基于線粒體D-loop序列的江蘇克氏原螯蝦養殖群體遺傳多樣性分析
2024-07-03 21:23:41魯蘇皖徐宇李佳佳高志寶嚴維輝許志強王興強李旭光
江蘇農業科學 2024年10期
魯蘇皖 徐宇 李佳佳 高志寶 嚴維輝 許志強 王興強 李旭光
doi:10.15889/j.issn.1002-1302.2024.10.026
摘要:江蘇是我國克氏原螯蝦的主產區,近年來繁養模式不斷變化,多代繁養后的遺傳多樣性變化情況尚未可知,以及江蘇地區克氏原螯蝦養殖種群的群體遺傳學特征數據尚不清楚。為研究當前江蘇主產區克氏原螯蝦養殖群體的遺傳多樣性特征,對來自江蘇南京、徐州、連云港、揚州、淮安、無錫、泰州興化和宿遷泗洪的8個克氏原螯蝦養殖群體樣本的線粒體控制區D-loop基因片段進行PCR擴增和測序分析,分析了不同養殖群體的遺傳結構特點。共測序獲得了8個群體223個樣本的線粒體D-loop序列,比對長度包括863個位點,其中,變異位點95個,簡約信息位點25個。基因序列中AT含量(80.01%)高于CG含量(19.99%),表現出較為明顯的堿基組成偏倚性。223個樣本共定義了37種單倍型,其中,單倍型Hap 1為8個群體的共有單倍型。泗洪群體獨有的單倍型最多,泰州興化群體獨有的單倍型最少。本研究所涉及的養殖群體總體單倍型多樣性指數(Hd)為0.39,總體核苷酸多樣性指數(Pi)為0.003 37,遺傳多樣性處于較低水平。其中,宿遷泗洪群體的遺傳多樣性指數最高(Hd為0.632),興化群體的遺傳多樣性指數最低(Hd為0.228)。不同群體間的遺傳距離總體較小,其中,南京群體與其他群體間的遺傳距離相對較大(0.005),提示南京群體與其他群體間的基因交流較少。8個養殖群體間的遺傳分化系數為-0.219 6~0.317 33,徐州、揚州、無錫及連云港群體間的遺傳分化系數為負值,表明這些群體間存在較為頻繁的基因交流。分子方差分析(A-MOVA)結果顯示,95.35%的遺傳變異來自于群體內,群體間的遺傳變異較少(4.65%)。最小跨度網絡圖和系統發育分析均未顯示出單倍型與不同群體的對應關系,提示當前江蘇克氏原螯蝦養殖群體間的基因交流較為頻繁。
關鍵詞:克氏原螯蝦(Procambarus clarkii);江蘇地區;遺傳多樣性;遺傳結構
中圖分類號:S968.22? 文獻標志碼:A
文章編號:1002-1302(2024)10-0192-05
收稿日期:2023-07-21
基金項目:江蘇省種業振興“揭榜掛帥”項目(編號:JBGS[2021]032);江蘇現代農業(克氏原螯蝦)產業技術體系建設項目(編號:JATS[2022]411);江蘇省農業重大新品種創制項目(編號:PZCZ201746)。
作者簡介:魯蘇皖(1997— ),男,江蘇宿遷人,碩士研究生,主要從事甲殼動物增養殖研究。E-mail:136502596@qq.com。
通信作者:李旭光,博士,研究員,主要從事水生甲殼類繁育和生態養殖研究。E-mail:xuguangli1981@163.com。
克氏原螯蝦(Procambarus clarkii)俗稱小龍蝦,屬于節肢動物門甲殼亞目軟甲綱十足目螯蝦科,原產于美洲,20世紀30年代從日本引入我國南京,2021年,我國克氏原螯蝦養殖面積173萬hm2、產量263.36萬t,現已成為我國最重要的水產經濟物種之一[1]。長期以來,國內克氏原螯蝦養殖過程中普遍采用繁養一體的繁育方式、捕大留小的捕撈手段,捕撈剩下的小個體成蝦留塘自繁自養,繁養過程中的“逆向選擇”問題較為嚴重。水產養殖群體的遺傳多樣性評估對良種選育和資源保護等方面具有重要意義。王長忠等對長江中下游地區野生的克氏原螯蝦的遺傳多樣性進行了本底調查,研究表明4個克氏原螯蝦群體的遺傳多樣性處于中等水平[2]。曹玲亮使用了微衛星標記分析了安徽3個水系中野生克氏原螯蝦群體的遺傳多樣性和遺傳結構,發現調查水域中克氏原螯蝦群體的遺傳多樣性處于較高水平,但多個群體存在瓶頸效應[3]。彭剛對安徽地區的克氏原螯蝦養殖和野生群體進行了遺傳多樣性分析,發現安徽地區克氏原螯蝦人工養殖群體具有較高的遺傳多樣性[4]。邢智珺對江蘇地區克氏原螯蝦野生群體進行了群體遺傳學特征研究,發現江蘇克氏原螯蝦野生群體遺傳多樣性較高,不同群體間存在中度遺傳分化[5]。而有關江蘇地區克氏原螯蝦養殖群體的遺傳多樣性鮮有報道。
線粒體D-loop區缺乏編碼的壓力,是線粒體基因中變異速率最快的區域,常被用來研究種群間及種群內的遺傳分化。江蘇是我國克氏原螯蝦產業的發源地,養殖時間長,養殖業發展規模大,同樣存在繁養過程中“逆向選擇”的問題,目前產生的種質資源退化問題與遺傳多樣性之間的關系尚不明確。本研究以江蘇克氏原螯蝦主要養殖地區群體作為研究對象,基于線粒體D-loop區序列變異分析研究了多個養殖群體的群體遺傳學特征,以期為克氏原螯蝦良種選育及合理選擇繁養模式提供參考依據。
1? 材料與方法
1.1? 樣本采集
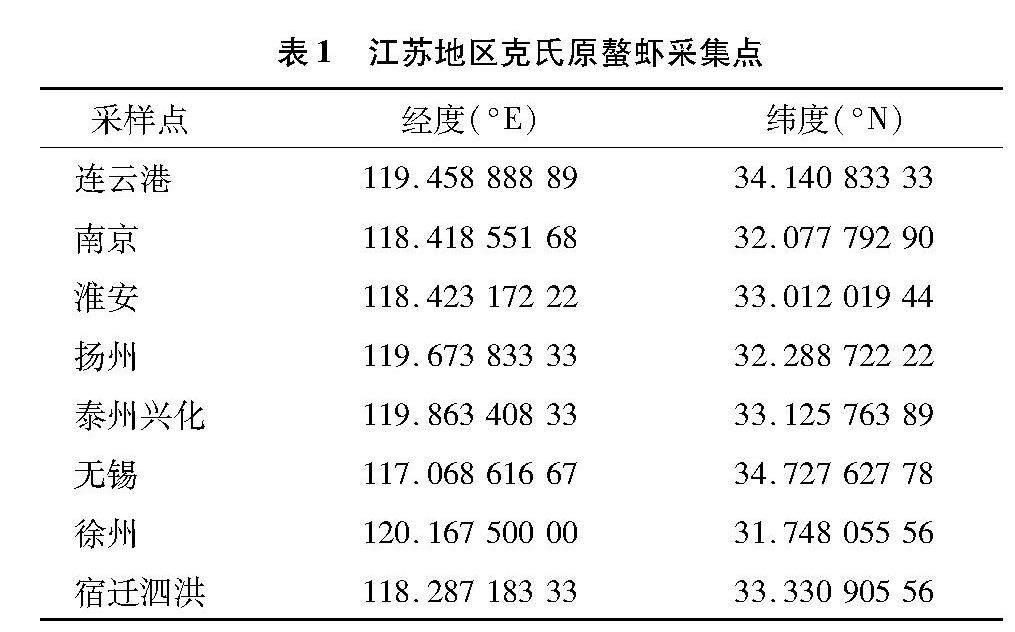
克氏原螯蝦樣本于2021年5—9月收集于無錫、南京、揚州、淮安、泰州興化、宿遷泗洪、連云港、徐州8個地區的養殖群體(表1),采樣地為江蘇地區克氏原螯蝦養殖主要產區。每個地區樣品采集量30尾以上,共計260尾,所有樣品均活體運輸至實驗室后,取尾部肌肉保存于-20 ℃備用。
1.2? 基因組DNA提取
采用TIANGEN公司的基因組DNA提取試劑盒(DP304)提取不同樣本的基因組DNA,使用核酸蛋白儀(TGem Pro 分光光度計)測定DNA濃度,使用1%瓊脂糖凝膠電泳檢測DNA質量。
1.3? PCR擴增、測序
參照克氏原螯蝦線粒體序列信息(JAIWQB 000000000.1)設計引物用于擴增D-loop區序列,引物序列為:Crayfish DLOOP-F(5′-TCGCTGTAAAGTTGAAGAAGTT-3′)和Crayfish DLOOP-R(5′-TTAATCTCTTCATATCTTTAATTAC-3′)引物由上海生工生物公司合成。PCR反應體系為50 μL,其中DNA模版6 μL,正反引物(10 μmoL/L)各2 μL,mix 25 μL(諾唯贊,南京),ddH2O 15 μL。PCR反應程序為:98 ℃預變性30 s;98 ℃變性10 s,48.5 ℃退火5 s,72 ℃延伸1 s,32個循環;72 ℃延伸1 min。PCR產物經1%瓊脂糖凝膠電泳檢測,條帶單一且明亮,送南京生工生物有限公司進行Sanger雙向測序。
1.4? 數據分析
測序所得線粒體D-loop區序列,使用SeqMan軟件進行手動拼接。匯總后使用MEGA 7軟件對齊后手動校對。計算序列堿基組成,變異點位數及群體間遺傳距離等信息,并基于Neis遺傳距離構建UPGMA進化樹。使用DnaSP軟件計算序列的單倍型數(H)、群體的單倍型多樣性(Hd)、核苷酸多樣性(Pi)、平均核苷酸差異數(K)、中性測試Tajimas D。使用Arlequin 3.5軟件計算群體間遺傳分化指數(Fst),并進行分子方差分析(AMOVA)。
2? 結果與分析
2.1? 序列特征與遺傳多樣性
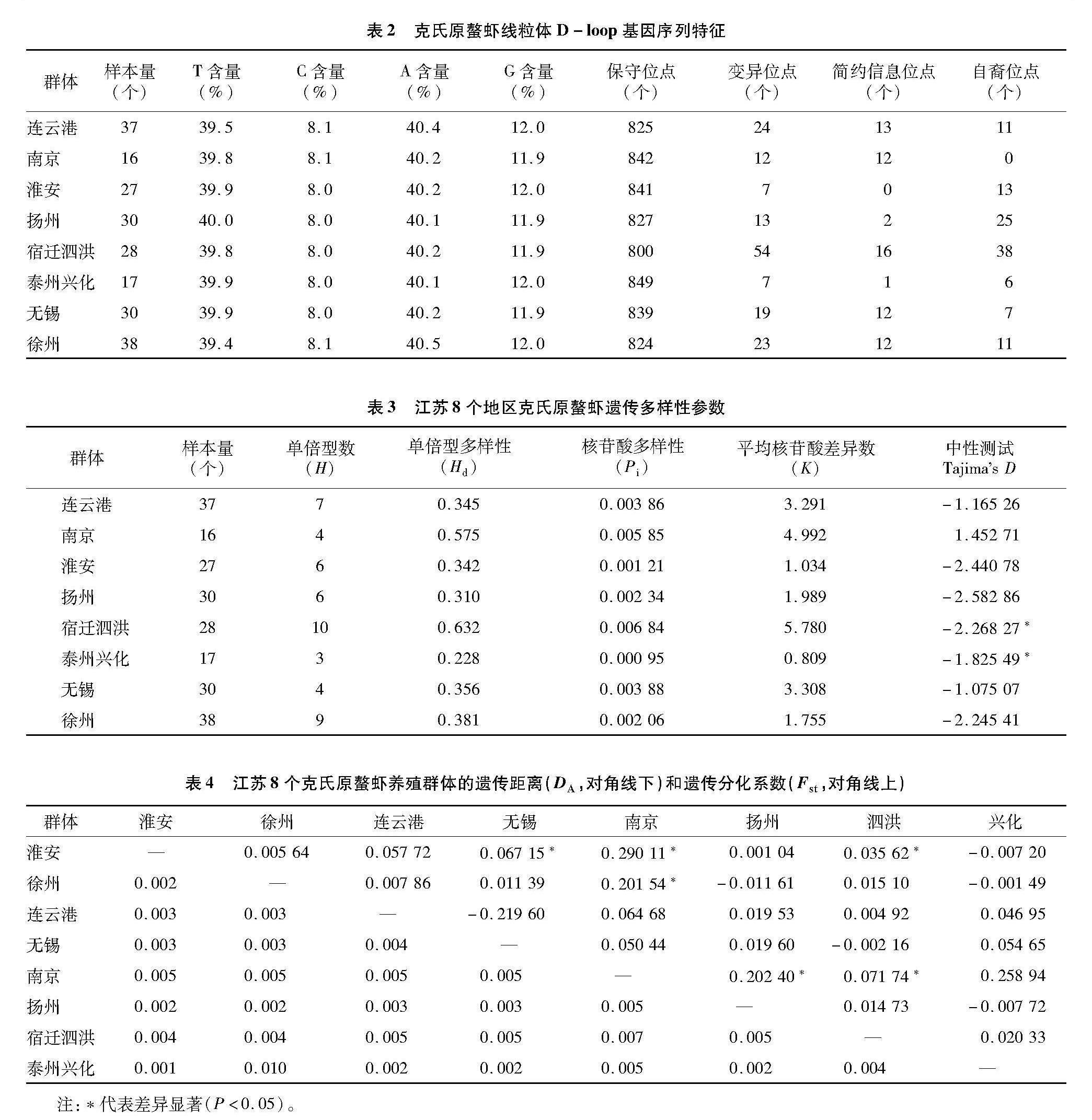
由表2可知,8個群體共260尾的克氏原螯蝦經過基因組DNA提取、PCR擴增、測序后共獲得223條線粒體D-loop基因序列,經過拼接、排序、校對后,獲得全長為863 bp的序列片段,CG含量20.1%。863個核苷酸位點中共有保守位點763個(占88.41%)、突變位點95個、簡約信息位點25個、自裔位點70個。
223條序列中共檢出37種單倍型。其中,Hap 1為8個群體共有單倍型。宿遷泗洪群體獨有的單倍型最多,徐州群體其次,泰州興化獨有的單倍型最少。單倍型Hap 1被173個個體共享,包含個體最多;其次Hap 12,被11個個體共享。
基于克氏原螯蝦線粒體D-loop區序列,在8個養殖群體中共定義了37個單倍型。由表3可知,單倍型Hap1分布最廣,在8個地區中均有分布,宿遷泗洪地區存在的單倍型數最多,單倍型多樣性最高(Hd=0.632),核苷酸多樣性也最高(Pi=0.00 684)。泰州興化地區存在的單倍型數最少(H=3),單倍型多樣性(Hd=0.228)最低。本研究中涉及到的8個養殖群體的總體單倍型多樣性指數為0.39,江蘇克氏原螯蝦養殖群體遺傳多樣性整體處于較低水平。
2.2? 種群遺傳結果
由表4可知,江蘇地區8個克氏原螯蝦群體間的遺傳分化系數Fst為-0.219 60~0.317 33,其中,在無錫地區與連云港地區和宿遷泗洪地區、揚州地區與徐州地區和泰州興化地區、泰州興化地區和淮安地區、徐州地區和揚州地區這幾個地區間遺傳分化系數為負值,表明這幾個地區間并未出現顯著的遺傳分化。南京地區的遺傳分化系數顯著高于其他地區。揚州地區的遺傳分化系數相對低于其他地區。不同群體間的遺傳距離總體較小,其中,南京群體與其他群體間的遺傳距離相對較大(DA=0.005),表明南京群體與其他群體間的基因交流較少。
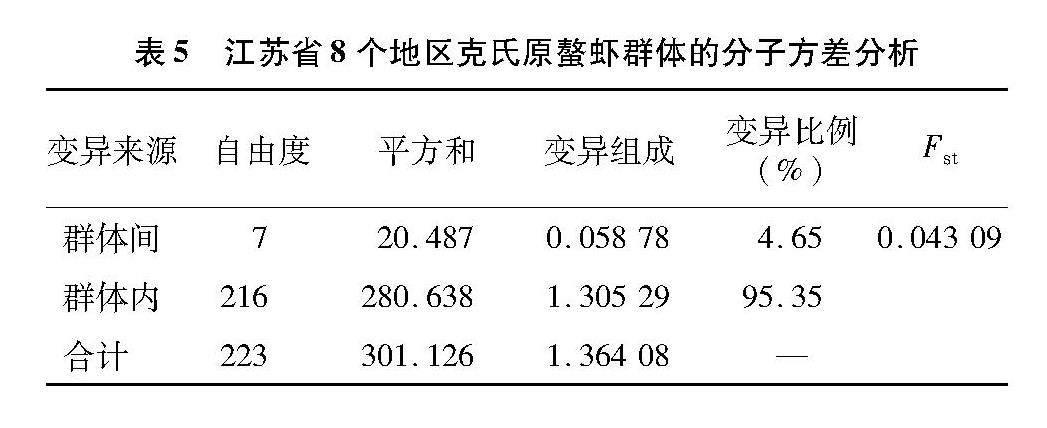
由AMOVA分析結果(表5)可知,總Fst為0.043 09,變異不顯著,其中,95.35%遺傳變異來自于群體內,僅有4.65%來自于群體間。
2.3? 單倍型系統進化樹與網絡圖
由圖1可知,基于Neis遺傳距離對江蘇地區克氏原螯蝦養殖群體構建UPGMA進化樹,8個群體可分為4組,分別為:淮安地區、泰州興化地區和無錫地區為一組,連云港地區、徐州地區和揚州地區為一組;宿遷泗洪地區和南京地區各為一組。
由使用Network軟件生成的單倍型網絡圖(圖2)可知,37種單倍型間關系中,Hap1單倍型位于網絡關系圖的中心位置,為8個地區共有的單倍型,可預測這一單倍型相對較為原始,周圍的單倍型由其衍生而來。不同的單倍型在各地區中均有分布,無明顯的譜系分化。
3? 討論
遺傳多樣性是物種對變化的環境不斷適應的結果,物種為適應環境變化會不斷進化自身,從而產生豐富的基因型[6-7]。本研究中江蘇地區克氏原螯蝦養殖群體的單倍型多樣性平均值為0.396,核苷酸多樣性平均值為0.003 37。與同樣基于線粒體D-loop區序列分析的其他甲殼類動物相比,江蘇克氏原螯蝦養殖群體的遺傳多樣性低于三疣梭子蟹種群(Hd=0.927,Pi=0.024 3),低于日本沼蝦種群(Hd=0.816,Pi=0.019 15)[8-9],研究發現,江蘇各克氏原螯蝦養殖群體的遺傳多樣性無明顯差異,遺傳多樣性整體處于較低水平。與山東克氏原螯蝦野生群體相比(Hd=0.204,Pi=0.003 6),江蘇養殖群體的單倍型多樣性較高[10]。與安徽滁州地區克氏原螯蝦養殖群體的遺傳多樣性水平(Hd=0.373 14,Pi=0.001 72)[11]相近。本研究中,江蘇地區克氏原螯蝦遺傳多樣性整體不高,造成原因諸多。首先,采樣地區的克氏原螯蝦苗種多來自池塘自繁,池塘內親蝦存在繁殖期內1個雌體和多個雄體交配的現象,會導致種質資源的退化[12]。其次,江蘇地區作為克氏原螯蝦的主要養殖地區,因捕撈壓力逐年增加,導致稀有位點丟失也可能是江蘇地區的克氏原螯蝦整體遺傳多樣性水平不高的原因之一[13]。泰州興化群體的單倍型多樣性(Hd=0.228)和核苷酸多樣性(Pi=0.000 95)均最低。泰州興化地區多采用蝦蟹混養模式養殖克氏原螯蝦,在每年4月初向河蟹養殖塘中投放約10尾的抱籽親蝦用于苗種繁殖,5月下旬至6月中旬將蝦蟹混養塘中的克氏原螯蝦盡可能捕盡,盡量減少對河蟹生長影響[14],這種繁養模式可能導致了興化群體較低的遺傳多樣性。研究發現,宿遷泗洪地區的單倍型多樣性水平和核苷酸多樣性水平最高。宿遷泗洪地區臨近洪澤湖,可能由于養殖群體與洪澤湖內野生群體交流頻繁,從而導致了宿遷泗洪群體較高的遺傳多樣性。本研究中宿遷泗洪地區遺傳多樣性較高與高揚等的研究結果[15]相一致。遺傳距離是判斷群體之間親緣關系的關鍵指標,遺傳距離越大表明群體間親緣關系越遠[16]。遺傳距離較近和遺傳分化系數的負值,表明江蘇地區的克氏原螯蝦存在近親繁殖、群體間的分化程度較低等問題。江蘇省不同地區間盲目引種,是導致遺傳距離較近的原因之一。遺傳結構是判斷新物種適應棲息地能力的主要判斷標準[17]。本研究克氏原螯蝦8個群體中共發現37種單倍型,其中,單倍型Hap1在各群體中占比最高,在網絡圖中也處于中心位置。從而推斷單倍型Hap1是克氏原螯蝦群體中穩定的優勢單倍型。從單倍型網絡圖中發現,以單倍型Hap1為中心呈放射狀散開,各群體間并未產生明顯遺傳分化,各群體間的遺傳分化水平整體較低。本研究AMOVA結果顯示,有95.35%的變異發生在群體內,僅有4.65%的變異發生在群體間,這與刑智珺等研究中克氏原螯蝦的變異多發生在群體內,僅有極少數發生在群體間的結果[5,18-19]相一致。
江蘇地區克氏原螯蝦養殖群體的遺傳多樣性整體處于較低水平,建議及時對江蘇克氏原螯蝦養殖群體進行遺傳改良,以提高克氏原螯蝦養殖群體的遺傳多樣性,同時盡快改變自繁自育的苗種繁育和養殖模式。
參考文獻:
[1]王亞民,曹文宣. 中國水生外來入侵物種對策研究[J]. 農業環境科學學報,2006,25(1):7-13.
[2]王長忠,李? 忠,梁宏偉,等. 長江下游地區4個克氏原螯蝦群體的遺傳多樣性分析[J]. 生物多樣性,2009,17(5):518-523.
[3]曹玲亮,周立志,張保衛. 安徽三大水系入侵物種克氏原螯蝦的種群遺傳格局[J]. 生物多樣性,2010,18(4):398-407.
[4]彭? 剛,劉偉杰,李佳佳,等. 長江流域3個克氏原螯蝦野生群體遺傳結構的微衛星分析[J]. 江蘇農業學報,2010,26(5):1115-1117.
[5]邢智珺,姜虎成,陸? 偉,等. 江蘇8個克氏原螯蝦群體遺傳多樣性微衛星分析[J]. 上海海洋大學學報,2014,23(5):656-662.
[6]Vrijenhoek R C. Genetic diversity and fitness in small populations[M]//Conservation genetics. Basel:Birkhuser,1994:37-53.
[7]李大命,李? 康,張彤晴,等. 洪澤湖大銀魚(Protosalanx hyalocranius)cytb和COⅠ基因序列多態性分析[J]. 漁業科學進展,2017,38(6):25-31.
[8]董志國,李曉英,王普力,等. 基于線粒體D-loop基因的中國海三疣梭子蟹遺傳多樣性與遺傳分化研究[J]. 水產學報,2013,37(9):1304-1312.
[9]董新培,武小斌,萬海付,等. 河北3個日本沼蝦野生群體線粒體DNA D-Loop基因序列變異及種群遺傳結構分析[J]. 水產學報,2017,41(2):182-188.
[10]楊? 玲,李? 寧,朱樹人,等. 山東省3個克氏原螯蝦地理群體線粒體COⅠ基因的序列差異分析[J]. 安徽農業科學,2015,43(20):41-44.
[11]余紅喜,劉? 帆,朱國美,等. 基于線粒體COⅠ基因滁州地區克氏原螯蝦養殖群體遺傳多樣性分析[J]. 安徽農業科學,2022,50(21):111-113,125.
[12]Villanelli F,Gherardi F. Breeding in the crayfish,Austropotamobius pallipes:mating patterns,mate choice and intermale competition[J]. Freshwater Biology,1998,40(2):305-315.
[13]夏愛軍,唐建清. 克氏原螯蝦現狀分析與研究思路[J]. 水產養殖,2006,27(6):9-11.
[14]張鳳翔,陳? 杰,徐美香,等. 蝦蟹混養模式的創新試驗與思考[J]. 科學養魚,2020(2):32-34.
[15]高? 楊,田? 燦,姜京京,等. 基于微衛星標記的克氏原螯蝦種群遺傳多樣性和遺傳結構分析[J]. 江蘇農業科學,2023,51(5):191-199.
[16]張鶴千,楊子拓,李桂峰,等. 珠江流域野生黃顙魚Pelteobagrus fulvidraco的Cytb基因序列分析[J]. 中山大學學報(自然科學版),2015,54(5):102-108.
[17]Bazin E,Glémin S,Galtier N. Population size does not influence mitochondrial genetic diversity in animals[J]. Science,2006,312(5773):570-572.
[18]崔文濤,鄒宇凡,白志毅,等. 安徽地區克氏原螯蝦群體的遺傳多樣性和遺傳結構分析[J]. 水產學報,2022,10(28):1-10.
[19]Yi S K,Li Y H,Shi L L,et al. Characterization of population genetic structure of red swamp crayfish,Procambarus clarkii,in China[J]. Scientific Reports,2018,8:5586.
