]。
馬鈴薯干腐病病原菌以菌絲體或孢子在病殘組織或土壤中越冬,可以在土壤中存活多年,給該病害的防治帶來很大挑戰[4]。迄今為止,尚未有抗所有致病鐮孢的馬鈴薯品種,且不同鐮孢菌的致病力和危害方式也不同[5]。目前對馬鈴薯干腐病主要采取化學防治的手段,常用藥劑有噻菌靈、抑霉唑和仲丁胺等[67]。但長期使用化學藥劑會導致病菌產生抗藥性和耐藥性,降低化學藥劑的防治效果。因此使用對人體和生態環境無害的生防菌菌劑替代化學藥劑,已成為世界范圍內植物病害防治的方向。
芽胞桿菌作為生防菌的一大類,具有抗逆性強和抑菌譜廣的特點,被廣泛用于植物病害防治。多項研究表明,貝萊斯芽胞桿菌可有效防治核桃黑斑病[8]、番茄灰霉病[9]、煙草青枯病[10]、馬鈴薯炭疽病[11]等多種病害,并且一些芽胞桿菌具有溶磷,產IAA和鐵載體的能力,并能促進植物生長[12]。沈碩[13]研究發現,特氏鹽芽胞桿菌Halobacillus?trueperi對馬鈴薯干腐病病原菌具有良好的抑制活性,抑制率為55.67%,其發酵液可以產生蛋白酶、淀粉酶和纖維素酶等,并且能產生脂肽類抑菌物質。目前,用于馬鈴薯干腐病的芽胞桿菌類生防菌的研究較少,本研究以腐皮鐮孢為病原菌,從多方面評估貝萊斯芽胞桿菌219CJK2的抑菌活性,旨在豐富馬鈴薯生防菌資源,為馬鈴薯干腐病的生物防治提供一定的理論依據。
1?材料與方法
1.1?材料
1.1.1?供試菌株
腐皮鐮孢Fusarium?solani,極細鏈格孢Alternaria?tenuissima,互生鏈格孢Alternaria?alternata,燕麥鐮孢Fusarium?avenaceum,球炭疽菌Colletotrichum?coccodes,立枯絲核菌Rhizoctonia?solani,灰葡萄孢Botrytis?cinerea,由青海省農林科學院微生物研究室保存。菌株219CJK2分離自馬鈴薯塊莖,由青海省農林科學院微生物研究室分離并保存。
1.1.2?供試培養基
PDA培養基:馬鈴薯200?g,葡萄糖20?g,瓊脂20?g,蒸餾水1?000?mL,自然pH?(不添加瓊脂為PDB液體培養基)。
NB培養基:蛋白胨10?g,牛肉膏3?g,氯化鈉5?g,蒸餾水1?000?mL,pH?7.2~7.4?(添加瓊脂為NA固體培養基)。
蒙金娜無機磷培養基:葡萄糖10?g,硫酸銨0.5?g,氯化鈉0.3?g,氯化鉀0.3?g,硫酸鎂0.3?g,硫酸亞鐵0.03?g,硫酸錳0.03?g,磷酸鈣10?g,瓊脂16?g,蒸餾水1?000?mL,pH?7.0~7.5。
蛋白酶檢測培養基:脫脂奶粉12?g,瓊脂粉20?g,蒸餾水1?000?mL,自然pH。
淀粉酶檢測培養基:蛋白胨5?g,牛肉膏3?g,可溶性淀粉10?g,瓊脂18?g,蒸餾水1?000?mL,自然pH。
纖維素酶檢測培養基:?蛋白胨5?g,酵母浸粉5?g,羧甲基纖維素鈉5?g,氯化鈉2.5?g,磷酸二氫鉀0.5?g,瓊脂8?g,蒸餾水500?mL。
血瓊脂培養基:胰酶消化酪蛋白10?g,氯化鈉5?g,肉胃酶消化物5?g,玉米淀粉1?g,心胰酶消化物3?g,酵母浸粉5?g,瓊脂15?g,蒸餾水1?000?mL,pH?7.1~7.5。121℃高壓滅菌15?min,冷卻至55℃左右加入7%~10%無菌脫纖維羊血,混勻后制成平板。無菌脫纖維羊血(貨號TX0020100ml)購自北京索萊寶科技有限公司。
1.2?試驗方法
1.2.1?菌株219CJK2對腐皮鐮孢的抑菌活性
采用平板對峙法[14],將腐皮鐮孢在PDA平板上于28℃黑暗培養5?d,在菌落邊緣打取直徑7?mm的菌餅,置于PDA平板中心,在距離菌餅25?mm相對四點處,用接種針點接生防菌株,以不接種生防菌株為空白對照。待對照菌絲快長滿整個培養皿時測量菌落直徑,并計算生防菌株對腐皮鐮孢的抑制率,每個處理重復3次。抑菌率=(對照菌落直徑-處理菌落直徑)/(對照菌落直徑-菌餅直徑)×100%。在對峙平板上挑取病原菌菌落邊緣菌絲,置于光學顯微鏡下觀察病原菌菌絲形態。
1.2.2?菌株219CJK2培養液不同組分對腐皮鐮孢的抑菌活性
菌株種子液制備:挑取菌株219CJK2單菌落接種在NB液體培養基中,28℃,180?r/min過夜培養,即為種子液。
發酵液制備:將種子液按1%接種量接種到NB液體培養基中,28℃,180?r/min培養72?h,獲得發酵液。
無菌發酵液制備:將發酵液在4℃,12?000?r/min離心15?min,收集上清,經0.22?μm孔徑濾膜過濾除去菌體,即為無菌發酵液。
菌懸液制備:將發酵液在4℃,12?000?r/min離心15?min,收集菌體,使用PBS緩沖液清洗3遍后,加入PBS緩沖液至原體積,即為菌懸液。
胞內組分制備:將菌懸液在冰浴下破碎6?s,間隔6?s,破碎20?min,4℃,12?000?r/min離心15?min,收集上清,即為胞內組分。
在錐形瓶中加入90?mL?PDA培養基并滅菌,待培養基冷卻至50℃左右分別加入10?mL發酵液,無菌發酵液,菌懸液和胞內組分,使各組分的最終含量為10%?(V/V),搖勻后倒平板,以不添加其他成分的PDA培養基為對照,每處理重復3次。平板凝固后在平板中央接入直徑為7?mm的腐皮鐮孢菌餅,28℃培養7?d,測量菌落直徑,計算抑菌率。
1.2.3?菌株219CJK2產生的揮發性有機物對腐皮鐮孢的抑菌活性
采用平板對扣法,將菌株219CJK2的種子液100?μL涂布于NA平板上,將7?mm的腐皮鐮孢菌餅接種于PDA平板中央,將2個平板對扣,用封口膜密封,28℃黑暗培養7?d,測量菌落直徑,計算抑菌率。每處理重復3次,對照使用空白NA平板。
1.2.4?菌株219CJK2的鑒定
1.2.4.1?形態學鑒定
參照《常見細菌系統鑒定手冊》[15]對生防菌株219CJK2進行形態學特征觀察。將菌株在NA平板上劃線,35℃培養48?h,觀察記錄菌落形態、顏色和透明程度,并對菌株219CJK2進行革蘭氏染色并置于顯微鏡下觀察。
1.2.4.2?生理生化鑒定
參照《常見細菌系統鑒定手冊》[15]的方法對生防菌株219CJK2進行甘露醇、乳糖、檸檬酸鹽、丙酸鹽的利用,耐鹽性,VP試驗,硝酸鹽還原等生理生化特性鑒定。
1.2.4.3?分子生物學鑒定
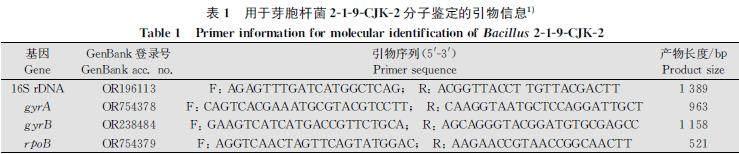
利用細菌基因組DNA提取試劑盒提取菌株219CJK2基因組DNA,選擇可用于鑒定芽胞桿菌種屬的保守基因16S?rDNA、DNA回旋酶亞基A(DNA?gyrase?subunit?A,?gyrA)、DNA回旋酶亞基B(DNA?gyrase?subunit?B,?gyrB)和RNA聚合酶β亞基(RNA?polymerase?subunit?β,?rpoB)進行PCR擴增與測序。PCR擴增產物用1%瓊脂糖凝膠電泳進行檢測后回收,送至艾優稷生物科技(西安)有限公司測序,所得序列上傳至NCBI獲得登錄號(表1)。以軟件MEGA?7.0和Sequence?Matrix將16S?rDNA、gyrA、gyrB、rpoB的基因序列連接后比對,采用最大似然法(自展值設為1?000)構建系統進化樹,確定219CJK2分類地位[16]。
1.2.5?菌株219CJK2發酵液對馬鈴薯干腐病菌的抑制作用
菌株219CJK2不同濃度發酵液制備:按照1.2.2制備生防菌株發酵液,用空白培養基將發酵液分別稀釋為1×105、1×106、1×107、1×108、1×109?cfu/mL。
病原菌孢子懸浮液制備:將腐皮鐮孢Fusarium?solani菌餅接種于PDB培養基中,28℃,180?r/min培養4?d,用3層無菌紗布過濾除去菌絲,濾液在4℃,10?000?r/min離心10?min,棄上清,用無菌水將孢子濃度調整為1×107個/mL。
1.2.5.1?馬鈴薯切片抑菌試驗
參照Loria等[17]的方法,用馬鈴薯塊莖切片法評價菌株219CJK2不同濃度發酵液對腐皮鐮孢的抑菌活性。用自來水洗凈馬鈴薯上的泥土,在室溫下晾干后用1%的次氯酸鈉溶液消毒10?min后切片(直徑27?mm,厚度3?mm)。將馬鈴薯切片放在不同濃度的發酵液中浸泡15?min,以在無菌水中浸泡15?min為對照,置于含有濕潤無菌濾紙的培養皿中。2?h后接種直徑為5?mm的F.solani菌餅。每處理3次重復,在28℃條件下黑暗培養5?d,觀察病原菌在塊莖切片上受抑制情況及馬鈴薯切片發病情況。
1.2.5.2?馬鈴薯塊莖抑菌試驗
參照Zhang等[18]的方法,評價不同濃度發酵液對馬鈴薯干腐病的防治效果。用自來水洗凈馬鈴薯上的泥土,在室溫下晾干后用1%的次氯酸鈉溶液消毒10?min,在超凈臺上晾干。用無菌打孔器在馬鈴薯塊莖表面制造1個圓形傷口(直徑5?mm,深7?mm)。分別接種不同濃度的219CJK2發酵液40?μL,以無菌水作為對照。2?h后,每個創面接種40?μL?F.solani孢子懸浮液(1×107個/mL)。自然干燥后,置于28℃,相對濕度90%~95%的培養箱中貯藏7?d,測量其病灶直徑及腐爛情況。抑菌率=(對照病灶直徑-處理病灶直徑)/(對照病灶直徑-5?mm)×100%。每處理重復3次,每次10個馬鈴薯塊莖。
1.2.6?菌株219CJK2胞外酶活性、解磷能力檢測及溶血安全性的評價
按照1.2.2制備生防菌種子液。采用透明圈法測定生防菌株219CJK2的產酶能力。在蛋白酶檢測培養基、淀粉酶檢測培養基和蒙金娜無機磷培養基平板上各打3孔(d=7?mm),每孔加入100?μL種子液,28℃恒溫培養3?d,查看蛋白酶檢測平板是否出現水解圈,有水解圈表明菌株能產生蛋白酶。將魯戈氏碘液加入淀粉酶檢測平板,若淀粉酶檢測培養基出現水解圈,表明菌株能產生淀粉酶。將適量0.1%剛果紅染液加入纖維素酶檢測平板,靜置染色15?min,用NaCl溶液沖洗,觀察是否出現黃色暈圈,若有則表明菌株能產生纖維素酶。查看蒙金娜無機磷平板是否出現水解圈,有水解圈表明菌株有解磷能力。將菌株劃線接種于血瓊脂平板上,置于35℃恒溫培養箱中倒置培養24?h,觀察菌株溶血現象,判斷其安全性。
1.2.7?菌株219CJK2生物膜形成能力檢測
在6孔板中加入5?mL?LB培養基,而后接種新鮮過夜培養的500?μL種子液,置于35℃培養箱分別靜置培養24?h和48?h;小心取出6孔板,用無菌注射器小心吸出6孔板中生物膜下的培養基和游離細胞,用無菌PBS輕輕柔洗1遍后,加入5?mL甲醇溶液固定15?min;小心棄去甲醇溶液,在室溫下靜置至生物膜干燥,加入5?mL?1%結晶紫溶液染色15?min,小心棄去染色劑,用無菌PBS將染液洗凈,倒置放在37℃恒溫培養箱中烘干;加入5?mL?33%冰醋酸溶液在37℃溫浴30?min使生物膜完全溶解,分光光度計測定OD590。每處理重復6次,以空白培養基為對照。
1.2.8?菌株219CJK2抑菌譜測定
參照1.2.1中平板對峙法測定菌株219CJK2對其他6種病原菌的抑菌活性。
2?結果與分析
2.1?菌株219CJK2對腐皮鐮孢的抑菌活性
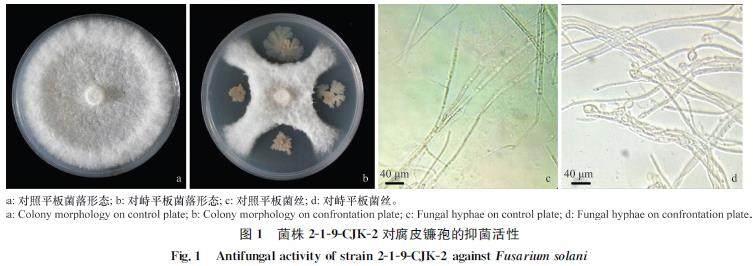
平板對峙試驗結果顯示,當對照菌落長至7.76?cm時,對峙菌落僅長至3.21?cm,生防菌對腐皮鐮孢有很強的抑菌活性,菌絲生長抑制率為64.45%。正常的腐皮鐮孢可形成完整的輻射狀菌落,而生防菌株219CJK2能明顯抑制腐皮鐮孢菌絲的生長。使用顯微鏡觀察對峙菌絲和對照菌絲,發現與正常生長的腐皮鐮孢菌絲相比,與219CJK2對峙培養的腐皮鐮孢菌絲頂端膨大,且形成不規則畸變,整個菌絲呈不規則彎曲凸起狀(圖1)。
2.2?菌株219CJK2不同組分對腐皮鐮孢的抑菌活性
使用添加菌株219CJK2不同組分的培養基進行抑菌活性測定,不同組分對腐皮鐮孢均有抑菌活性(表2)。添加菌株219CJK2發酵液的抑菌活性最高,抑菌率為93.37%;添加菌懸液對腐皮鐮孢的抑菌率為80.82%;添加無菌發酵液對腐皮鐮孢抑菌率為61.92%,添加胞內活性組分對腐皮鐮孢生長抑制能力較弱,抑菌率為44.01%。
2.3?菌株219CJK2揮發性有機物對腐皮鐮孢的抑菌活性
腐皮鐮孢經菌株219CJK2揮發性有機物處理7?d后,對照菌落直徑為7.82?cm,處理菌落直徑為4.32?cm,菌絲生長明顯受到抑制,抑菌率為49.16%,表明菌株219CJK2揮發性有機物對腐皮鐮孢有較好的抑菌活性。
2.4?菌株219CJK2的鑒定結果
2.4.1?形態學鑒定
菌株219CJK2在NA平板上生長狀態良好,35℃培養48?h后,菌落圓形或不規則形,邊緣光滑濕潤,表皮有褶皺,乳白色不透明,挑取時呈黏稠狀。顯微鏡觀察菌株呈單細胞桿狀,革蘭氏染色呈陽性(圖2)。
2.4.2?生理生化鑒定
菌株219CJK2能利用甘露醇、乳糖和葡萄糖,VP試驗陽性,能在7%氯化鈉、pH?5.7條件下生長,不能利用檸檬酸鹽和丙酸鹽,不能還原硝酸鹽,不產生IAA(表3)。
2.4.3?分子生物學鑒定
基于菌株219CJK2的16S?rDNA、gyrA、gyrB和rpoB基因序列,構建219CJK2多基因系統進化樹(圖3)。結果表明,菌株219CJK2與貝萊斯芽胞桿菌LMG?22478聚在同一支。綜合菌落形態、生理生化和分子生物學鑒定結果,最終將菌株219CJK2鑒定為貝萊斯芽胞桿菌Bacillus?velezensis。
2.5?菌株219CJK2發酵液對馬鈴薯干腐病的抑制效果
2.5.1?馬鈴薯切片抑菌試驗結果
不同濃度的219CJK2發酵液對腐皮鐮孢引起的馬鈴薯切片腐爛有不同的抑制效果(圖4A)。對照切片上布滿菌絲,且呈現出褐變。與對照相比,1×109?cfu/mL的219CJK2處理防治效果最好,切片沒有發生褐變,菌絲在切片上幾乎不生長,抑制效果呈現濃度依賴性。
2.5.2?馬鈴薯塊莖抑菌試驗結果
菌株219CJK2對馬鈴薯塊莖上腐皮鐮孢的抑制效果如圖4B和表4。由圖表可看出,馬鈴薯塊莖的病灶直徑隨著發酵液濃度增加而減小,且明顯小于對照。防治效果隨著發酵液濃度的增加,呈上升趨勢。1×105、1×106、1×107、1×108?cfu/mL和1×109?cfu/mL濃度處理的防治效果分別為26.92%、40.44%、57.85%、80.99%和88.55%。結果表明,219CJK2對馬鈴薯干腐病具有明顯的抑制效果。
2.6?菌株219CJK2胞外酶活性、解磷能力檢測及溶血安全性的評價
胞外酶活性及解磷能力檢測結果表明,生防菌219CJK2具有產蛋白酶、淀粉酶、纖維素酶和解磷的能力(圖5)。
將菌株219CJK2接種于血瓊脂平板中培養24?h后,溶血性試驗結果如圖5f所示。結果顯示,在培養24?h后,血瓊脂平板上未出現溶血現象,因此可認為菌株219CJK2不具溶血性,初步認為219CJK2為一株安全的菌株。
2.7?菌株219CJK2生物膜形成能力
結晶紫染色后,孔板內壁形成一圈紫環,說明菌株219CJK2能形成生物膜。靜置培養24?h和48?h,其在590?nm處的吸光度分別為2.989和3.240,說明48?h生物膜形成量比24?h略有增長。
2.8?菌株219CJK2的抑菌譜
平板對峙試驗結果表明,菌株219CJK2對其他6種病原菌均表現出良好的抑菌活性(圖6),抑菌率均在55%以上,其中對球炭疽菌的抑菌活性最強,抑菌率可達86.60%,對互生鏈格孢的抑菌活性最弱,抑菌率為59.77%(表5)。
3?結論與討論
農業生產上經常依賴化學藥劑和化肥來獲得豐厚的農業產出[19]。在過去的幾個世紀里,農藥對提高作物的質量和產量起到了非常重要的作用[20]。近年來,由于環境污染和農藥殘留對人類健康和地球生態系統的影響,人們對化學肥料和農藥的態度發生了變化[21]。相對于化學防治,生物防治具有專一性,對環境友好,對人類和動物安全無害,是化學防治的良好替代措施[22]。芽胞桿菌已成為很有吸引力的生物菌劑,其能夠產生堅硬、耐藥的芽胞和抗生素,對多種病原菌都有抑制作用[23]。據報道,芽胞桿菌屬中的貝萊斯芽胞桿菌可以抑制多種微生物病原體的生長,包括細菌、真菌和線蟲[24]。
本研究從馬鈴薯塊莖中分離得到一株生防細菌219CJK2,其對馬鈴薯干腐病病原腐皮鐮孢有很強的抑菌活性,抑制率為64.45%。對其不同組分抑菌活性進行研究發現,發酵液的抑菌活性最高,抑制率為93.37%,胞內組分抑菌活性最低,抑制率為44.01%。微生物產生的次生代謝產物有的分泌于細胞外,形成細胞外次生代謝產物,有的則留在細胞內,形成細胞內次生代謝產物[25]。目前對于芽胞桿菌的抑菌活性物質的研究多集中在細胞外代謝產物,對于芽胞桿菌細胞內代謝產物研究甚少[26],菌株219CJK2的胞內抑菌活性物質有待進一步研究。無菌發酵液的抑制率為61.92%,有較強的抑菌活性,說明菌株219CJK2產生的胞外次級代謝產物對病原菌有良好的抑菌活性。
細菌產生的揮發性有機物(volatile?organic?compounds,?VOCs),能夠影響病原真菌的生長代謝,促進植物的生長。Fernando等[27]從油菜和大豆以及相關作物土壤中分離出的假單胞菌Pseudomonas?adaceae產生的揮發物有抑制菌核活性和限制子囊孢子產生和萌發的作用。陳奕鵬等[28]從健康香蕉植株分離得到一株解淀粉芽胞桿菌Bacillus?amyloliquefaciens?BEB17,該菌株揮發性有機物可抑制多種病原菌生長,并能抑制病原菌孢子萌發。菌株219CJK2揮發性有機物對腐皮鐮孢具有較強的抑菌活性,有開發為生物熏蒸劑的潛力,其產生的揮發性有機物物質還待進一步研究鑒定。
胞外酶可以通過溶解病原菌的細胞壁、抑制菌絲生長、孢子生長等,干擾病原菌對植物的侵染,從而防治植物病害,可以用來評估生防菌的生防潛力[2930]。已報道的相關胞外酶有幾丁質酶、蛋白酶、纖維素酶等。張紅霞等[31]發現致病疫霉Phytophthora?infestans可誘導細菌HT6產生纖維素酶,且纖維素酶在抑制致病疫霉菌絲生長和引起菌絲體畸變中發揮了重要作用。土壤中含有較多磷元素,但大多以溶解度極低的化合物形態存在,難以被植物吸收和利用,自然環境中普遍存在可利用磷缺少的情況[3234]。芽胞桿菌是主要的解磷細菌之一,能通過分泌有機酸溶解土壤中的無機磷[35],促進植物生長和對其他微量營養元素的吸收。本研究獲得的生防菌219CJK2能夠分泌蛋白酶、淀粉酶和纖維素酶,同時具有溶磷能力,表明分泌水解酶是其抑菌機理之一,且菌株219CJK2具有潛在的促生能力。
生防菌在植物體內成功定殖直接決定其生防能力,而菌株生物膜的形成是菌株定殖成功的關鍵因素。司方潔等[9]通過構建貝萊斯芽胞桿菌5YN8生物膜相關基因突變體發現,生物膜形成能力增強突變體菌株在番茄葉面的定殖能力增強,對番茄灰霉病的防治效果顯著增強,為80.77%,而生物膜形成能力減弱突變體菌株在葉面的定殖能力顯著降低,對灰霉病的防效顯著降低,僅為16.02%。菌株219CJK2具有形成生物膜的能力,且隨著時間增長,生物膜形成量增加。有溶血性的芽胞桿菌具有致病性,能引起人和動物的多種疾病[36]。菌株219CJK2溶血安全性試驗表明,菌株不具有溶血性,可作為開發為生防菌劑的候選菌株。
本研究以腐皮鐮孢為病原菌,檢測貝萊斯芽胞桿菌219CJK2的抑菌活性。該菌株發酵液、無菌發酵液、菌懸液、胞內組分和揮發性有機物對腐皮鐮孢均具有良好的抑菌活性。該菌株能分泌多種胞外酶,具有解磷能力、生物膜形成能力和溶血安全性,對其他6種植物病原菌均具有抑菌活性,是一株有應用潛力的生防菌株。接下來將進一步探索菌株219CJK2的抑菌機理,通過盆栽試驗檢測生防菌株實際抑菌效果和抑菌穩定性,為生防菌的開發和利用奠定基礎。
參考文獻
[1]?CAMIRE?M?E,?KUBOW?S,?DONNELLY?D?J.?Potatoes?and?human?health?[J].?Critical?Reviews?in?Food?Science?and?Nutrition,?2009,?49(10):?823840.
[2]?LUI?L?H,?KUSHALAPPA?A?C.?Response?surface?models?to?predict?potato?tuber?infection?by?Fusarium?sambucinum?from?duration?of?wetness?and?temperature,?and?dry?rot?lesion?expansion?from?storage?time?and?temperature?[J].?Internal?Journal?of?Food?Microbiology,?2002,?76(1/2):?1925.
[3]?李金花,?王蒂,?柴兆祥,?等.?甘肅省馬鈴薯鐮刀菌干腐病優勢病原的分離鑒定[J].?植物病理學報,?2011,?41(5):?456463.
[4]?BOJANOWSKI?A,?AVIS?T?J,?PELLETIER?S,?et?al.?Management?of?potato?dry?rot?[J].?Postharvest?Biology?Technology,?2013,?84:?99109.
[5]?王文重,?高云飛,?閔凡祥,?等.?馬鈴薯鐮刀菌干腐病研究進展及防控手段[J].?植物保護,?2020,?46(5):?612.
[6]?CARNEGIE?S?F,?RUTHVEN?A?D,?LINDSAY?D?A,?et?al.?Effects?of?fungicides?applied?to?seed?potato?tubers?at?harvest?or?after?grading?on?fungal?storage?diseases?and?plant?development?[J].?Annals?of?Applied?Biology,?2010,?116(1):?6172.
[7]?WHARTON?P?S,?KIRK?W?W,?BERRY?D,?et?al.?Seed?treatment?applicationtiming?options?for?control?of?Fusarium?decay?and?sprout?rot?of?cut?seedpieces?[J].?American?Journal?of?Potato?Research,?2007,?84(3):?237244.
[8]?王迪,?高巖,?吳小雙,?等.?兩株貝萊斯芽胞桿菌鑒定及其對薄殼山核桃黑斑病病原菌的拮抗效果[J].?中國生物防治學報,?2022,?38(6):?15721581.
[9]?司方潔,?任金瑤,?黃濤祥,?等.?貝萊斯芽胞桿菌5YN8生物被膜在防治番茄灰霉病過程中的功能研究[J].?中國生物防治學報,?2022,?38(5):?12231230.
[10]周向平,?滕凱,?肖啟明,?等.?貝萊斯芽胞桿菌F10促生作用及對煙草青枯病的防治效果[J].?煙草科技,?2022,?55(7):?916.
[11]李統華.?馬鈴薯炭疽菌生防細菌的篩選、鑒定及發酵工藝優化[D].?蘭州:?甘肅農業大學,?2022.
[12]HASHEM?A,?TABASSUM?B,?FATHI?ABD?ALLAH?E.?Bacillus?subtilis:?A?plantgrowth?promoting?rhizobacterium?that?also?impacts?biotic?stress?[J].?Saudi?Journal?of?Biological?Sciences,?2019,?26(6):?12911297.
[13]沈碩.?中度嗜鹽菌S61生防因子分析及其對馬鈴薯干腐病的防效[J].?南方農業學報,?2021,?52(10):?26192631.
[14]ZHAO?Yueju,?SELVARAJ?J?N,?XING?Fuguo,?et?al.?Antagonistic?action?of?Bacillus?subtilis?strain?SG6?on?Fusarium?graminearum?[J/OL].?PLoS?ONE,?2014,?9(3):?e92486.?DOI:?10.1371/journal.pone.0092486.
[15]東秀珠,?蔡妙英.?常見細菌系統鑒定手冊[M].?北京:?科學出版社,?2001.
[16]LI?Jieling,?HU?Ming,?XUE?Yang,?et?al.?Screening,?identification?and?efficacy?evaluation?of?antagonistic?bacteria?for?biocontrol?of?soft?rot?disease?caused?by?Dickeya?zeae?[J/OL].?Microorganisms,?2020,?8(5):?697.?DOI:?10.3390/microorganisms8050697.
[17]LORIA?R,?BUKHALID?R?A,?CREATH?R?A,?et?al.?Differential?production?of?thaxtomins?by?pathogenic?Streptomyces?species?in?vitro?[J].?Phytopathology,?1995,?85:?537541.
[18]ZHANG?Xiaoyun,?WU?Feng,?GU?Ning,?et?al.?Postharvest?biological?control?of?rhizopus?rot?and?the?mechanisms?involved?in?induced?disease?resistance?of?peaches?by?Pichia?membranefaciens?[J/OL].?Postharvest?Biology?and?Technology,?2020,?163(1):?111146.?DOI:10.1016/j.postharvbio.2020.111146.
[19]GAGIC?V,?KLEIJN?D,?BLDI?A,?et?al.?Combined?effects?of?agrochemicals?and?ecosystem?services?on?crop?yield?across?Europe?[J].?Ecology?Letter,?2017,?20(11):?14271436.
[20]SANCHEZ?P?A,?BUOL?S?W.?Soils?of?the?tropics?and?the?world?food?crisis?[J].?Science,?1975,?188(4188):?598603.
[21]CULLEN?M?G,?THOMPSON?L?J,?CAROLAN?J?C,?et?al.?Fungicides,?herbicides?and?bees:?A?systematic?review?of?existing?research?and?methods?[J/OL].?PLoS?ONE,?2019,?14(12):?e0225743.?DOI:?10.1371/journal.pone.0225743.
[22]TEEM?J?L,?ALPHEY?L,?DESCAMPS?S,?et?al.?Genetic?biocontrol?for?invasive?species?[J/OL].?Frontiers?Bioengineering?Biotechnology,?2020,?8:?452.?DOI:?10.3389/fbioe.2020.00452.
[23]GUAN?Junfeng,?ZENG?Kaifang,?CHEN?Zhihui.?Editorial:?Postharvest?disease?management?in?fruits?and?vegetables:?recent?advances?and?mechanisms?[J/OL].?Frontiers?in?Microbiology,?2023,?14:?1203010.?DOI:?10.3389/fmicb.2023.1203010.
[24]RABBEE?M?F,?ALI?M?S,?CHOI?J,?et?al.?Bacillus?velezensis:?A?valuable?member?of?bioactive?molecules?within?plant?microbiomes?[J/OL].?Molecules,?2019,?24(6):?1046.?DOI:?10.3390/molecules24061046.
[25]劉霞,?李騫,?許賢,?等.?昆蟲病原線蟲共生菌YL001細胞內代謝產物抑菌作用研究初報[J].?農藥學學報,?2006,?8(1):?9598.
[26]XU?Zhihui,?MANDICMULEC?I,?ZHANG?Huihui,?et?al.?Antibiotic?bacillomycin?D?affects?iron?acquisition?and?biofilm?formation?in?Bacillus?velezensis?through?a?Btrmediated?FeuABCdependent?pathway?[J].?Cell?Reports,?2019,?29(5):?11921202.
[27]FERNANDO?W?G?D,?RAMARATHNAM?R,?KRISHNAMOORTHY?A?S,?et?al.?Identification?and?use?of?potential?bacterial?organic?antifungal?volatiles?in?biocontrol?[J].?Soil?Biology?Biochemistry,?2005,?37(5):?955964.
[28]陳奕鵬,?楊揚,?桑建偉,?等.?拮抗內生芽孢桿菌BEB17分離鑒定及其揮發性物質抑菌活性分析[J].?植物病理學報,?2018,?48(4):?537546.
[29]李桂娥,?吳小芹.?水拉恩氏菌JZGX1及其誘變菌株對馬尾松苗的促生效應[J].?南京林業大學學報(自然科學版),?2014,?38(3):?8387.
[30]陳思宇,?陳志誼,?張榮勝.?水稻紋枯病菌拮抗細菌的篩選及鑒定[J].?植物保護學報,?2013,?40(3):?211218.
[31]張紅霞,?蔣繼志,?張荷花,?等.?致病疫霉誘導細菌HT6產生纖維素酶發揮抑菌作用[J].?河北農業大學學報,?2019,?42(4):?8287.
[32]GOLDSTEIN?A?H.?Bacterial?solubilization?of?mineral?phosphates:?Historical?perspective?and?future?prospects?[J].?American?Journal?of?Alternative?Agriculture,?1986,?1(2):?5157.
[33]SUNDARA?B,?NATARAJAN?V,?HARI?K.?Influence?of?phosphorus?solubilizing?bacteria?on?the?changes?in?soil?available?phosphorus?and?sugarcane?and?sugar?yields?[J].?Field?Crops?Research,?2002,?77(1):?4349.
[34]李雪艷,?張濤,?楊紅梅,?等.?棉花黃萎病拮抗細菌產鐵載體測定及其對抑菌活性的影響[J].?微生物學通報,?2019,?46(5):?10741080.
[35]SANG?Yue,?JIN?Long,?ZHU?Rui,?et?al.?Phosphorussolubilizing?capacity?of?Mortierella?species?isolated?from?rhizosphere?soil?of?a?poplar?plantation?[J/OL].?Microorganisms,?2022,?10(12):?2361.?DOI:?10.3390/microorganisms10122361.
[36]王捷.?蘋果腐爛病生防芽孢桿菌的篩選鑒定及其防治效果初探[D].?楊凌:?西北農林科技大學,?2022.
(責任編輯:楊明麗)
