不同F(xiàn)ecB基因型和不同直徑綿羊卵泡中BMP/SMAD通路活性及蛋白表達(dá)差異
2024-10-14 00:00:00龔一鳴賈一軒李佳駿王翔宇賀小云儲(chǔ)明星狄冉
畜牧獸醫(yī)學(xué)報(bào) 2024年9期






摘 要: 旨在探究不同F(xiàn)ecB基因型對(duì)綿羊卵泡中BMP/SMAD通路活性和蛋白表達(dá)的影響;揭示成熟大卵泡和小卵泡之間BMP/SMAD通路活性和蛋白表達(dá)的差異。本研究采用TaqMan分型方法篩選出不同F(xiàn)ecB基因型的母羊,同期發(fā)情后取卵泡期成熟卵泡和黃體期卵巢表面小卵泡,利用免疫印跡法(Western blot)測(cè)定BMP/SMAD通路相關(guān)蛋白表達(dá)水平和通路活性。結(jié)果表明,對(duì)于小卵泡組,F(xiàn)ecB突變型卵泡中骨形態(tài)發(fā)生蛋白1B型受體(bone morphogenetic protein receptor type 1B,BMPR1B)表達(dá)量顯著高于野生型卵泡(Plt;0.05),但SMAD家族成員4(SMAD family member 4,SMAD4)表達(dá)量和SMAD1/5/9的磷酸化水平顯著低于野生型卵泡(Plt;0.05);對(duì)于成熟大卵泡組,F(xiàn)ecB突變型卵泡中FKBP脯氨酰異構(gòu)酶1A(FKBP prolyl isomerase 1A,F(xiàn)KBP1A)和SMAD4表達(dá)量顯著低于野生型卵泡(Plt;0.05),Ⅰ型受體(BMPR1B)和Ⅱ型受體 (BMPR2) 的蛋白表達(dá)量及SMAD1/5/9的磷酸化水平在兩種基因型之間未顯示出顯著差異。另一方面,對(duì)比FecB突變型小卵泡和成熟大卵泡發(fā)現(xiàn):成熟大卵泡中BMPR1B和SMAD4蛋白表達(dá)量和SMAD1/5/9磷酸化程度顯著高于小卵泡 (Plt;0.05)。上述結(jié)果表明,由于SMAD4表達(dá)量的下降,F(xiàn)ecB突變型大、小卵泡中結(jié)合到基因組靶區(qū)域的SMAD4-SMAD1/5/9蛋白復(fù)合物均相對(duì)較少,將導(dǎo)致通路活性降低,而且由于小卵泡中較低的SMAD1/5/9磷酸化水平,其通路活性更低。另外,綿羊突變型卵泡生長(zhǎng)發(fā)育成熟后BMP/SMAD通路活性顯著增強(qiáng)。
關(guān)鍵詞: 綿羊;FecB突變;卵泡;BMP/SMAD通路;SMAD4
中圖分類(lèi)號(hào): S826.3
文獻(xiàn)標(biāo)志碼:A 文章編號(hào): 0366-6964(2024)09-3957-11
BMP/SMAD Pathway Activity and Protein Expression Profiles in Ovarian Follicles
with Different Diameters in Diverse FecB Genotyped Ewes
GONG" Yiming, JIA" Yixuan, LI" Jiajun, WANG" Xiangyu, HE" Xiaoyun,
CHU" Mingxing*, DI" Ran*
(State Key Laboratory of Animal Biotech Breeding, Institute of Animal Science,
Chinese Academy of Agricultural Sciences, Beijing 100193," China)
Abstract:" This study aimed to investigate the effects of different FecB genotypes on the BMP/SMAD signaling pathway activity and protein expression in sheep follicles, and explored the differences in BMP/SMAD signaling pathway activity and protein expression between mature large follicles and small follicles. Firstly, TaqMan method was used to screen ewes with different genotypes of FecB. Secondly, after estrus synchronization, mature follicles in follicular phase and small follicles in luteal phase were sampled, and the pathway activity and the protein expression of BMP/SMAD pathway were determined by Western blot. The result showed that, for the small follicle group, the expression of bone morphogenetic protein receptor type 1B (BMPR1B) in FecB mutant follicles was significantly higher than that in wild-type follicles (Plt;0.05), but the expression of SMAD family member 4 (SMAD4) and the phosphorylation levels of SMAD1/5/9 were significantly lower than those in wild-type follicles (Plt;0.05). For the mature large follicle group, the expression of FKBP prolyl isomerase 1A (FKBP1A) and SMAD4 in FecB mutant follicles was significantly lower than that in wild-type follicles (Plt;0.05), and there was no significant difference in the protein expression of type I receptor (BMPR1B) and type II receptor (BMPR2) and the phosphorylation levels of SMAD1/5/9 between the two genotypes. On the other hand, when comparing FecB mutant small follicles with mature large follicles, the protein expression levels of BMPR1B and SMAD4 and the phosphorylation level of SMAD1/5/9 in mature large follicles were significantly higher than those in small follicles (Plt;0.05). The results indicated that due to the decrease in the expression of SMAD4, the FecB mutant large and small follicles have relatively fewer SMAD4-SMAD1/5/9 protein complexes bound to the genomic target regions, which means a reduction in pathway activity. Moreover, due to the lower phosphorylation level of SMAD1/5/9 in small follicles, its pathway activity was lower. Additionally, after the development and maturation of the mutant sheep follicles, the activity of the BMP/SMAD pathway was significantly enhanced.
Key words: sheep; FecB mutation; follicle; BMP/SMAD pathway; SMAD4
*Corresponding authors:DI Ran, E-mail: dirangirl@163.com; CHU Mingxing, E-mail:mxchu@263.net
BMPR1B基因作為提高綿羊產(chǎn)羔數(shù)的主效基因,其FecB突變?cè)黾赢a(chǎn)羔數(shù)的具體分子機(jī)制尚未被完全解析。近年來(lái),有學(xué)者推測(cè)FecB突變可能通過(guò)改變BMP/SMAD通路活性進(jìn)而降低下游靶基因轉(zhuǎn)錄水平,最終縮短排卵周期和提高排卵數(shù)[1-2]。通過(guò)蛋白建模分析發(fā)現(xiàn),F(xiàn)ecB突變后的BMPR1B蛋白與抑制蛋白FKBP1A可以形成強(qiáng)氫鍵[3],阻礙FKBP1A從BMPR1B蛋白上正常解離下來(lái),BMP/SMAD通路活性可能受到抑制。BMP/SMAD通路主要由BMP受體(bone morphogenetic protein receptor,BMPR)和SMADs家族蛋白構(gòu)成[4]。該通路中BMP受體和SMADs家族蛋白密切參與卵泡顆粒細(xì)胞的增殖與凋亡過(guò)程[5-6]。BMP配體與Ⅰ型受體(bone morphogenetic protein receptor Ⅰ,BMPR1)和Ⅱ型受體(bone morphogenetic protein receptor Ⅱ,BMPR2)結(jié)合后,形成異源四聚體復(fù)合物進(jìn)行級(jí)聯(lián)磷酸化反應(yīng)[7]。首先BMP抑制蛋白FKBP1A從BMPⅠ型受體(BMPR1B)上解離,使得Ⅰ型受體GS區(qū)域中絲氨酸和蘇氨酸殘基被BMPR2激活,從而具備了Ⅰ型受體激酶活性[8]。激活后的Ⅰ型受體誘導(dǎo)下游SMAD1/5/9蛋白發(fā)生磷酸化反應(yīng),磷酸化的SMAD1/5/9蛋白須要與共同通路型Smad(co-SMAD,SMAD4)互相結(jié)合形成異源四聚體復(fù)合物遷移至細(xì)胞核中調(diào)控下游靶基因的轉(zhuǎn)錄[4,7,9-11]。
目前,有關(guān)綿羊FecB突變對(duì)BMP/SMAD通路活性及蛋白表達(dá)變化的研究匱乏。Fabre等[12]研究發(fā)現(xiàn),在綿羊FecB野生型和突變型卵泡中添加BMP配體后,均顯著激活BMP/SMAD通路,進(jìn)而抑制了顆粒細(xì)胞中孕酮的分泌;在添加相同劑量的BMP配體時(shí),與大卵泡相比,小卵泡抑制孕酮分泌的作用更強(qiáng);與野生型小卵泡相比,F(xiàn)ecB突變型小卵泡抑制孕酮分泌的能力顯著降低。上述結(jié)果表明,F(xiàn)ecB突變通過(guò)BMP/SMAD通路調(diào)控了下游孕酮的分泌。因此,為了進(jìn)一步探究FecB突變對(duì)BMP/SMAD通路的具體影響,本研究通過(guò)比較不同F(xiàn)ecB 基因型卵泡中BMP/SMAD通路活性及相關(guān)蛋白表達(dá)量,探究FecB突變對(duì)綿羊卵泡中該通路的影響,探討綿羊小卵泡和成熟大卵泡中BMP/SMAD通路活性及蛋白表達(dá)的變化,有助于揭示該通路對(duì)綿羊卵泡發(fā)育的調(diào)控作用,為進(jìn)一步揭示FecB突變提高綿羊產(chǎn)羔數(shù)的分子機(jī)制提供理論基礎(chǔ)。另外,也有研究表明豬的卵泡發(fā)育過(guò)程中顆粒細(xì)胞增殖加快,BMP/SMAD通路活性改變[13]。因此,本研究從卵泡發(fā)育的角度探討了綿羊小卵泡和成熟大卵泡中BMP/SMAD通路活性及蛋白表達(dá)的變化,有助于揭示該通路對(duì)綿羊卵泡發(fā)育的調(diào)控作用
1 材料與方法
1.1 綿羊卵泡組織采集
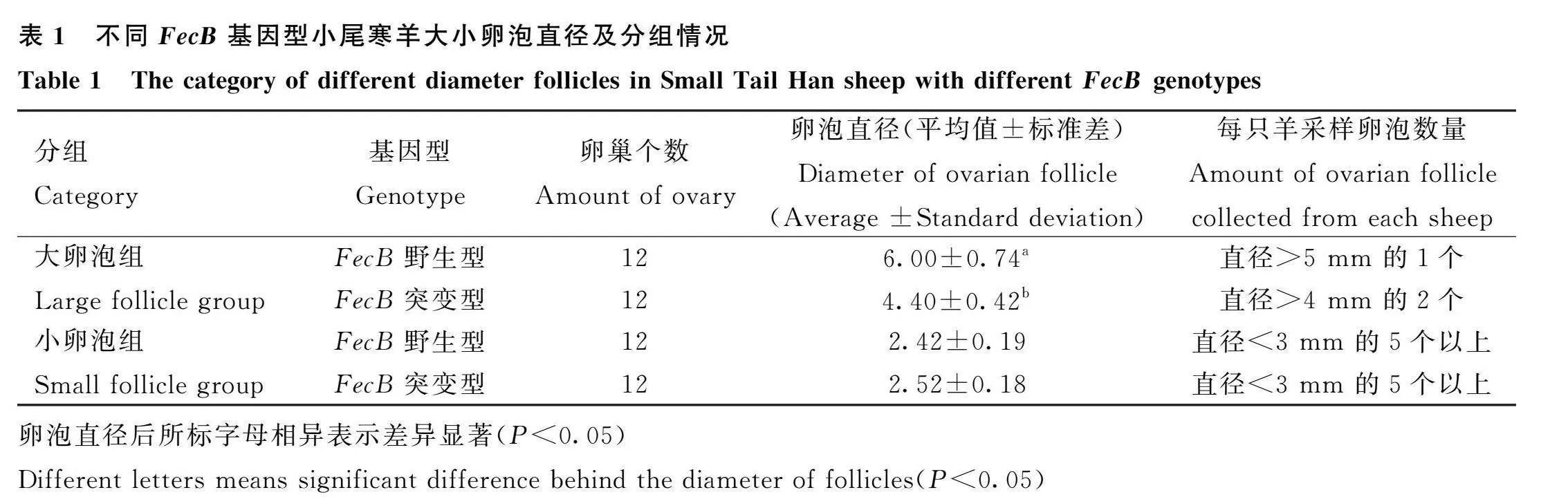
在山東省菏澤市同一個(gè)羊場(chǎng)內(nèi)選擇3歲、健康、體重均在70 kg左右的小尾寒經(jīng)產(chǎn)母羊。采集母羊頸靜脈血以提取基因組DNA,利用自主研發(fā)的TaqMan探針對(duì)所有DNA樣本進(jìn)行快速FecB分型[14]。從中篩選出突變型純合型(BB)和野生型(WW)母羊各24只,采用CIDR栓進(jìn)行同期發(fā)情處理。在卵泡期(撤栓后45 h)對(duì)12只突變型和12只野生型母羊進(jìn)行手術(shù),用游標(biāo)卡尺測(cè)量卵巢上卵泡的直徑,然后迅速抽取成熟大卵泡的卵泡液速凍在液氮中,然后復(fù)位卵巢進(jìn)行腹部皮膚的縫合。在黃體期(撤栓后第10天)對(duì)剩余12只突變型和12只野生型母羊進(jìn)行手術(shù),測(cè)量卵泡直徑,迅速抽取小卵泡的卵泡液速凍在液氮中,然后復(fù)位卵巢進(jìn)行腹部皮膚的縫合。
1.2 卵泡液蛋白提取及WB試驗(yàn)分組
取小尾寒羊冷凍后的卵泡液,快速放入盛有液氮的研缽中,使用研磨工具將其研磨至粉末狀。利用全蛋白提取試劑盒(Solarbio,中國(guó)北京,BC3710)對(duì)組織蛋白進(jìn)行提取,將來(lái)自4個(gè)不同個(gè)體的相同基因型蛋白進(jìn)行混勻,利用BCA蛋白濃度測(cè)定試劑盒對(duì)提取到的蛋白濃度進(jìn)行測(cè)量(Solarbio,中國(guó)北京,PC0020)。按照等蛋白體積(20 ng)制備上樣液。Western blot試驗(yàn)分組如下:FecB不同基因型組中設(shè)置3個(gè)生物學(xué)重復(fù),每個(gè)重復(fù)由4個(gè)卵泡直徑相近的卵泡蛋白進(jìn)行混勻上樣,確保試驗(yàn)真實(shí)準(zhǔn)確。
1.3 免疫印跡法(Western blot)檢測(cè)BMP/SMAD通路中蛋白表達(dá)和磷酸化水平
本試驗(yàn)采用碧云天10%預(yù)制膠(BeyoGelTM,中國(guó)上海,P0455M)進(jìn)行電泳,濃縮膠電壓調(diào)整為 90 V,30 min,待蛋白被壓縮為一條線(xiàn)時(shí),更改電壓,120 V,45 min,利用活化液活化PVDF膜,160 mA,120 min條件下進(jìn)行轉(zhuǎn)膜,采用碧云天快速封閉液(BeyoGelTM,中國(guó)上海,P0252)室溫封閉2 h。一抗采用Abcam公司提供的抗體(Anti-BMPR1B,ab175385、Anti-FKBP1A,ab58072、Anti-BMPR2,ab130206、重組Anti-Smad4,ab40759和重組Anti-SMAD1/5/9,ab300164)和Cell Signaling TechnologyTM公司提供的磷酸化SMAD1/5/9抗體[15](Phospho-SMAD1(Ser463/465)/SMAD5 (Ser463/465)/SMAD9(Ser465/467)(D5B10)Rabbit mAb,13820S),內(nèi)參抗體采用GAPDH抗體(Proteintech,中國(guó)武漢,204941-1-AP),二抗采用Proteintech公司提供的抗體(HRP-conjugated Affinipure Goat Anti-Rabbit IgG(H+L),SA00001-2 和HRP-conjugated Affinipure Goat Anti-Mouse IgG(H+L),SA00001-1))。
1.4 數(shù)據(jù)處理
試驗(yàn)結(jié)果利用ImageJ[16]進(jìn)行灰度值統(tǒng)計(jì),遵循目標(biāo)蛋白灰度值/內(nèi)參灰度值計(jì)算蛋白相對(duì)表達(dá)量和相對(duì)磷酸化水平。使用SPSS單因素方差分析進(jìn)行P值計(jì)算,通過(guò)GraphPad(GraphPad Software,www.graphpad.com)進(jìn)行繪圖。
2 結(jié) 果
2.1 不同F(xiàn)ecB基因型小尾寒羊卵泡直徑大小分析
本研究利用電子游標(biāo)卡尺對(duì)FecB不同基因型綿羊卵泡直徑測(cè)量發(fā)現(xiàn):FecB突變型成熟大卵泡直徑顯著低于FecB野生型成熟大卵泡(Plt;0.05)。具體測(cè)定及分析結(jié)果如表1所示。
2.2 不同F(xiàn)ecB基因型小尾寒羊卵泡中BMP/SMAD通路蛋白表達(dá)及磷酸化水平分析
本研究將卵泡細(xì)分為小卵泡和成熟大卵泡組,分別探討了大、小卵泡中FecB突變對(duì)BMP/SMAD通路蛋白表達(dá)量及通路活性的影響。
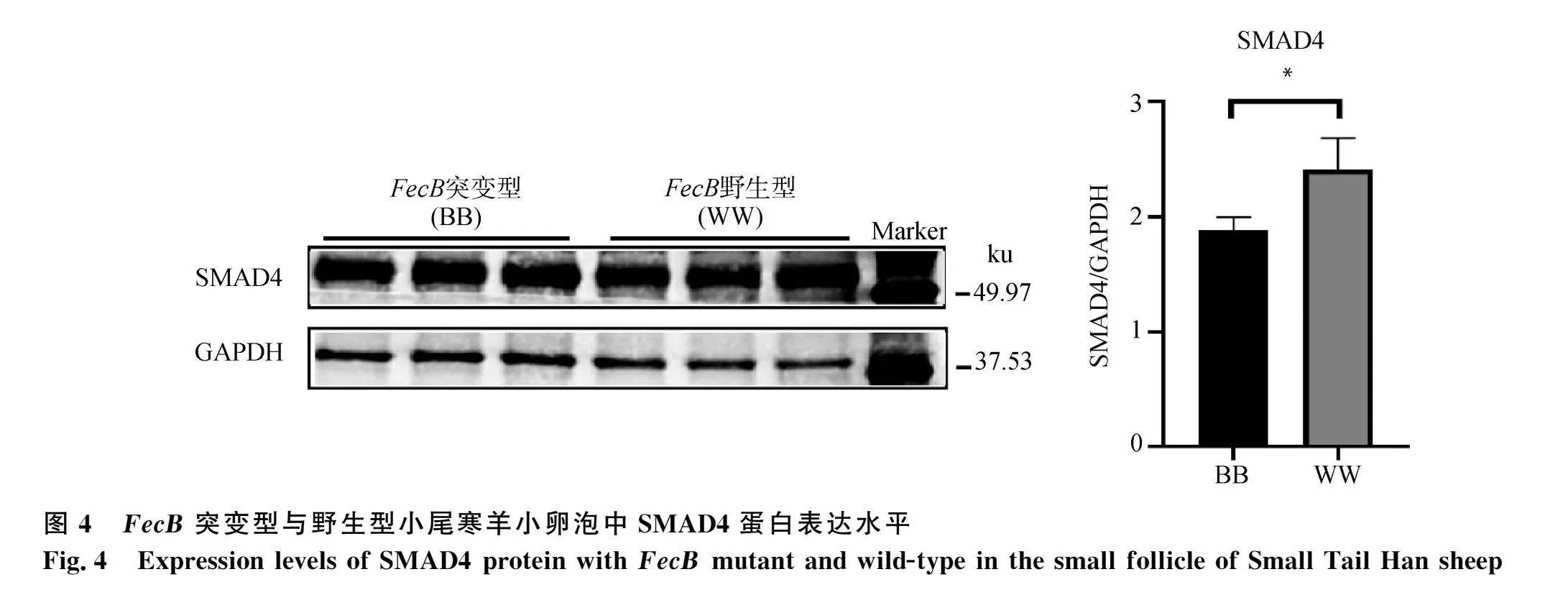
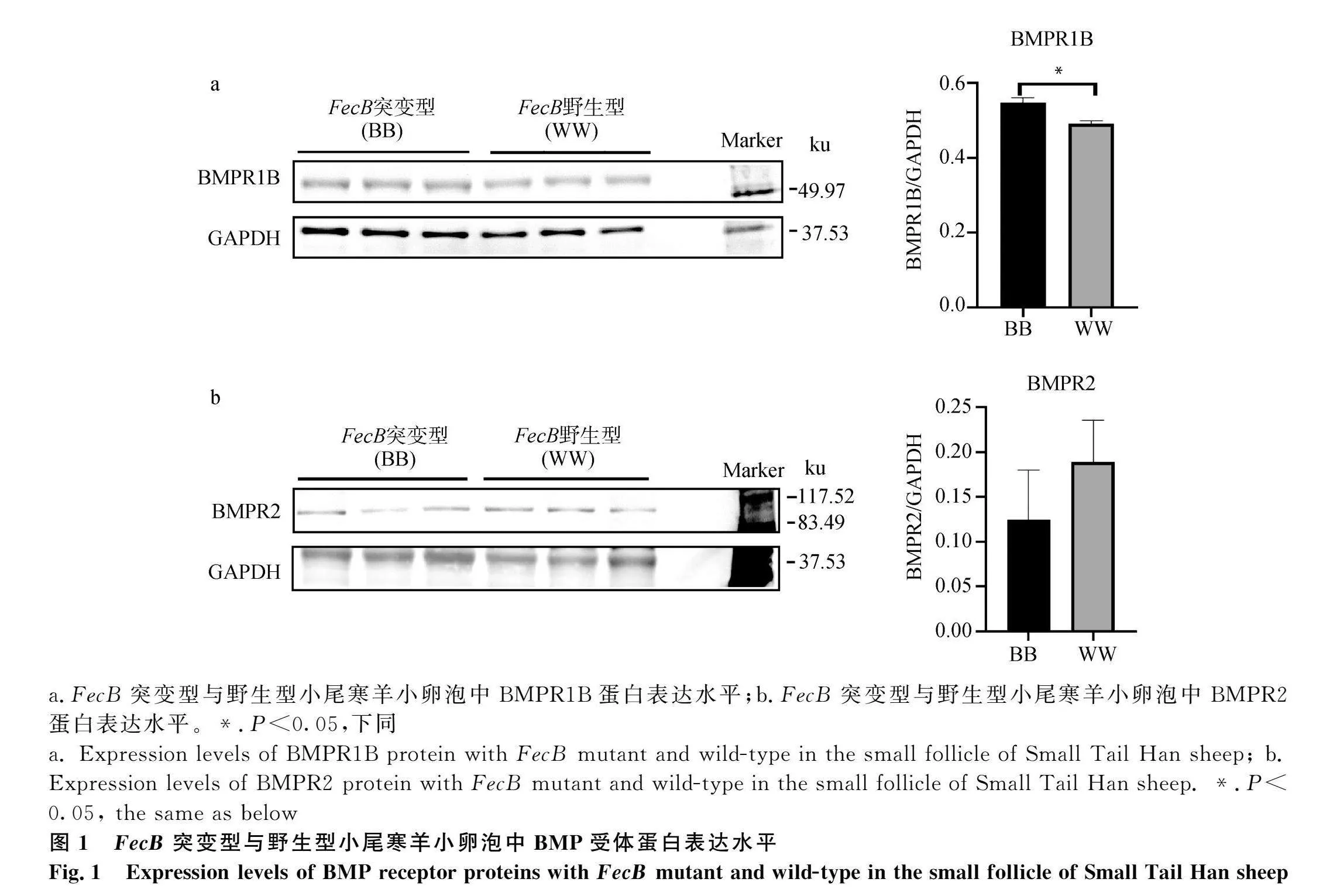
針對(duì)小卵泡組分析,發(fā)現(xiàn)FecB突變型小尾寒羊卵泡組織BMP/SMAD通路中Ⅰ型受體蛋白BMPR1B的表達(dá)量顯著高于野生型小尾寒羊(Plt;0.05,圖1a),但是Ⅱ型受體BMPR2的表達(dá)水平在突變型和野生型小尾寒羊卵泡組織之間無(wú)顯著差異(Pgt;0.05,圖1b)。突變型和野生型小尾寒羊卵泡組織中抑制蛋白FKBP1A的表達(dá)水平無(wú)顯著差異(Pgt;0.05,圖2)。FecB突變型小尾寒羊卵泡組織中SMAD1/5/9的磷酸化水平顯著低于野生型小尾" 寒羊(Plt;0.05,圖3),同時(shí)轉(zhuǎn)錄因子SMAD4的蛋白表達(dá)水平也顯著低于野生型小尾寒羊(Plt;0.05,圖4)。因此,與野生型小尾寒羊相比,F(xiàn)ecB突變型小尾寒羊卵泡組織中結(jié)合到基因組靶區(qū)域的SMAD4-SMAD1/5/9蛋白復(fù)合物相對(duì)較少,加之較低的磷酸化水平,都將使通路活性顯著降低。
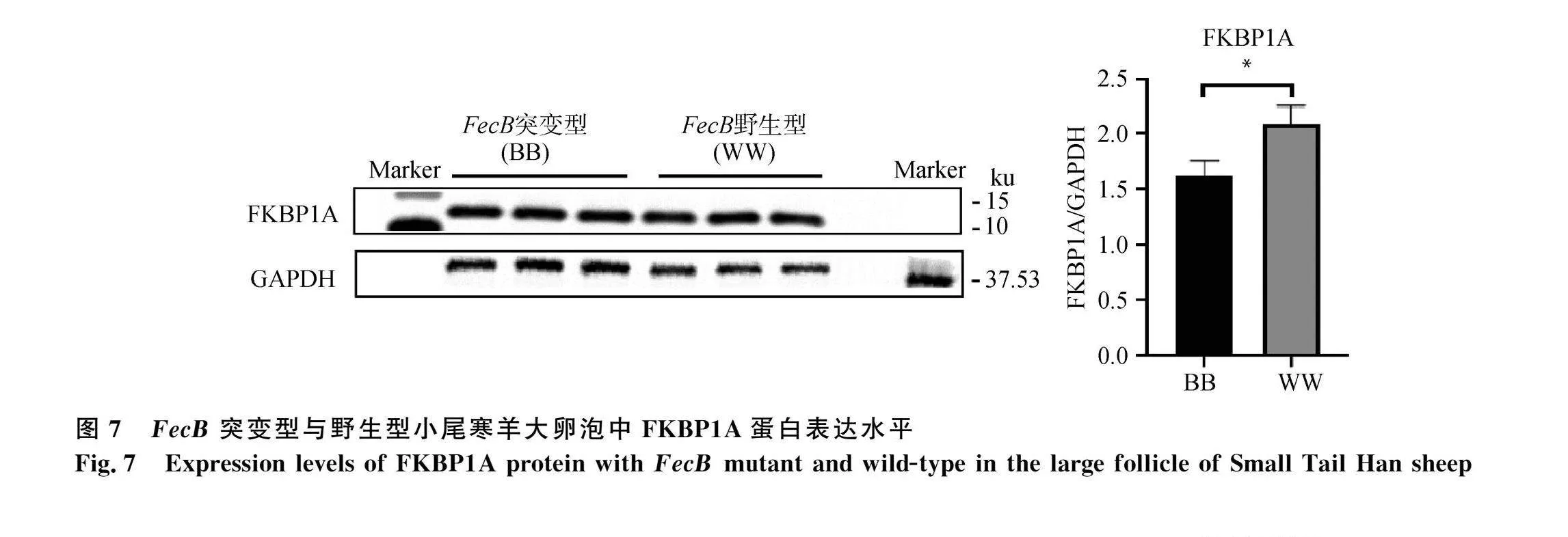
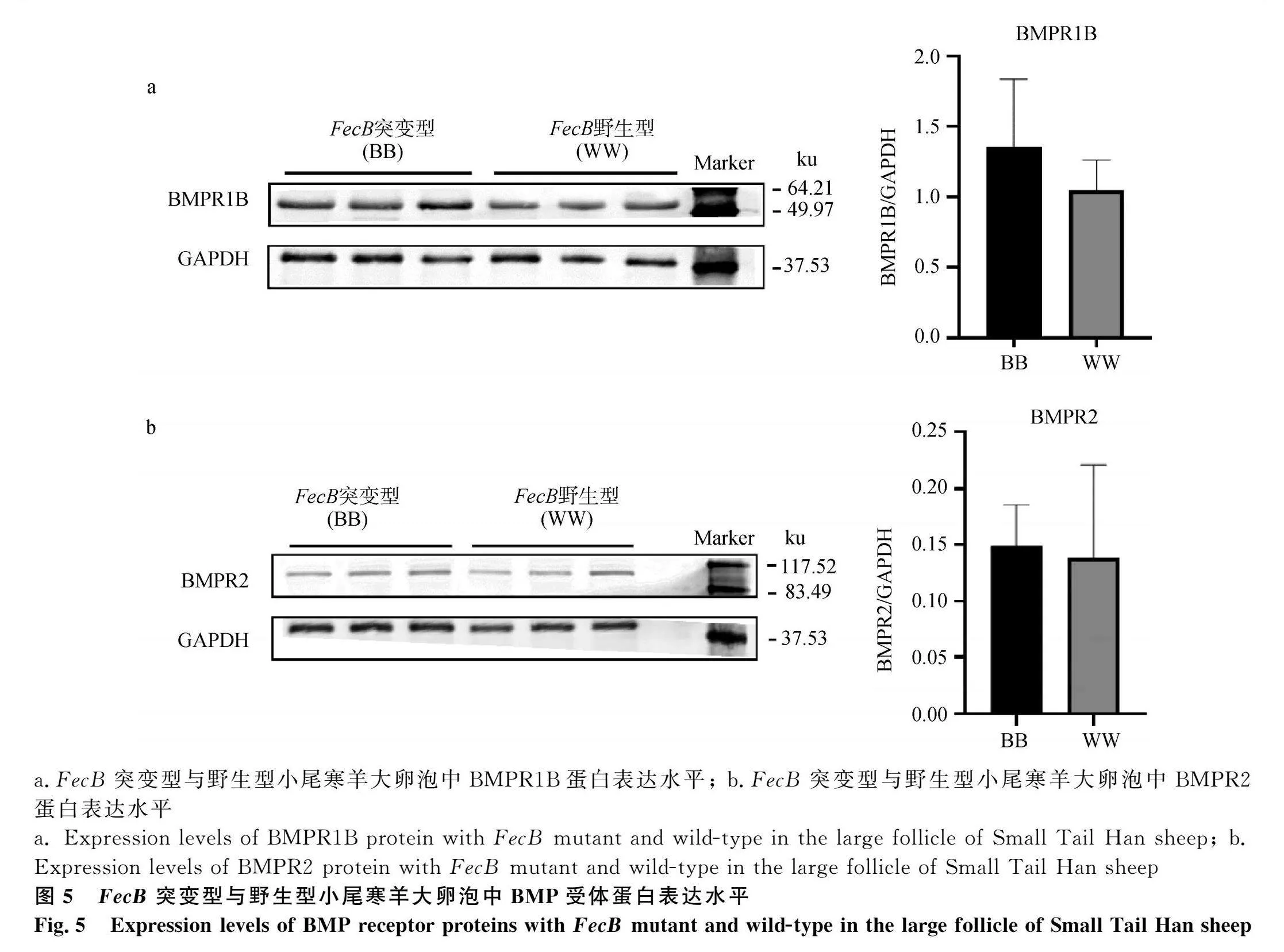
同時(shí),本研究對(duì)比了FecB突變型和野生型小尾寒羊大卵泡中BMP/SMAD通路蛋白表達(dá)水平及通路活性。BMP/SMAD通路中兩個(gè)受體蛋白BMPR1B(圖5a)和BMPR2(圖5b)的表達(dá)在FecB突變型和野生型小尾寒羊卵泡之間均無(wú)顯著差異(Pgt;0.05)。FecB突變型小尾寒羊大卵泡中轉(zhuǎn)錄因子SMAD4的蛋白表達(dá)量顯著低于野生型卵泡組織(Plt;0.05,圖6)。抑制蛋白FKBP1A在FecB突變型大卵泡中表達(dá)量顯著低于野生型大卵泡(Plt;0.05,圖7)。BMPR1B下游轉(zhuǎn)錄因子SMAD1/5/9的磷酸化水平在兩種基因型小尾寒羊卵泡之間無(wú)顯著差異(圖8)。綜上,由于FecB突變型小尾寒羊大卵泡中轉(zhuǎn)錄因子SMAD4的蛋白表達(dá)量顯著低于野生型卵泡組織,這將導(dǎo)致FecB突變型小尾寒羊大卵泡中結(jié)合到基因組靶區(qū)域的SMAD4-SMAD1/5/9蛋白復(fù)合物相對(duì)較少,啟動(dòng)下游靶基因轉(zhuǎn)錄的能力將隨之降低,即突變型小尾寒羊大卵泡中BMP/SMAD通路活性降低。
2.3 小尾寒羊卵巢小卵泡和大卵泡中BMP/SMAD通路活性及蛋白表達(dá)差異
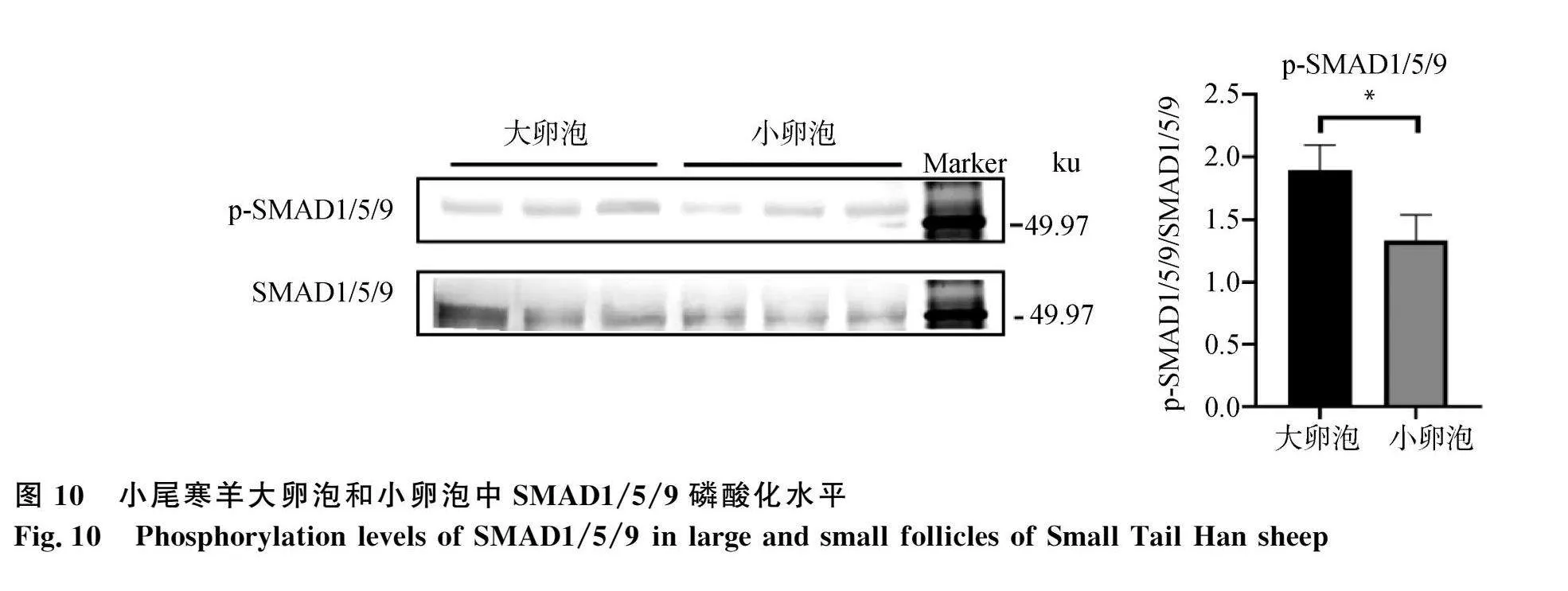
從卵泡生長(zhǎng)發(fā)育的角度,本研究分析了母羊卵" 巢中成熟大卵泡和小卵泡組織BMP/SMAD通路活性及BMPR1B和SMAD4的蛋白表達(dá)差異。Western blot結(jié)果顯示,成熟大卵泡中BMPR1B(圖9)蛋白表達(dá)水平和SMAD1/5/9的磷酸化水平(圖10)顯著高于小卵泡(Plt;0.05),而且大卵泡中SMAD4蛋白表達(dá)水平極顯著高于小卵泡(Plt;0.01,圖11)。這一結(jié)果表明,綿羊卵巢中卵泡生長(zhǎng)發(fā)育成熟后BMP/SMAD通路活性顯著增強(qiáng)。
3 討 論
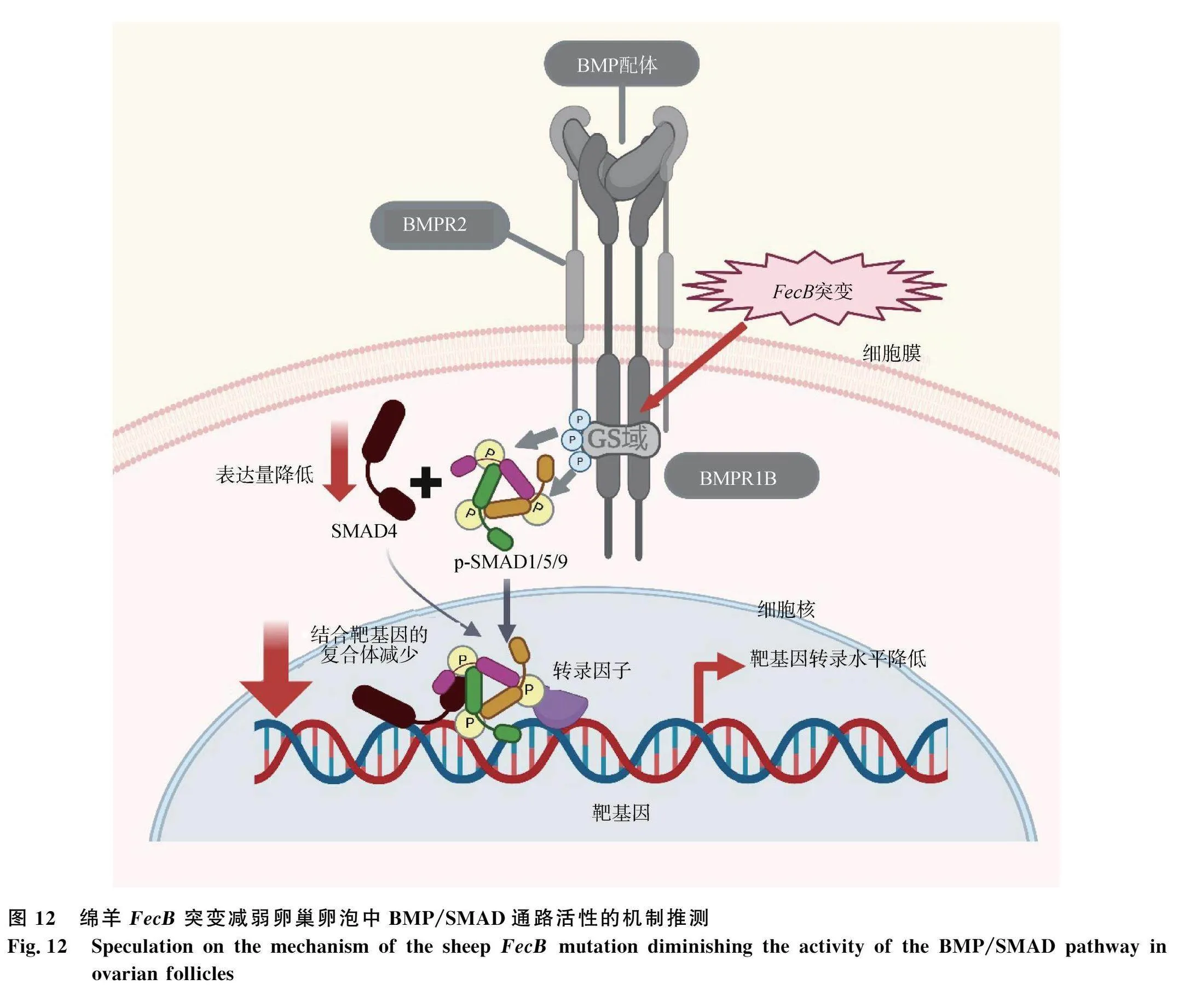
近年來(lái)BMP/SMAD通路在動(dòng)物繁殖中發(fā)揮的作用受到廣泛關(guān)注[17-18]。BMPR1B是該通路的重要受體之一,在發(fā)生FecB突變時(shí),其編碼區(qū)發(fā)生A764G突變,導(dǎo)致氨基酸序列中第294位谷氨酰胺被置換為精氨酸[3,19-20],有學(xué)者推測(cè)該突變可能導(dǎo)致抑制蛋白FKBP1A無(wú)法全部解離,致使其GS區(qū)域暴露的可磷酸化位點(diǎn)減少,抑制該通路的活性[2]。
雖然前期研究表明,F(xiàn)ecB突變參與到了BMP/SMAD通路活性的調(diào)節(jié),但是FecB突變對(duì)該通路活性及相關(guān)蛋白表達(dá)量的具體影響未見(jiàn)報(bào)道。近年來(lái)Fabre等[12]在FecB野生型和突變型卵泡顆粒細(xì)胞中添加BMP配體激活通路后,探究了FecB突變對(duì)BMPR1B受體功能的影響,研究發(fā)現(xiàn),不論是在野生型還是突變型卵泡中,添加BMP配體后,被激活的BMP/SMAD通路均顯著抑制了顆粒細(xì)胞中孕酮的分泌;其次,在添加相同配體量的情況下,與大卵泡相比,小卵泡抑制孕酮分泌的作用更強(qiáng);并且,通過(guò)對(duì)比小卵泡中FecB野生型和突變型顆粒細(xì)胞中BMPR1B對(duì)配體的反應(yīng)活性,發(fā)現(xiàn)FecB突變降低了BMPR1B對(duì)BMP配體的反應(yīng)能力,致使BMP/SMAD通路活性受到改變,突變型顆粒細(xì)胞抑制分泌孕酮的能力降低,最終導(dǎo)致該通路中顆粒細(xì)胞孕酮水平更高,暗示FecB突變對(duì)BMP/SMAD通路活性有調(diào)控作用。另有研究發(fā)現(xiàn),SMADs蛋白家族在顆粒細(xì)胞增殖與凋亡、卵泡發(fā)育和排卵周期方面發(fā)揮重要作用[17-18,21-23]。SMAD4是BMP/SMAD通路中重要成員,它可與磷酸化后的SMAD1/5/9結(jié)合形成復(fù)合物,進(jìn)而易位到細(xì)胞核中激活下游基因的轉(zhuǎn)錄[24]。前期研究表明,在FecB突變型綿羊卵巢內(nèi)BMP/SMAD信號(hào)通路中SMADs mRNA表達(dá)量顯著低于野生型[25]。在本研究中,通過(guò)比較不同F(xiàn)ecB基因型綿羊卵泡中BMP/SMAD通路活性和相關(guān)蛋白表達(dá)量發(fā)現(xiàn),突變型大卵泡和小卵泡中SMAD4的表達(dá)量均顯著降低,并且突變型小卵泡中SMAD1/5/9的磷酸化水平相較野生型顯著降低。綜合以上結(jié)果提出以下推測(cè)(圖12):與 FecB野生型卵泡相比,F(xiàn)ecB突變型卵泡中SMAD4表達(dá)量的降低將導(dǎo)致BMP/SMAD通路中結(jié)合到下游靶基因區(qū)域的SMAD4-SMAD1/5/9蛋白復(fù)合物減少,啟動(dòng)下游靶基因轉(zhuǎn)錄的能力將減弱,即突變可抑制通路活性,并且在突變型小卵泡中SMAD1/5/9的磷酸化水平顯著低于野生型,因此結(jié)合到下游基因靶區(qū)域的復(fù)合體的磷酸化水平更低,抑制通路的作用將更強(qiáng)。
另有學(xué)者通過(guò)測(cè)量不同F(xiàn)ecB基因型卵巢對(duì)促性腺激素刺激的反應(yīng),發(fā)現(xiàn)攜帶FecB突變的卵泡直徑更小,并通過(guò)抑制內(nèi)源性促性腺激素分泌的方式刺激排卵[26-27]。FecB突變會(huì)降低卵泡顆粒細(xì)胞的增殖水平,導(dǎo)致卵泡中顆粒細(xì)胞數(shù)量降低[28-29],提高了對(duì)FSH的敏感性,該通路抑制FSH信號(hào)接受的能力減弱,卵泡提前成熟[3,26]。此外,在顆粒細(xì)胞中,BMP/SMAD通路對(duì)FSH通路的抑制水平降低,使得顆粒細(xì)胞對(duì)FSH產(chǎn)生更高的敏感性[30],而且這些依賴(lài)于FSH激素分化的分子標(biāo)記物均出現(xiàn)在直徑更小的卵泡中[26]。Fabre等[26]證實(shí),攜帶FecB突變母羊的卵泡顆粒細(xì)胞在直徑較小的卵泡中對(duì)LH更為敏感,且LHCGR的mRNA水平顯著高于野生型。作為卵泡成熟的標(biāo)志,LHCGR的表達(dá)水平受到FecB的影響,導(dǎo)致突變型卵巢組織提前排卵,因此突變型卵巢上的成熟卵泡往往具有更小的卵泡直徑[31-33]。本研究同樣發(fā)現(xiàn),F(xiàn)ecB突變型卵巢上的成熟卵泡直徑明顯小于野生型的成熟卵泡直徑。
近年來(lái)研究表明,BMP/SMAD通路活性及通路相關(guān)蛋白會(huì)隨著卵泡發(fā)育和顆粒細(xì)胞的增殖發(fā)生改變[13,34]。針對(duì)BMP受體和SMADs在豬卵泡發(fā)育過(guò)程中的表達(dá)情況,王偉[13]對(duì)比了豬大、小卵泡顆粒細(xì)胞中相關(guān)基因的mRNA水平,發(fā)現(xiàn)在大卵泡的顆粒細(xì)胞中SMAD1、SMAD5、SMAD9的表達(dá)水平有顯著增加的趨勢(shì)。Wei等[35]利用RT-PCR探究8個(gè)不同發(fā)育階段鵝卵泡中SMADs的表達(dá)水平,發(fā)現(xiàn)與未成熟卵泡相比,排卵前卵泡中SMAD1和SMAD4的表達(dá)量顯著升高。同樣,徐夢(mèng)思等[36]利用RT-PCR的方法,探究TGFβ-SMAD信號(hào)通路相關(guān)基因在豬不同大小卵泡中的表達(dá)規(guī)律,發(fā)現(xiàn)與小卵泡相比,大卵泡中SMAD4基因的表達(dá)量顯著升高。Du等[37]敲除豬顆粒細(xì)胞中的SMAD4基因,發(fā)現(xiàn)敲除SMAD4后顆粒細(xì)胞中FSHR的表達(dá)量顯著低于正常水平,表明SMAD4在豬顆粒細(xì)胞中正向調(diào)節(jié)FSHR的表達(dá),發(fā)揮促進(jìn)卵泡發(fā)育和顆粒細(xì)胞增殖的作用。另外,BMPR1B作為該通路重要受體之一,其表達(dá)量在大鼠卵泡的早期發(fā)育過(guò)程中隨著卵泡的發(fā)育不斷增加[18]。本研究發(fā)現(xiàn),與小卵泡相比,綿羊成熟大卵泡中SMAD1/5/9的磷酸化水平以及BMPR1B、SMAD4和SMAD1/5/9的表達(dá)量均顯著提高,這一結(jié)果證明在綿羊卵泡發(fā)育過(guò)程中BMP/SMAD通路的活性顯著增高。
4 結(jié) 論
與野生型卵泡相比,F(xiàn)ecB突變型大卵泡和小卵泡中SMAD4的表達(dá)量均顯著降低,并且在FecB突變型小卵泡中SMAD1/5/9磷酸化水平顯著降低,暗示發(fā)生FecB突變的大、小卵泡中結(jié)合到基因組靶區(qū)域的SMAD4-SMAD1/5/9蛋白復(fù)合物均相對(duì)較少,即通路活性降低,而且在小卵泡中由于較低的SMAD1/5/9磷酸化水平,BMP/SMAD通路的活性更低。另一方面,與小卵泡相比,綿羊成熟大卵泡中BMPR1B和SMAD4蛋白表達(dá)量及SMAD1/5/9磷酸化程度更高,說(shuō)明綿羊卵泡生長(zhǎng)發(fā)育成熟后BMP/SMAD通路活性顯著增強(qiáng)。
參考文獻(xiàn)(References):
[1] XU Y F,LI E L,HAN Y D,et al.Differential expression of mRNAs encoding BMP/Smad pathway molecules in antral follicles of high- and low-fecundity Hu sheep[J].Anim Reprod Sci,2010,120(1-4):47-55.
[2] 龔一鳴,王翔宇,賀小云,等.綿羊FecB突變對(duì)BMPR1B活性及BMP/SMAD通路的影響研究進(jìn)展[J].遺傳,2023,45(4):295-305.
GONG Y M,WANG X Y,HE X Y,et al.Progress on the effect of FecB mutation on BMPR1B activity and BMP/SMAD pathway in sheep[J].Hereditas,2023,45(4):295-305.(in Chinese)
[3] MULSANT P,LECERF F,F(xiàn)ABRE S,et al.Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mrino ewes[J].Proc Natl Acad Sci U S A,2001,98(9):5104-5109.
[4] LI Q L.Inhibitory SMADs:potential regulators of ovarian function[J].Biol Reprod,2015,92(2):50.
[5] YAO Y L,REHEMAN A,XU Y F,et al.miR-125b contributes to ovarian granulosa cell apoptosis through targeting BMPR1B,a major gene for sheep prolificacy[J].Reprod Sci,2019,26(2):295-305.
[6] SHIMIZU T,KAYAMORI T,MURAYAMA C,et al.Bone morphogenetic protein (BMP)-4 and BMP-7 suppress granulosa cell apoptosis via different pathways:BMP-4 via PI3K/PDK-1/Akt and BMP-7 via PI3K/PDK-1/PKC[J].Biochem Biophys Res Commun,2012,417(2):869-873.
[7] WRANA J L.TGF-β receptors and signalling mechanisms[J].Miner Electrolyte Metab,1998,24(2-3):120-130.
[8] HUSE M,CHEN Y G,MASSAGU "J,et al.Crystal structure of the cytoplasmic domain of the type I TGF β receptor in complex with FKBP12[J].Cell,1999,96(3):425-436.
[9] GALAT A.Functional diversity and pharmacological profiles of the FKBPs and their complexes with small natural ligands[J]. Cell Mol Life Sci,2013,70(18):3243-3275.
[10] MASSAGU "J,BLAIN S W,LO R S.TGFβ signaling in growth control,cancer,and heritable disorders[J].Cell,2000,103(2):295-309.
[11] ATTISANO L,WRANA J L.Signal transduction by members of the transforming growth factor-β superfamily[J].Cytokine Growth Factor Rev,1996,7(4):327-339.
[12] FABRE S,PIERRE A,PISSELET C,et al.The Booroola mutation in sheep is associated with an alteration of the bone morphogenetic protein receptor-IB functionality[J].J Endocrinol,2003,177(3):435-444.
[13] 王 偉.BMP/Smad信號(hào)通路對(duì)豬卵泡顆粒細(xì)胞的影響[D].南京:南京農(nóng)業(yè)大學(xué),2010.
WANG W.Effect of BMP/SMAD signaling on porcine follicular granulosa cells[D].Nanjing:Nanjing Agricultural University,2010.(in Chinese)
[14] WANG X Y,GUO X F,HE X Y,et al.Effects of FecB mutation on estrus,ovulation,and endocrine characteristics in small tail han sheep[J].Front Vet Sci,2021,8:709737.
[15] HIRSCHHORN T,LEVI-HOFMAN M,DANZIGER O,et al.Differential molecular regulation of processing and membrane expression of Type-I BMP receptors:implications for signaling[J].Cell Mol Life Sci,2017,74(14):2645-2662.
[16] SCHNEIDER C A,RASBAND W S,ELICEIRI K W.NIH Image to ImageJ:25 years of image analysis[J].Nat Methods,2012, 9(7):671-675.
[17] ZHENG X L,ZHENG Y Q,QIN D X,et al.Regulatory role and potential importance of GDF-8 in ovarian reproductive activity[J].Front Endocrinol (Lausanne),2022,13:878069.
[18] ERICKSON G F,SHIMASAKI S.The spatiotemporal expression pattern of the bone morphogenetic protein family in rat ovary cell types during the estrous cycle[J].Reprod Biol Endocrinol,2003,1:9.
[19] SOUZA C J H,MACDOUGALL C,CAMPBELL B K,et al.The Booroola (FecB) phenotype is associated with a mutation in the bone morphogenetic receptor type 1 B (BMPR1B) gene[J].J Endocrinol,2001,169(2):R1-R6.
[20] WILSON T,WU X Y,JUENGEL J L,et al.Highly prolific Booroola sheep have a mutation in the intracellular kinase domain of bone morphogenetic protein IB receptor (ALK-6) that is expressed in both oocytes and granulosa cells[J].Biol Reprod,2001,64(4): 1225-1235.
[21] CHU Y L,XU Y R,YANG W X,et al.The role of FSH and TGF-β superfamily in follicle atresia[J].Aging (Albany NY),2018,10(3):305-321.
[22] OTSUKA F,MOORE R K,SHIMASAKI S.Biological function and cellular mechanism of bone morphogenetic protein-6 in the ovary[J].J Biol Chem,2001,276(35):32889-32895.
[23] JIAO Y,JIANG T T,LIN Q Y,et al.Molecular characterization of the follicular development of BMP15-edited pigs[J].Reproduction,2023,166(4):247-261.
[24] CHAIKUAD A,BULLOCK A N.Structural basis of intracellular TGF-β signaling:receptors and smads[J].Cold Spring Harb Perspect Biol,2016,8(11):a022111.
[25] BAHIRE S V,RAJPUT P K,KUMAR V,et al.Quantitative expression of mRNA encoding BMP/SMAD signalling genes in the ovaries of Booroola carrier and non-carrier GMM sheep[J].Reprod Domest Anim,2019,54(10):1375-1383.
[26] FABRE S,PIERRE A,MULSANT P,et al.Regulation of ovulation rate in mammals:contribution of sheep genetic models[J].Reprod Biol Endocrinol,2006,4:20.
[27] 種玉晴.綿羊產(chǎn)羔數(shù)性狀的分子遺傳機(jī)理研究[D].武漢:華中農(nóng)業(yè)大學(xué),2021.
ZHONG Y Q.Study on molecular genetic mechanism of litter size in sheep[D].Wuhan:Huazhong Agricultural University,2021.(in Chinese)
[28] MONTGOMERY G W,GALLOWAY S M,DAVIS G H,et al.Genes controlling ovulation rate in sheep[J].Reproduction, 2001,121(6):843-852.
[29] MONTGOMERY G W,MCNATTY K P,DAVIS G H.Physiology and molecular genetics of mutations that increase ovulation rate in sheep[J].Endocr Rev,1992,13(2):309-328.
[30] SHACKELL G H,HUDSON N L,HEATH D A,et al.Plasma gonadotropin concentrations and ovarian characteristics in Inverdale ewes that are heterozygous for a major gene (FecX1) on the X chromosome that influences ovulation rate[J].Biol Reprod,1993,48(5):1150-1156.
[31] MCNATTY K P,HEATH D A,CLARK Z,et al.Ovarian characteristics in sheep with multiple fecundity genes[J].Reproduction, 2017,153(2):233-240.
[32] JUENGEL J L,F(xiàn)RENCH M C,QUIRKE L D,et al.Differential expression of CART in ewes with differing ovulation rates[J].Reproduction,2017,153(4):471-479.
[33] MCNATTY K P,HEATH D A,HUDSON N L,et al.Gonadotrophin-responsiveness of granulosa cells from bone morphogenetic protein 15 heterozygous mutant sheep[J].Reproduction,2009,138(3):545-551.
[34] ONGARO L,SCHANG G,HO C C,et al.TGF-β superfamily regulation of follicle-stimulating hormone synthesis by gonadotrope cells:is there a role for bone morphogenetic proteins?[J].Endocrinology,2019,160(3):675-683.
[35] WEI C C,CHEN X Y,PENG J Z,et al.BMP4/SMAD8 signaling pathway regulated granular cell proliferation to promote follicle development in Wanxi white goose[J].Poult Sci,2023,102(1):102282.
[36] 徐夢(mèng)思,黃 濤,劉麗娟,等.TGFβ-SMAD信號(hào)通路基因在梅山豬與杜洛克豬不同級(jí)別卵泡中的表達(dá)分析[J].家畜生態(tài)學(xué)報(bào),2016,37(2):12-18.
XU M S,HUANG T,LIU L J,et al.Expression patterns of genes of TGFβ-SMAD signaling pathway in meishan and duroc follicle of different size[J].Journal of Domestic Animal Ecology,2016,37(2):12-18.(in Chinese)
[37] DU X,ZHANG L F,LI X Y,et al.TGF-β signaling controls FSHR signaling-reduced ovarian granulosa cell apoptosis through the SMAD4/miR-143 axis[J].Cell Death Dis,2016,7(11):e2476.
(編輯 郭云雁)
