石榴蔗糖代謝相關酶SPS和INV基因家族鑒定與表達分析
2024-06-11 20:39:45馮立娟李英朋王傳增尹燕雷郭琳譚偉
山東農業科學 2024年3期
馮立娟 李英朋 王傳增 尹燕雷 郭琳 譚偉

摘要:蔗糖磷酸合成酶(SPS)和蔗糖轉化酶(INV)是蔗糖代謝的關鍵調控酶,在植物生長發育過程中起重要作用。本研究利用生物信息學和熒光定量PCR等分子手段,鑒定石榴SPS和INV基因家族成員,分析其理化性質、保守結構域、保守基序、二級結構、亞細胞定位、系統進化關系和表達模式。結果表明,從石榴基因組中鑒定出4個SPS基因和11個INV基因,其編碼蛋白均為不穩定蛋白,具有典型的保守結構域,家族成員間特征motif數量和種類大致相同,蛋白結構高度保守:這些蛋白不均勻地分布在染色體上,均定位于葉綠體中,二級結構主要由α-螺旋和無規則卷曲組成。石榴SPS和INV基因家族成員間存在不同程度的親緣關系,與巨桉同源性較高;不同SPS和INV基因在石榴果實不同發育時期的表達模式存在差異,PgINV3在9月15日(果實增大期)表達水平最高,顯著高于其他時期。本研究結果對解析石榴果實中蔗糖代謝的分子機理具有重要意義。
關鍵詞:石榴:SPS基因:INV基因:生物信息學分析:基因表達
中圖分類號:S665.4:Q781 文獻標識號:A 文章編號:1001-4942(2024)03-0011-08
石榴(Punica granatum L)是我國重要的特色果樹之一,果實營養價值高,保健功能強,越來越受到消費者青睞。山東石榴栽培歷史悠久,種質資源豐富,發展面積日益增加,成為山東省打造鄉村振興齊魯樣板的良好選擇。果實品質提升是提高石榴市場競爭力的重要途徑。蔗糖積累是決定石榴果實風味和品質的重要因子,在其生長發育和產量形成過程中起重要作用。研究石榴果實蔗糖代謝機理對其果實品質調控具有重要的理論意義。
蔗糖磷酸合成酶(sucrose phosphate synthase,SPS)催化尿苷二磷酸葡糖(UDPG)和果糖-6-磷酸(F6P)生成蔗糖-6-磷酸(S6P),S6P在蔗糖磷酸酯酶(SPP)作用下不可逆形成蔗糖。SPS是植物體內控制蔗糖合成的關鍵酶,由多基因家族編碼,在不同物種中的成員數量不同,且同一物種中不同家族成員調控蔗糖合成的能力也不相同。目前已在柑橘和甜櫻桃中鑒定出4個SPS基因,在蘋果和梨中鑒定出8個SPS基因。蔗糖轉化酶(invertase,INV)不可逆地催化蔗糖裂解為果糖和葡萄糖,分為酸性細胞壁轉化酶(CWIN),酸性液泡轉化酶(VIN)和堿性/中性轉化酶(A/N-Inv)。在草莓中鑒定出8個Fa,A/N-Inv基因家族成員,其中4個不僅具有組織特異性,還具有果實發育階段特異性和品種特異性表達特點,受細胞分裂素、赤霉素和生長素誘導。過表達SoSPS1基因可提高轉基因甘蔗葉片的SPS活性和蔗糖含量,提高可溶性酸性轉化酶(SAI)活性及葡萄糖和果糖水平。
目前,國內外在石榴蔗糖代謝方面的研究甚少,SPS和INV基因家族成員鑒定與表達方面的研究尚未見報道。因此,本研究基于全基因組測序結果對石榴SPS和INV基因家族成員進行鑒定,分析其理化性質、保守結構、二級結構及系統進化關系等生物信息學特性,并解析其在果實發育過程中的表達模式,以期為深入研究石榴果實蔗糖積累的分子機制提供理論依據。
1 材料與方法
1.1 石榴SPS和INV基因家族成員鑒定
從NCBI數據庫中下載PgSPS和PgINV蛋白序列,與石榴全基因組(ASM765513v2)數據庫進行同源比對,初步獲得SPS和INV候選序列。通過Pfam在線數據庫(http://pfam.xfam.org)進行進一步的蔗糖合成結構域(PF00862)、糖基轉移結構域(PF00534)和蔗糖-6-磷酸磷酸水解酶結構域(PF05116)驗證。利用SMART(http://smart.embl-heidelberg.de)進行蛋白結構域分析,去除結構不完整的序列,最終確定目的基因。
1.2 石榴SPS和INV基因家族成員生物信息學分析
利用在線軟件ProtParam預測石榴SPS和INV蛋白分子量、等電點、脂肪系數等理化性質。利用CDD和MEME軟件分析保守結構域和保守基序。利用SOPMA和Plant-mPLoc軟件預測二級結構和亞細胞定位。
1.3 系統進化樹構建
使用MEGA6.0軟件,采用NJ法(neighbor-joining)對石榴、蘋果、葡萄和巨桉的SPS和INV蛋白進行系統進化樹構建。Bootstrap值設為1000,去除Bootstrap支持率低于50%的節點,顯示各分支長度。
1.4 熒光定量表達分析
2022年7月15日開始采集‘泰山紅石榴果實,分別在7月30日、8月15日、8月30日、9月15日、9月30日采1次,直至果實成熟(10月15日)。利用RNA prep Pure Plant Kit試劑盒提取籽粒總RNA,反轉錄成cDNA。使用SYBR GreenPCR Master Mix試劑盒、利用BIO-RAD IQ5實時熒光定量PCR儀進行qRT-PCR分析。每個樣品設3次生物學重復。采用Primer Premier 5.0軟件設計引物(表1)。以石榴Actin(GU376750.1)為內參,默認條件下讀取Ct值,相對表達量用2-△△Ct法計算。
1.5 數據處理與分析
利用Microsoft Excel 2010進行數據處理及作圖,用SPSS 19.0軟件進行差異顯著性分析。
2 結果與分析
2.1 石榴SPS和INV基因家族成員鑒定
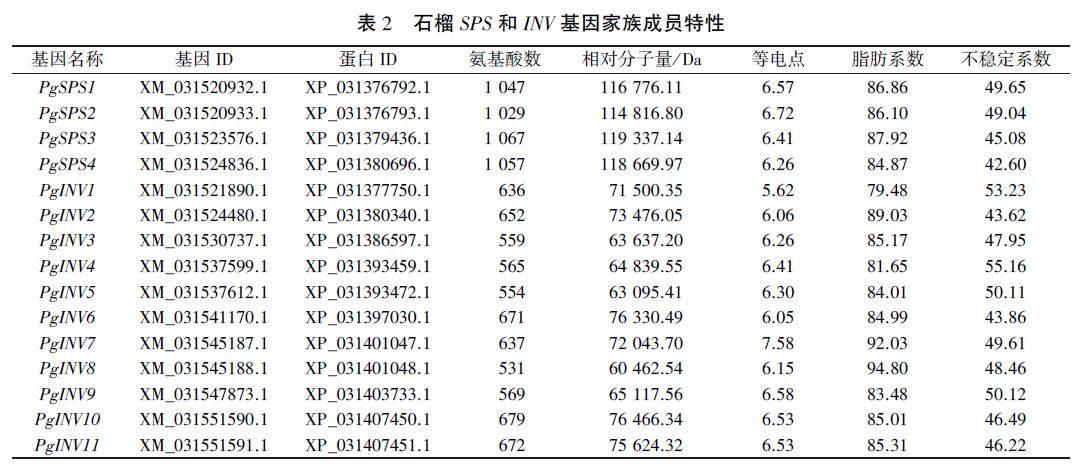
從石榴中鑒定出4個SPS基因和11個INV基因,分別命筆為PgSPS1-PgSPS4和PgINV1-PgINV11(表2)。通過分析蛋白理化性質可知,4個SPS蛋白的氨基酸數量在1029-1067之間,相對分子量在114816.80-119337.14 Da范圍內:等電點在6.26-6.72之間,均在酸性范圍內:脂肪系數在84.87-87.92之間,不穩定系數在42.60-49.65范圍內,均為不穩定蛋白。11個INV家族成員的氨基酸數量在531-679范圍內,等電點在5.62-7.58之間:脂肪系數為79.48-94.80.不穩定系數介于43.62-55.16之間,也均為不穩定蛋白。
2.2 石榴SPS和INV蛋白保守結構域分析
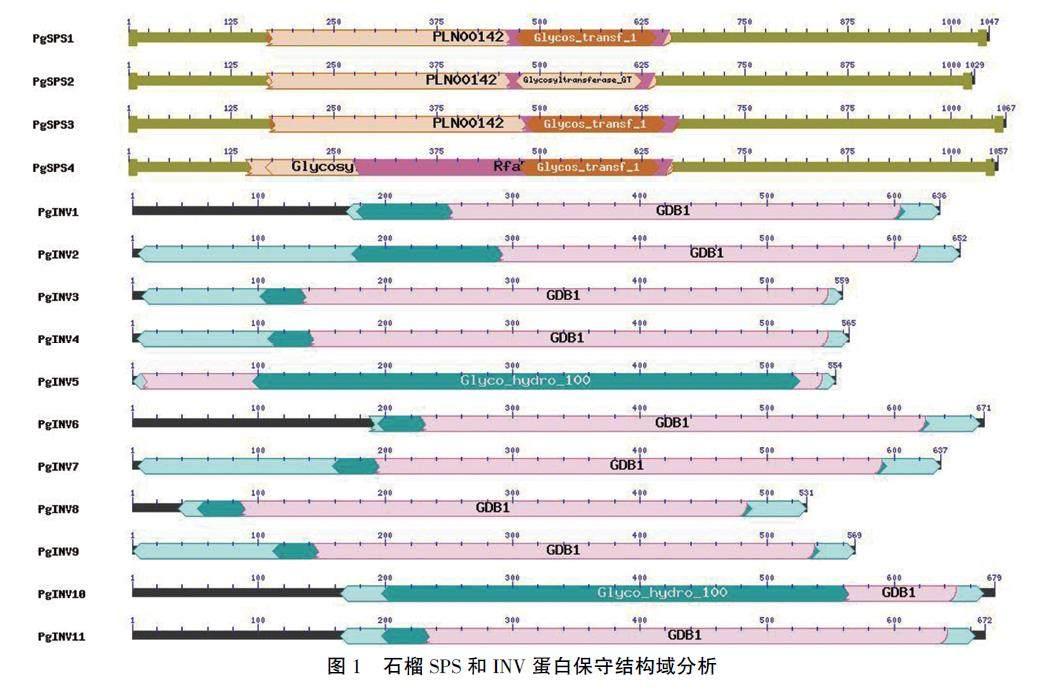
PgSPS1、PgSP52和PgSPS3蛋白屬于PLN00142超級家族成員,其中PgSPSI和PgSP53包含糖基轉移酶(Clycos_transf_1)保守結構域,PgSPS2含GlycosVltransferase_GT保守結構域;Pg-SPS4也含有Glycos_transf_1保守結構域(圖1)。
PgINV5和PgINV10含有Glyco_hydro_100糖基水解酶結構域。除PgINV5外,其余10個INV蛋白均含有GDB1結構域。
2.3 石榴SPS和INV蛋白保守基序分析
由圖2可見,4個SPS家族成員的保守基序數量均為7,且均為Motif2、Motif9、Motif11、Mo-tif12、Motif13、Motif14和Motif15基序。11個INV家族成員均包含12個保守基序,分別為Motif1、Motif2、Motif3、Motif4、Motif5、Motif6、Motif7、Mo-tif8、Motif9、Motif10、Motif11和Motif12。同一基因家族成員間的保守基序數量和種類大致相同。
2.4 石榴SPS和INV各成員蛋白的二級結構與亞細胞定位
又表3可見,石榴SPS和INV蛋白二級結構主要由α-螺旋、β-轉角、延伸鏈和無規則卷曲組成,以α-螺旋和無規則卷曲占比較高;其中,Pg-SPS3及PgINV1、PgINV2、PgINV6、PgINV7、Pg-INVII各組分占比表現為無規則卷曲>α-螺旋>延伸鏈>β-轉角,其他蛋白則均表現為α-螺旋>無規則卷曲>延伸鏈>β-轉角。亞細胞定位預測表明,石榴SPS和INV家族成員均定位于葉綠體中。
染色體分布顯示,pgSPS1和PgSPS2位于Chr1,其余兩個SPS基因位于Chr2。PgINV1、PgINV6、Pg-INV10、PgINV11位于Chr1,PgINV4和PgINV5位于Chr4,PgINV7和PgINV8位于Chr6,PgINV2、PgINV3和PgINV9分別位于Chr2、Chr3和Chr7。
2.5 石榴SPS和INV基因系統進化樹分析
用4個石榴SPS蛋白、5個巨桉SPS蛋白、9個蘋果SPS蛋白和10個葡萄SPS蛋白構建系統進化樹(圖3A),可分為兩大類。PgSPS1、PgSP52和PgSPS4聚為一類,其中PgSPS1和Pg-SPS2聚為一小類。PgSPS3則與EgSPS2、MdSPS2和MsSPS2聚為一類,親緣關系較近。
11個石榴INV蛋白、9個巨桉INV蛋白、7個蘋果INV蛋白和10個葡萄INV蛋白的系統進化樹如圖3B所示,也主要分為兩大類。在第一大類中,PgINV2、PgINV7和PgINV8聚為一類,其中PgINV2與EgINV2聚為一小類,PgINV7和Pg-INV8聚為一小類:PgINV6、PgINV10和PgINV11聚為一類,其中PgINV6與EgINV5親緣關系較近,PgINV10和PgINV11緣關系較近。在第二大類中,PgINV1與EgINV4聚為一類,PgINV3與PgINV5親緣關系較近。PgINV4與PgINV9聚為一類,其中PgINV4與EgINV3、EgINV6聚為一小類,PgINV9與EgINV8親緣關系較近。
2.6 PgSPSs和PgINVs基因表達分析
本研究選用2個SPS基因(PgSPS2和PgASPS4)、6個INV基因(PgINV2、PgINV3、Pg-INV4、PgINV7、PgINV9、PgINV11),均以7月15日的表達量為1,分析其在石榴果實發育期的表達模式,結果(圖4)顯示,兩個SPS基因均下調表達,但隨著石榴果實發育進程的推進,其變化趨勢存在差異,PgSP52表現為降→升→降→升→降→升的較規律波動變化,PgSPS4則表現為降→升→降→升,均以9月15日的表達水平較高。PgINV2整體下調表達,隨著生育進程的推進呈先降低后升高的變化趨勢;PgINV3在7月30日、9月15日和10月15日上調表達,以9月15日的相對表達量最高,8月30日的相對表達量最低;PgINV4、PgINV7和PgINV9在石榴果實發育期間的表達水平較低,除PgINV7在8月15日下調表達較少外,其余時期均明顯下調表達:PgINV11的變化趨勢與PgINV3相似,但僅在9月15日明顯上調表達。
3 討論與結論
蔗糖是植物體內光合產物從“源器官”運輸到“庫器官”的主要形式,是影響產量和果實品質形成的重要因素。SPS和INV是蔗糖代謝的關鍵調控酶,影響植物生長發育過程中的生物量形成和糖分積累。本研究從石榴中鑒定出4個SPS基因,其編碼蛋白均為不穩定的酸性蛋白,與柑橘和甜櫻桃中的數量一致:鑒定出11個INV基因家族成員,均為不穩定蛋白,數量少于無籽蜜柚而多于草莓。
通過對蛋白保守結構域和保守基序分析發現,石榴SPS和INV蛋白均具有典型的保守結構域,家族成員間特征motif數量和種類大致相同,蛋白結構高度保守,這表明石榴SPS和INV基因家族在進化上具有保守性,這與蘋果和獼猴桃上的研究結果一致。其中,PgINV5和Pg-INV10具有中堿性蔗糖轉化酶典型的Glyco_hydro_100糖基水解酶結構域,等電點分別為6.30和6.53,有可能是中堿性蔗糖轉化酶,這與黑皮果蔗和南瓜中的研究結果相似。
蛋白質二級結構的預測與空間結構分析對了解其功能具有重要意義。石榴SPS和INV蛋白二級結構主要由α-螺旋和無規則卷曲組成,延伸鏈和β-轉角所占比例較低,說明α-螺旋和無規則卷曲在石榴SPS和INV蛋白結構中起重要作用,這與甘蔗中的研究結果相似。本研究發現,石榴SPS和INV家族成員不均勻地分布在染色體上,均定位于葉綠體中,其精確定位和調控功能還需深入研究。系統進化樹分析表明,石榴SPS和INV基因家族成員間存在不同程度的親緣關系,石榴與巨桉、蘋果的SPS基因家族成員同源性較高,與巨桉的INV基因家族成員同源性較高。這進一步證實了石榴與巨桉的親緣關系較近,為深入研究石榴SPS和INV基因家族成員的生物學功能奠定了理論基礎。
本研究進一步對2個SPS基因和6個INV基因在石榴果實不同發育時期的表達模式進行了分析,發現其在不同時期的表達量存在差異。在9月15日(果實增大期),PgSPS2和PgSPS4的相對表達量較高,PgINV3和PgINV11的相對表達量顯著高于其他時期。說明石榴SP^S和INV基因家族不同成員調控蔗糖代謝的功能具有特異性,這與甘蔗和萱草中的研究結果一致。
綜上,不同SPS和INV基因調控石榴果實蔗糖代謝的機理存在差異,具有發育時期表達特異性,后續將從轉錄調控、功能鑒定、蛋白互作等方面深入研究石榴SPS、INV基因家族成員調控蔗糖代謝的分子機理。
基金項目:棗莊學院山東省石榴精深加工工程技術研究中心/山東省石榴資源綜合開發工程實驗室開放課題(SLKF2021001);山東省重點研發計劃項目(2022TZXD009);山東省農業科學院農業科技創新工程項目(CXGC2023A12);山東省農業科學院揭榜科技難題項目(SHJB2022-42)
