
不同生境水曲柳種子的形態結構及抗旱性分析
2024-07-10 06:54:16韓春麗,衛星,韋慶鈺,賈紅濤
森林工程 2024年3期
韓春麗,衛星,韋慶鈺,賈紅濤
摘要:為選育水曲柳耐干旱品種提供理論依據,選取不同生境水曲柳(Fraxinus mandshurica)種子為試驗材料,對比分析不同生境下水曲柳種子的形態結構特征和吸水特性,采用不同質量濃度的聚乙二醇(Polyethylene glycol,PEG)溶液對水曲柳種子進行干旱脅迫,比較分析不同生境水曲柳種子在干旱環境下的萌發表現。研究結果顯示,1)半干旱生境水曲柳的種子和種翅大小、千粒質量及種皮厚度等形態結構均顯著大于適生生境種子;2)半干旱生境種子萌發時的吸水率顯著低于適生生境種子7.75%;3)當PEG溶液質量濃度為10%時,半干旱生境水曲柳種子的發芽率、發芽指數、活力指數以及胚根長度開始顯著高于適生生境種子。對發芽率進行線性回歸,半干旱生境水曲柳種子的耐旱極限質量濃度是適生生境種子的1.23倍,具有更強的耐旱能力;4)干旱脅迫下,兩生境水曲柳種子的過氧化氫酶、過氧化物酶、超氧化物歧化酶活性均表現出先上升后下降的趨勢,且半干旱生境種子的酶活性始終高于適生生境。此研究結果表明在干旱環境造林選種時,可優先選用半干旱生境水曲柳種子作為造林種源,以提高種子萌發率和苗木成活率,為水曲柳資源擴大提供保障。
關鍵詞:水曲柳;種子萌發;形態結構;吸水特性;干旱脅迫
中圖分類號:S722.5文獻標識碼:A文章編號:1006-8023(2024)03-0088-13
Analysis of Morphological Structure and Drought Resistance?of Fraxinus mandshurica Seeds in Different Habitats
HAN Chunli1,? WEI Xing1,2*,? WEI Qingyu1, JIA Hongtao1
(1.College of Forestry, Northeast Forestry University, Harbin 150040, China;2.Key Laboratory of Sustainable Forest Ecosystem Management (Northeast Forestry University),?Ministry of Education, Harbin 150040, China)
Abstract:To provide a theoretical basis for selecting drought resistant varieties of Fraxinus mandshurica, this article selected Fraxinus mandshurica seeds from different habitats as experimental materials, and compared and analyzed the morphological and structural characteristics and water absorption characteristics of Fraxinus mandshurica seeds in different habitats. Different mass concentrations of polyethylene glycol (PEG) solution was used to treat drought stress on the seed of Fraxinus mandshurica, and the germination performance of Fraxinus mandshurica seeds in different habitats under drought conditions was compared and analyzed. The research results showed that, 1) the seed and wing size, thousand grain mass, and seed coat thickness of Fraxinus mandshurica in semi-arid habitats were significantly larger than those in suitable habitats. 2) The water absorption rate of seeds during germination in semi-arid habitats was significantly lower than that of seeds in suitable habitats by 7.75%. 3) When the mass concentration of PEG solution was 10%, the germination rate, germination index, vitality index, and radicle length of Fraxinus mandshurica seeds in semi-arid habitats began to be significantly higher than those in suitable habitats. Linear regression was performed on the germination rate, and the drought tolerance limit mass concentration of Fraxinus mandshurica seeds in semi-arid habitats was 1.23 times that of seeds in suitable habitats, indicating stronger drought resistance. 4) Under drought stress, the activities of catalase, peroxidase, and superoxide dismutase of Fraxinus mandshurica seeds in both habitats showed a trend of first increasing and then decreasing, and the enzyme activities of seeds in semi-arid habitats were consistently higher than those in suitable habitats. The results of this study indicate that in the selection of afforestation species in arid environments, priority should be given to selecting the seeds of Fraxinus mandshurica from semi-arid habitats as the afforestation source, in order to improve seed germination rate and seedling survival rate, and provide guarantees for the expansion of Fraxinus mandshurica resources.
Keywords:Fraxinus mandshurica; seed germination; morphological structure; water-absorbing character; drought stress
0引言
水分是影響植物生長和發育的關鍵環境因子[1]。目前,隨著全球氣候變暖,土壤水分減少呈逐年增加的趨勢,研究植物在水分虧缺環境下的生長策略對保護植物資源、促進植物生長具有重要的意義[2]。種子作為新生命階段的開端,如何在各類非生物逆境脅迫下順利完成萌發并建成幼苗茁壯成長,是植物分子生物學和生態學領域的重要研究方向和熱點之一。植物生態學、植物發育學以及遺傳學方面的相關研究表明,長期處于逆境中的植物具有更強的環境適應結構、生理特性[3]和生存力[4]。隨著地理和氣候因子的不斷變化,處在不同生境中的植物會進化出相應策略來適應復雜多變的環境[5-6]。為了種子能夠更好地萌發,通過調節生理代謝和提高保護酶等一系列活動來幫助其更好地適應水分虧缺的環境,在形態結構[7]和生理特性[8]上會產生一定的變異。抗性強的種質能夠更好地適應逆境環境,開發優質的抗性種質資源,對提高植物抗性、擴大植物栽植范圍具有重要的意義。
水曲柳(Fraxinus mandshurica)是木犀科梣屬落葉大喬木,國家Ⅱ級保護樹種,具有重要的生態和經濟價值。近年來,國內外學者對水曲柳種子的研究都集中于種子引發、提高種子適應性等機制,閆月等[9]使用25%的PEG溶液對種子進行3 d的引發處理,可顯著提高其發芽率;但在長時間或者高質量濃度引發處理下則對種子不利,可以證明水曲柳種子具有一定的抗旱性。20世紀80年代,為增加東北地區珍貴樹種資源儲備,擴大水曲柳栽植面積,我國在東北地區松嫩平原半干旱地區栽植了大面積水曲柳人工林。目前林分已經成熟,進入結實旺盛期。研究證實來自不同種源產生的水曲柳種子在長度、寬度以及千粒質量方面具有顯著差異[10],但若使水曲柳種子長期處在干旱環境下,其形態結構及萌發特性是否有變化尚不清楚,因此,本研究分析討論不同生境的水曲柳種子對干旱環境的適應性差異,進一步探索逆境植物種子的潛在價值,為篩選水曲柳抗逆品種、擴大水曲柳種植范圍提供理論依據。
1材料與方法
1.1種子采集與試驗地概況
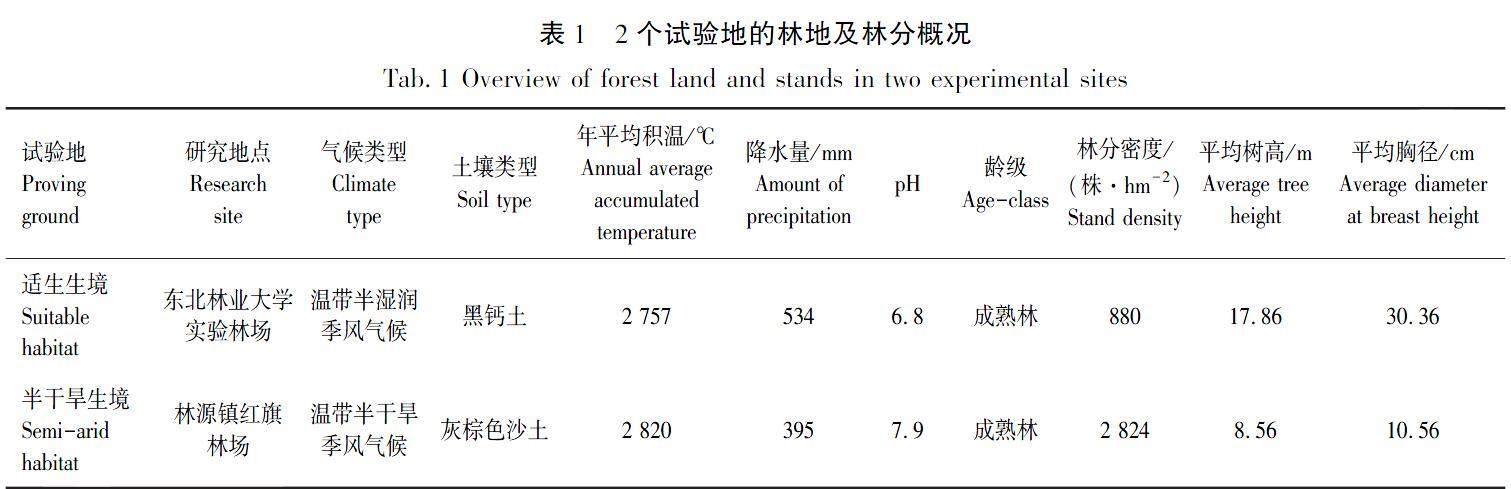
水曲柳種子采集于2022年10月初,分別采自黑龍江省大慶市大同區林源鎮紅旗林場(125°19′ E,46°23′ N)和哈爾濱市東北林業大學實驗林場(126°37′ E,45°43′ N),其中東北林業大學實驗林場位于長白山北部山地,是水曲柳人工林分布最多、生產力最高的中心產區[11]。各試驗地分別選取3株樹干通直、生長健康、無病蟲害、達到正常開花結實的母樹進行種子采集,并將采集下的種子充分混勻備用。本試驗中2個試驗地的林地概況見表1[12]。
1.2試驗方法
1.2.1種實表型特征
水曲柳種翅及種子的長度、寬度、厚度:將各生境下采集的水曲柳種子分別充分混勻,各隨機選取20粒種子,使用游標卡尺測定各指標,取其平均值。
種子千粒質量:將各生境下采集的水曲柳種子分別充分混勻,各隨機選取100粒種子,使用萬分之一天平秤稱質量(精確至0.001 g),重復8次,用百粒法計算種子千粒質量[13]。
種翅和種子面積:使用體視顯微鏡(SZX-ILLB2-200)對種翅及種子拍照,用數字化掃描儀(EPSON,Expression 10000XL 1.0)掃描種翅的面積,計算翅載力。
1.2.2種子解剖結構特征
種皮結構:分別從不同生境中取完整飽滿成熟的水曲柳種子各20粒,用蒸餾水浸泡24 h,分離出種皮和胚。采用常規石蠟切片法制作種皮切片[14],將保存好的切片用電子顯微鏡(OLYMPUS BX-51)觀察并拍攝水曲柳種子橫切面,測量對比種皮厚度。采用掃描電子顯微鏡(日本JSM-7500F)觀察上種皮特征[15]。
種皮透性:隨機選取不同生境水曲柳種子各30粒風干后稱質量(精確至0.001 g),重復3次;之后將種子分別放入蒸餾水中浸泡,每隔8 h稱一次,直至吸脹至飽滿至恒重。分別計算各生境種子的吸水率和吸水速率[16]并記錄初始萌發時間。
1.2.3種子萌發指標測定
使用聚乙二醇(Polyethylene glycol, PEG-6000)溶液模擬干旱脅迫,PEG溶液梯度設置為0%、5%、10%、15%、20%、25%,對應的滲透勢分別是0、0.05、0.20、0.40、0.60、0.86 MPa。以無菌蒸餾水(CK)為對照溶液。將處理好的種子置于直徑為12 cm的培養皿中,培養皿的底部放兩層濾紙并滴入6 mL相應質量濃度的PEG溶液,做好標記后置于10? ℃的培養箱黑暗條件下萌發。每個處理隨機選取30粒種子,重復3次。
種子萌發過程中,以種子胚根突破種皮2 mm為萌發標志,每天定時觀察發芽數量并做好試驗記錄。試驗結束后計算種子的發芽率、發芽勢、發芽指數、活力指數和平均發芽時間,以發芽結束后的發芽率為因變量(y)、以PEG溶液為自變量(x)建立回歸方程,并計算種子耐旱適宜質量濃度(發芽率達到對照發芽率75%時相對應的質量濃度)、耐旱半致死質量濃度(發芽率達到對照發芽率50%時對應的質量濃度)和耐旱極限質量濃度(發芽率達到對照發芽率10%時相對應的質量濃度)[17]。種子萌發結束后每培養皿選取9粒長勢相同的種子,測量其主根長和子葉長,測量后用錫紙包裹后置于液氮中保存備用,用于各生境種子生理指標測定。
測定生理指標時,從液氮中取出樣本,稱取0.1 g進行冰浴研磨,利用分光光度計法[18]測定種子的過氧化氫酶(CAT)、過氧化物酶(POD)和超氧化物歧化酶(SOD),每處理重復3次,
1.3數據分析
用Microsoft Excel 2020和SPSS 26.0統計軟件進行數據整理與分析,采用獨立樣本T檢驗法分析兩生境水曲柳種子各指標間的差異顯著性,利用Origin 2022軟件進行繪圖。本研究圖表中的所有數據均采用平均值±標準誤差表示。
2結果與分析
2.1不同生境水曲柳種實的表型特征差異
兩生境水曲柳種翅成熟時均為黃褐色,呈倒卵狀披針形,中部最寬。與半干旱生境種翅相比,適生生境水曲柳種翅底端的扭曲程度更深,如圖1(a)所示。除種翅厚度外,半干旱生境種翅長度、寬度、重量和翅載力均顯著高于適生生境(P<0.05),見表2,分別為適生生境種翅的1.06、1.11、1.41、1.29倍。兩生境種子顏色形態基本相似,均為深棕褐色,形狀為倒卵形,中部寬兩側窄,如圖1(b)所示。半干旱生境水曲柳種子的長度、寬度、厚度以及千粒質量都顯著大于適生生境水曲柳種子(P<0.05),見表3,分別是適生生境種子的1.12、1.11、1.12、1.56倍。
2.2不同生境水曲柳種子的解剖結構差異
不同生境水曲柳種子上表皮細胞排列差異顯著,如圖2(a)所示。半干旱生境種子上表皮細胞大小不均勻,平均細胞表面積比適生生境種子小12.65%,但細胞行間排列比適生生境種子松散,是適生生境種子的1.84倍,單位面積內細胞數量僅為17個。
種子上表皮與胚乳連接,胚乳內富含大量蛋白質等營養物質,如圖2(b)所示。半干旱生境水曲柳種子的種皮厚度和胚乳厚度均顯著大于適生生境水曲柳種子(P<0.05),見表4,分別為適生生境的1.24倍和1.04倍。兩生境水曲柳種胚結構相同,如圖2(c)所示,除胚軸寬度外,半干旱生境種胚的子葉長度、胚軸長度及胚率均顯著大于適生生境(P<0.05),見表4,分別為適生生境的1.12、1.15、1.05倍。
2.3不同生境水曲柳種子的吸水特性差異
不同生境水曲柳種子的吸水率隨著吸水時間的延長而增大。半干旱生境水曲柳種子和適生生境種子的吸水率在各個時間段均無顯著性差異,但是半干旱生境種子吸水時的初始萌發時間(胚根露出1 mm時即為初始萌發)比適生生境種子早16 h,且半干旱生境種子初始萌發時的吸水率顯著低于適生生境種子7.75%,如圖3(a)所示。兩生境種子的吸水速率隨著吸水時間的延長逐漸降低,半干旱生境水曲柳種子的吸水速率在吸水32 h后顯著高于適生生境7.77%~8.42%,如圖3(b)所示。
2.4不同生境水曲柳種子的耐旱性差異
隨著干旱脅迫強度的增加,不同生境水曲柳種子的發芽率均逐漸下降,如圖4(a)所示。干旱脅迫下,半干旱生境水曲柳種子的發芽率始終高于適生生境。當PEG質量濃度為10%時,半干旱生境水曲柳種子的發芽率開始顯著高于適生生境種子,且隨干旱脅迫強度的增強,二者差異越大,最大差異達到61.57%。發芽勢的變化趨勢與發芽率基本相似,如圖4(b)所示,半干旱生境水曲柳種子的發芽勢在PEG質量濃度為10%和15%時顯著高于適生生境種子,分別高出55.98%和66.69%。隨著干旱脅迫強度的增強,兩生境水曲柳種子的平均發芽時間逐漸延長,如圖4(c)所示。當PEG質量濃度為5%~15%時,半干旱生境水曲柳種子的平均發芽時間均顯著短于適生生境種子8.21%~11.06%。根據兩生境水曲柳種子的發芽率進行線性回歸,如圖4(d)所示,半干旱生境水曲柳種子的耐旱適宜質量濃度、耐旱半致死質量濃度及耐旱極限質量濃度分別是適生生境種子的1.26、1.24、1.23倍。
隨著干旱脅迫程度的增強,不同生境水曲柳種子的發芽指數和活力指數均逐漸下降,如圖5所示。干旱脅迫下,半干旱生境水曲柳種子的發芽指數和活力指數始終高于適生生境種子。當PEG質量濃度為10%時,半干旱生境水曲柳種子的發芽指數和活力指數開始顯著高于適生生境種子,最大差異分別達到64.79%和115.69%。
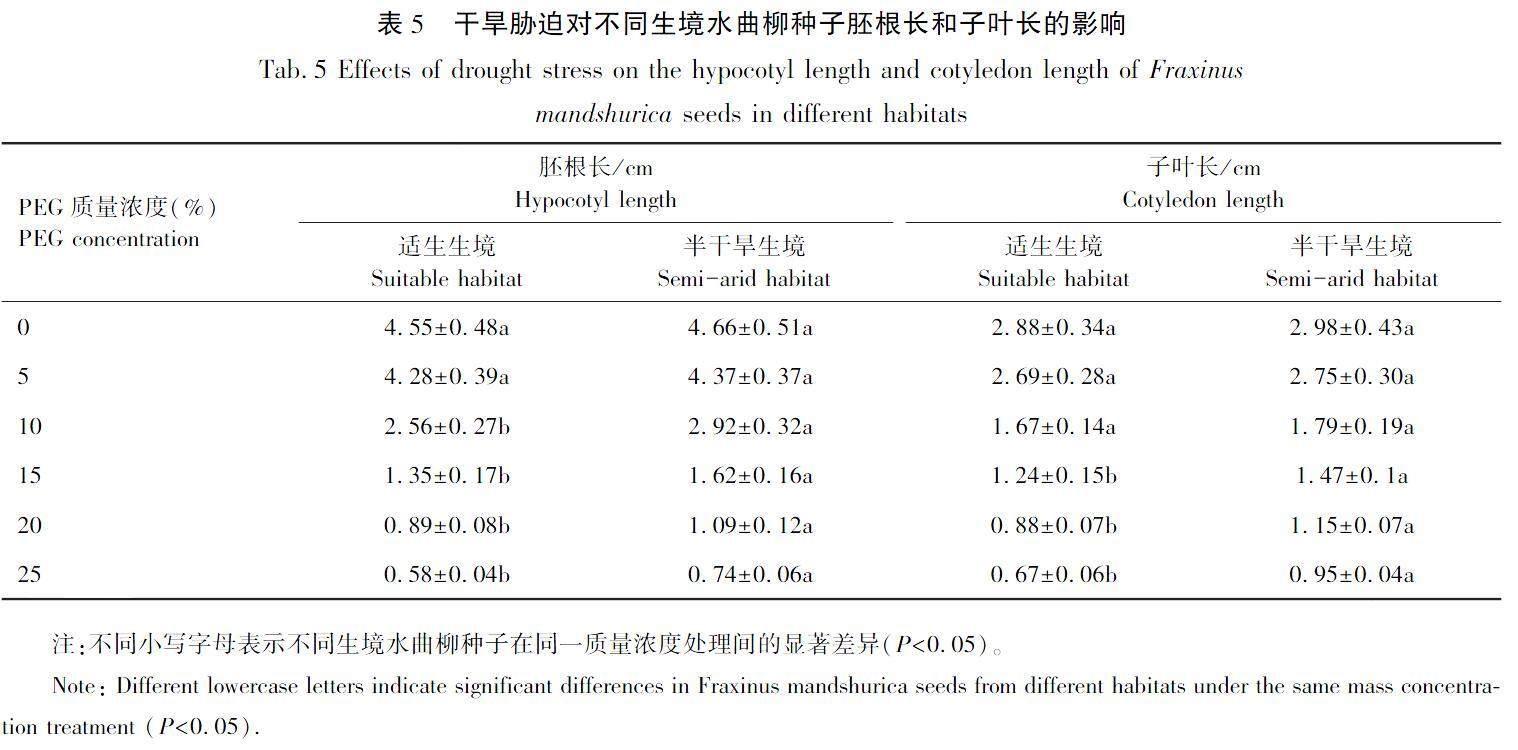
隨著干旱脅迫強度的增加,不同生境水曲柳種子的胚根長和子葉長均逐漸減小,如圖6和表5所示。當PEG質量濃度為10%時,半干旱生境水曲柳種子的胚根長度開始顯著高于適生生境種子,當PEG質量濃度為15%時,半干旱生境水曲柳種子的子葉長度開始顯著高于適生生境種子,兩生境種子的胚根最大差異達到27.59%,子葉達到41.79%。
2.5不同生境水曲柳種子的生理特性差異
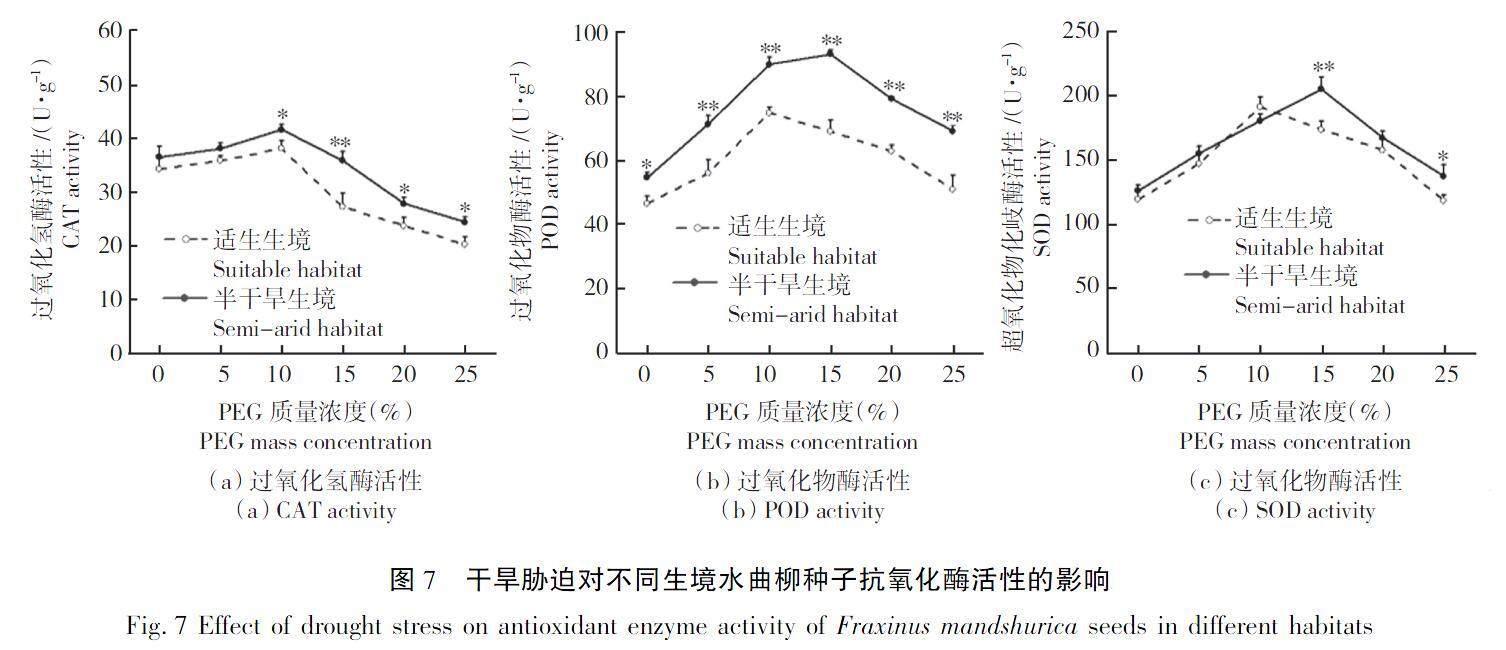
隨著干旱脅迫程度的增強,不同生境水曲柳種子的CAT、POD和SOD活性均先上升后下降,如圖7所示。當PEG質量濃度為10%時,半干旱生境水曲柳種子的CAT活性開始顯著高于適生生境種子,最大差異達到29.89%;半干旱生境水曲柳種子的POD活性始終顯著高于適生生境種子,最大差異達到35.63%;當PEG質量濃度為15%和25%時,半干旱生境水曲柳種子的SOD活性顯著高于適生生境種子,分別高出了17.98%和15.81%。
2.6不同生境水曲柳種子各指標間主成分分析
將兩生境水曲柳種子的形態結構性狀和吸水特性進行主成分分析發現,適生生境和半干旱生境種子特征值大于1的有4個主成分,第1主成分分別為26.4%和24.1%,其他主成分占比均較少,如圖8所示。適生生境種子第1主成分對應的特征向量中翅寬、千粒質量和種皮厚度最大,半干旱生境種子第1主成分對應的特征向量中翅厚、千粒質量和細胞表面積最大,可知適生生境中種翅寬、千粒質量和種皮厚度是影響種子吸水的主要指標,半干旱生境中種翅厚、千粒質量和細胞表面積是影響種子吸水的主要指標。
將兩生境的水曲柳種子萌發和生理參數進行主成分分析發現,適生生境和半干旱生境種子特征值大于1的均有2個主成分,累積貢獻率分別為97.09%和97.74%,其中適生生境種子第1主成分對應的特征向量中發芽指數和活力指數最大,是影響適生生境種子的主要萌發指標;而半干旱生境種子第1主成分對應的特征向量中發芽勢、活力指數和平均發芽時間最長,是影響半干旱生境種子的主要萌發指標。兩生境種子第2主成分對應的特征向量中均是過氧化氫酶和過氧化物歧化酶最大,因此過氧化氫酶和過氧化物歧化酶是影響兩生境種子的主要生理指標,如圖9所示。
3討論
3.1不同生境水曲柳種子的形態結構差異
同一物種內種子的質量是基本恒定的,但是同一物種的不同地理種群的種子品質有一定差別,這種差別與種群的生境條件有關[19]。孫穎等[20]認為,環境條件的變化是種子大小變異的主要選擇壓力。具有較大質量的種子其子葉或胚乳中具有更多供種子萌發的能量,在幼苗生物量積累上具有更大優勢[21]。此外,較大的種子可以增加發芽率,因為大種子產生較長的胚根,可以到達更深的水資源以供緩解供水短缺[22]。在調查的水曲柳種子中,半干旱生境水曲柳種子的種翅長和種翅寬等均顯著大于適生生境種子,且種子千粒質量也較大,這是因為種子的形態與翅果形態之間有協同變化,即種翅面積較大的物種其種子重量也較高[23],這種協同變化是植物對環境長期適應的結果[24]。半干旱生境中產生的大種子在積累代謝調節物質方面比小種子具有更大的優勢,為后期促進幼苗生長提供了充足的營養,因此大種子在逆境中表現出更強的抵抗力和適應性。
水分是種子萌發的先決條件,是一切生命活動的必須因素之一[25]。種皮厚度[26]、種皮完整性和種皮表面細胞排列方式[27]等都會影響種皮的透性,進而對種子吸水特性產生影響。種皮表面細胞是阻擋水分進入種子的第一道屏障[28],種子表皮細胞致密或覆有較厚的蠟質層都會成為影響種子吸水的重要原因[29]。Pascualides等[30]在研究菽麻(Crotalaria juncea)種子吸水特性時發現,雜色小種子的種皮越厚,其吸水速率越慢。本研究中,半干旱生境水曲柳種子的表皮細胞比適生生境種子排列松散,且種子表皮細胞更小,水分更易透過,因此半干旱生境種子吸水更快,但萌發時的吸水率卻顯著低于適生生境種子,這可能是因為半干旱生境種子長期處在干旱環境下,降雨不頻繁且蒸發量大,使得土壤表層水分消耗,土壤干旱程度惡化從而降低了種子對水分的需求。于露等[31]的研究也發現干旱生境下的植被種子(如檸條錦雞兒Caragana korshinskii和沙蘆草Agorpyron mongolicum)更新時所需水分更少,更易成活。另一方面,半干旱生境種子由于環境的影響形成了較厚的種皮,表明半干旱生境水曲柳種子對干旱環境產生了特殊的適應機制,即通過增厚種皮來抵抗逆境環境,厚種皮既可以有效的隔絕種胚與惡劣環境的接觸,更好地保護種胚正常生長發育,從而緩解干旱對種子內部結構的危害,又可以使種子在降雨不穩定的條件下具有更好的保水性能,使種子具有長期保存生活的能力,以適應干旱少雨的自然環境。
3.2不同生境水曲柳種子的抗旱性差異
環境的變化不僅影響種子的形態結構,而且可以直接改變種子的萌發和幼苗的生長[32]。種子萌發期是分辨植物抗旱性強弱的重要時期。研究發現長期生長在逆境環境下的采種母樹其種子表現出更高的發芽率[7]。本研究發現,半干旱生境水曲柳種子的發芽勢和發芽指數在種子抗旱中影響最大,且從PEG質量濃度為10%開始,半干旱生境水曲柳種子的發芽勢和發芽指數均顯著高于適生生境種子,表明半干旱生境下生長的水曲柳長期處于干旱的環境下,產生了特有的防御響應,且這種機制可能形成了穩定的“母體效應”[33]并通過種子不斷的傳遞下去。另外,研究還發現半干旱生境水曲柳種子在干旱脅迫下比適生生境種子萌發時間短且整齊,這可能是與此研究區降雨集中且雨季較短,地面蒸發迅速有關[12],種子只能充分利用短暫的降水條件和周圍環境快速萌發,增加優先形成幼苗的機會,以確保種群在不斷變化和壓力環境中的連續性[34],從而快速完成自己的生活周期繁衍后代。
生活在不同環境中的植物制定出特定的策略來維持遺傳多樣性,以確保其在這種不斷變化的環境中生存[35]。由于生境的不同,種子也會在生理生化方面發生變化,進而影響種子萌發和幼苗生長。徐振朋等[36]通過干旱脅迫對羅布麻(Apocynum venetum)種子萌發及生理特性研究,發現當PEG質量濃度大于10%時,種子的POD和SOD酶活性迅速降低,表明兩種酶在高質量濃度的脅迫下清除活性氧的能力下降。本研究中,兩生境水曲柳種子的POD和SOD是影響種子抗旱的主要生理指標,適生生境種子的POD和SOD在PEG質量濃度為10%時達到高峰,而半干旱生境水曲柳種子的POD和SOD在PEG質量濃度為15%時達到高峰,且POD活性始終顯著高于適生生境種子,表明適生生境種子對干旱脅迫下體內自由基積累傷害的反應更為敏感,而半干旱生境水曲柳種子在長期干旱環境條件適應中已經形成了特殊的耐旱機理,通過協調生理參數,提高了種子忍耐干旱的有效性[37]。這反映了半干旱生境水曲柳種子在生理方面的優勢和耐受性,也從另一方面印證了半干旱生境種子對逆境環境的抗旱適應性。
4結論
綜上可知,半干旱生境的水曲柳種子在形態、結構以及生理上均產生了一定的適應性變化,對干旱脅迫具有更強的耐受力。半干旱生境種子品質大、種皮厚,吸水保水性能好,萌發時需水量低,其種子發芽能力在PEG質量濃度為10%開始顯著高于適生生境種子,更具耐旱性。因此,半干旱地區生產的水曲柳種子對干旱環境具有更強的適應性,在干旱地區選種造林時可優先選用半干旱生境水曲柳種子,擴大種質資源范圍。
【參考文獻】
[1]GONG Z Z, XIONG L M, SHI H Z, et al. Plant abiotic stress response and nutrient use efficiency[J]. Science China Life Sciences, 2020, 63(5): 635-674.
[2]岳莉然,程貝貝.鹽堿脅迫及干旱脅迫對大花剪秋蘿種子萌發的影響[J].北方園藝,2019(10):86-93.
YUE L R, CHENG B B. Effects of saline-alkali stress and drought stress on seed germination of Lychnis fulgens Fisch[J]. Northern Horticulture, 2019(10): 86-93.
[3]KAPOOR D, BHARDWAJ S, LANDI M, et al. The impact of drought in plant metabolism: how to exploit tolerance mechanisms to increase crop production[J]. Applied Sciences, 2020, 10(16): 5692.
[4]劉長成,劉玉國,郭柯.四種不同生活型植物幼苗對喀斯特生境干旱的生理生態適應性[J].植物生態學報,2011,35(10):1070-1082.
LIU C C, LIU Y G, GUO K. Ecophysiological adaptations to drought stress of seedlings of four plant species with different growth forms in Karst habitats[J]. Chinese Journal of Plant Ecology, 2011, 35(10): 1070-1082.
[5]MAO P L, ZANG R Z, SHAO H B, et al. The ecological adaptability of four typical plants during the early successional stage of a tropical rainforest[J]. Plant Biosystems -An International Journal Dealing with All Aspects of Plant Biology, 2014, 148(2): 288-296.
[6]閆小紅,曾建軍,周兵,等.干旱脅迫對白花鬼針草異型瘦果萌發與幼苗生長的影響[J].生態學雜志,2019,38(11):3327-3334.
YAN X H, ZENG J J, ZHOU B, et al. Effects of drought stress on germination and seedling growth of heteromorphic achenes of Bidens alba[J]. Chinese Journal of Ecology, 2019, 38(11): 3327-3334.
[7]MIRA S, ARNAL A, PEREZ-GARCIA F. Habitat-correlated seed germination and morphology in populations of Phillyrea angustifolia L. (Oleaceae)[J]. Seed Science Research, 2017, 27(1): 50-60.
[8]張源一,李愛,嚴卓立,等.5種薄荷對PEG模擬干旱脅迫的生理響應與耐旱性評價[J].天津農學院學報,2022,29(1):1-7.
ZHANG Y Y, LI A, YAN Z L, et al. Physiological response and drought tolerance evaluation of five mints to PEG-simulated drought stress[J]. Journal of Tianjin Agricultural University, 2022, 29(1): 1-7.
[9]閆月,盧艷,崔程程,等.聚乙二醇引發處理對水曲柳種子在高溫下萌發的影響[J].東北林業大學學報,2020,48(6):13-16.
YAN Y, LU Y, CUI C C, et al. Effect of PEG priming treatment on the germination of Fraxinus mandshurica seeds at high temperature[J]. Journal of Northeast Forestry University, 2020, 48(6): 13-16.
[10]王繼剛.水曲柳最佳種源選擇與種源區劃[D].哈爾濱:東北林業大學,2001.
WANG J G. Study on theselection of optimal provenance and provenance[D]. Harbin: Northeast Forestry University, 2001.
[11]馬建路,石家琛,景鳳鳴.水曲柳立地區劃[J].東北林業大學學報,1991,19(S1):62-68.
MA J L, SHI J C, JING F M. Site classification for manchurian ash[J]. Journal of Northeast Forestry University, 1991, 19(S1): 62-68.
[12]馬常欽.水曲柳雌雄株復葉功能性狀對不同生境的響應[D].哈爾濱:東北林業大學,2022.
MA C Q. Responses of functional characters of compound leaves of male and female Fraxinus mandshurica to different habitats[D]. Harbin: Northeast Forestry University, 2022.
[13]宿昊,申耀榮,蔡靖,等.不同種源紅樺種子的萌發特性[J].西北林學院學報,2021,36(3):109-114.
SU H, SHEN Y R, CAI J, et al. Germination characteristics of Betula albosinensis seeds from different provenances[J]. Journal of Northwest Forestry University, 2021, 36(3): 109-114.
[14]郭學民,趙曉曼,徐珂,等.蓖麻種子結構的解剖和顯微觀察[J].作物學報,2020,46(6):914-923.
GUO X M, ZHAO X M, XU K, et al. Anatomy and microscopic observation of Ricinus communis seed structure[J]. Acta Agronomica Sinica, 2020, 46(6): 914-923.
[15]劉帥,陳良珂,房克鳳,等.板栗種子淀粉體發育的掃描電鏡觀察[J].電子顯微學報,2015,34(4):346-350.
LIU S, CHEN L K, FANG K F, et al. Observation of amyloplast development in chestnut seed by scanning electron microscope[J]. Journal of Chinese Electron Microscopy Society, 2015, 34(4): 346-350.
[16]艾沙江·阿不都沙拉木,迪麗娜爾·阿布拉,張凱,等.喀什霸王的結實和種子萌發特性[J].植物生態學報,2019,43(5):437-446.
AYSAJAN A B D S L M, DILINAER A B L, ZHANG K, et al. Fruit set and seed germination traits of Zygophyllum kaschgaricum[J]. Chinese Journal of Plant Ecology, 2019, 43(5): 437-446.
[17]王營,王麗昕,張彥妮.鹽堿及PEG6000脅迫對細香蔥種子萌發的影響[J].東北林業大學學報,2019,47(7):25-30.
WANG Y, WANG L X, ZHANG Y N. Effects of salt-alkali and PEG 6000 stress on seed germination of Allium schoenoprasum[J]. Journal of Northeast Forestry University, 2019, 47(7): 25-30.
[18]李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.
LI H S. Principles and techniques of plant physiological biochemical experiment[M]. Beijing: Higher Education Press, 2000.
[19]ARELLANO G, PECO B. Testing the role of seed size in annual legume seedling performance under experimental autumn moisture conditions[J]. Journal of Vegetation Science, 2012, 23(4): 690-697.
[20]孫穎,王阿香,楊雪,等.側金盞花種子發育過程中生理生化動態變化特性[J].東北林業大學學報,2015,43(7):35-37,135.
SUN Y, WANG A X, YANG X, et al. Physiological and biochemical characteristics of Adonis amurensis during seed development[J]. Journal of Northeast Forestry University, 2015, 43(7): 35-37, 135.
[21]TANVEER A, TASNEEM M, KHALIQ A, et al. Influence of seed size and ecological factors on the germination and emergence of field bindweed (Convolvulus arvensis)[J]. Planta Daninha, 2013, 31(1): 39-51.
[22]ZHAO L L, MO B T, WANG P C, et al. Relationship of Sophora davidii seed size to germination, dormancy, and mortality under water stress[J]. South African Journal of Botany, 2015, 99: 12-16.
[23]張金峰,葛樹森,李玉堂,等.長白山9種槭樹的翅果擴散及種子萌發研究[J].生態學報,2022,42(4):1441-1449.
ZHANG J F, GE S S, LI Y T, et al. Dispersal and germination of nine maple (Acer, spp) trees in Changbai Mountain Area[J]. Acta Ecologica Sinica, 2022, 42(4): 1441-1449.
[24]馮秋紅,李登峰,于濤,等.極小種群野生植物梓葉槭的種實表型變異特征[J].生物多樣性,2020,28(3):314-322.
FENG Q H, LI D F, YU T, et al. Phenotypic fruit and seed variations of Acer catalpifolium, a wild plant with extremely small populations in China[J]. Biodiversity Science, 2020, 28(3): 314-322.
[25]李茜,任運濤,牛得草,等.幾種旱生灌木種子萌發特性及化學成分[J].中國沙漠,2015,35(2):345-351.
LI Q, REN Y T, NIU D C, et al. Seed germination characteristics and chemical composition of eight xerophytic shrub species[J]. Journal of Desert Research, 2015, 35(2): 345-351.
[26]WU Y, SHEN Y B. Seed coat structural and permeability properties of Tilia miqueliana seeds[J]. Journal of Plant Growth Regulation, 2021, 40(3): 1198-1209.
[27]張琪,錢滕,王歡,等.加拿大紫荊種子硬實性解除及其吸水特性研究[J].南京林業大學學報(自然科學版),2021,45(3):137-142.
ZHANG Q, QIAN T, WANG H, et al. Hardness breaking and mechanisms of water absorption in Cercis canadensis seeds[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2021, 45(3): 137-142.
[28]文婷婷,王洋,利站,等.豌豆種皮結構和成分對種子透水性的影響[J].種子,2016,35(1):19-25.
WEN T T, WANG Y, LI Z, et al. Effects of seed coat structure and composition on water permeability of pea (Pisum sativum L.) seed[J]. Seed, 2016, 35(1): 19-25.
[29]陳麗,代松,馬青江,等.合歡種皮結構及其與吸水的關系[J].林業科學,2019,55(5):46-54.
CHEN L, DAI S, MA Q J, et al. Structure of seed coat of Albizia julibrissin and its relationship with water uptake[J]. Scientia Silvae Sinicae, 2019, 55(5): 46-54.
[30]PASCUALIDES A L, PLANCHUELO A M. Seed morphology and imbibition pattern of Crotalaria juncea L. (Fabaceae)[J]. Seed Science and Technology, 2007, 35(3): 760-764.
[31]于露,郭天斗,孫忠超,等.荒漠草原向灌叢地轉變過程中兩種優勢植物種子萌發及閾值特征[J].生態學報,2021,41(10):4160-4169.
YU L, GUO T D, SUN Z C, et al. The seed germination characteristics and thresholds of two dominant plants in desert grassland-shrubland transition of the eastern Ningxia, China[J]. Acta Ecologica Sinica, 2021, 41(10): 4160-4169.
[32]LI Z Q, LU W, YANG L, et al. Seed weight and germination behavior of the submerged plant Potamogeton pectinatus in the arid zone of Northwest China[J]. Ecology and Evolution, 2015, 5(7): 1504-1512.
[33]CALLEJAS-DIAZ M, CHAMBEL M R, SAN-MARTIN-LOREN J, et al. The role of maternal age, growth, and environment in shaping offspring performance in an aerial conifer seed bank[J]. American Journal of Botany, 2022, 109(3): 366-376.
[34]GUL B, ANSARI R, FLOWERS T J, et al. Germination strategies of halophyte seeds under salinity[J]. Environmental and Experimental Botany, 2013, 92: 4-18.
[35]SORIANO P, ESTRELLES E, MARTINEZ-NIETO M I, et al. Environmental predictors of seed germination in two Halocnemum species from Mediterranean (Balearic, Tyrrenic and Adriatic) and Red Sea coastal salt marshes[J]. Seed Science Research, 2022, 32(4): 246-263.
[36]徐振朋,宛濤,蔡萍,等.PEG模擬干旱脅迫對羅布麻種子萌發及生理特性的影響[J].中國草地學報,2015,37(5):75-80.
XU Z P, WAN T, CAI P, et al. Effects of PEG simulated drought stress on germination and physiological properties of Apocynum venetum seeds[J]. Chinese Journal of Grassland, 2015, 37(5): 75-80.
[37]閆興富,鄧曉娟,王靜,等.種子大小和干旱脅迫對遼東櫟幼苗生長和生理特性的影響[J].應用生態學報,2020,31(10):3331-3339.
YAN X F, DENG X J, WANG J, et al. Effects of seed size and drought stress on the growth and physiological characteristics of Quercus wutaishanica seedlings[J]. Chinese Journal of Applied Ecology, 2020, 31(10): 3331-3339.
