中國荷斯坦牛繁殖性狀的基因型與環境互作
2024-10-14 00:00:00師睿李珊珊張海亮路海博閆青霞張毅陳紹祜王雅春
畜牧獸醫學報 2024年9期



摘 要: 旨在估計奶牛繁殖性狀在不同地區的遺傳參數,檢測同一繁殖性狀在不同地區之間的基因型與環境互作(G×E)效應。本研究利用全國6個地區2 064個牧場2005至2022年的荷斯坦牛群繁殖記錄,計算了2個重要繁殖性狀:初產日齡(AFC)和產犢間隔(CI),共包含1 787 590和2 476 422條表型數據。同時對該原始表型數據進行詳細的質控和分組。隨后,通過BLUPF90軟件的airemlf90模塊利用單性狀動物模型和重復力模型對6個地區的2個繁殖性狀進行了遺傳分析,使用雙性狀動物和重復力模型估計同一性狀不同地區之間的遺傳相關,作為基因型與環境之間效應(G×E)的檢測指標。結果表明,研究所設定的質控條件能夠剔除分布異常的表型值;AFC的遺傳力較高且在地區間差異較大(0.06~0.40),而CI的遺傳力較低且各地區間差異較小(0.02~0.04);在大多數地區組合下,2個繁殖性狀皆檢測到了顯著的G×E效應(Plt;0.05)。綜上,同一繁殖性狀在不同地區的遺傳表現存在差異,且部分區域之間存在顯著的G×E效應。因此,對我國奶牛繁殖性狀進行遺傳改良時需考慮區域性差異及G×E效應對遺傳進展的影響。
關鍵詞: 奶牛;繁殖性狀;遺傳參數;基因型與環境互作
中圖分類號:S823.2; S823.3
文獻標志碼:A 文章編號: 0366-6964(2024)09-3968-10
Genotype by Environment Interaction of Fertility Traits for the Holstein Cattle in China
SHI" Rui" LI" Shanshan1, ZHANG" Hailiang1, LU" Haibo1,3, YAN" Qingxia4, ZHANG" Yi1, CHEN" Shaohu4, WANG" Yachun1*
(1.College of Animal Science and Technology,China Agricultural University, Beijing 100193,
China;
2.Wageningen University and Research,Wageningen 6708PB,Netherlands;
3.Beijing Consortium for Innovative Bio-Breeding, Beijing 101206, China;
4.Dairy Association of China, Beijing 100193," China)
Abstract:" This study aimed to estimate the genetic parameters of fertility traits in different regions, and to test the effect of genotype by environment interactions (G×E) across regions. The raw fertility data were collected from 2 064 Holstein dairy farms in 6 regions of China, including records from 2005 to 2022. A total of 1 787 590 and 2 476 422 phenotypic records were derived from the raw data for age at first calving (AFC), and calving interval (CI). Meanwhile, these phenotypes data were quality controlled and separated into different datasets for subsequent analysis. The statistical models used in this study were all incorporated in the airemlf90 module of BLUPF90. Univariate animal (and repeatability) models were used for estimating genetic parameters for these 2 traits in 6 regions, while bivariate animal (and repeatability) models were used to estimate genetic correlations between the regions, which are the indicators of G×E. The results indicated that the proposed quality control steps could filter abnormal phenotypes records. The heritabilities of AFC were relatively high and varied across regions (0.06-0.40), whereas the heritabilities of CI were low but remained similar across regions (0.02-0.04). Significant G×E effect (Plt;0.05) was observed across most of the regions for both traits. In conclusion, the genetic parameters of the same fertility trait varied across the regions in China, and significant G×E effect was detected for the trait in partial regions. Therefore, it is highly recommended to consider the impacts of regional difference and G×E effect on genetic advance when breeding for fertility traits of dairy cattle in China.
Key words: dairy cows; fertility traits; genetic parameters; genotype by environment interaction
*Corresponding author:" WANG Yachun, E-mail: wangyachun@cau.edu.cn
繁殖性狀是奶牛最為重要的功能性狀之一,其屬于低遺傳力性狀,即受遺傳因素以外的影響較大[1]。對奶牛產奶性能的高度選擇,造成了牛群繁殖能力下降在高產奶牛群體中尤為明顯[2]。繁殖性狀的衰退導致了母牛輸精、獸醫治療和被動淘汰率的增加,對牧場會造成嚴重的經濟損失[3-4]。
隨著各國逐漸意識到繁殖性狀的重要性,對繁殖性狀的遺傳分析也逐步展開。Miglior等[5]報道了15個國家的選擇指數,繁殖性狀在新指數的投入使用下,取得了明顯的遺傳進展。相比之下,我國還未能將此類性狀納入選擇指數。由于早期繁殖數據記錄體系不完善,各地區數據資料收集有所差別,加之繁殖表型記錄質控困難,導致我國目前對繁殖性狀的遺傳評估技術體系尚不完善[6]。但在繁殖性狀逐漸受到重視的大背景下,我國也亟需一套系統性處理繁殖數據的技術流程,便于后續展開區域性乃至全國性的遺傳評估。初產日齡(age at first calving, AFC)可以同時反映母牛個體的性成熟早晚和受孕能力。有研究表明,北京地區荷斯坦牛的初產日齡遺傳力為0.25[7],李欣等[8]對寧夏荷斯坦牛群的遺傳力進行了評估(0.37),Zhu等[9]研究表明南方荷斯坦牛初產日齡遺傳力為0.193;Ghavi Hossein-Zadeh[10]研究發現伊朗荷斯坦牛的初產日齡遺傳力為0.34,而Elahi Torshizi[11]的研究則表明伊朗荷斯坦牛的初產日齡遺傳力為0.075;Ojango和Pollott[12]研究表明東非荷斯坦牛初產日齡的遺傳力為0.38;Pirlo等[13]研究指出初產日齡遺傳力的范圍較廣(0.05~0.75)。可見,初產日齡在不同地區的遺傳力呈現差異性,這可能與氣候和牛場管理相關。
近年來,解析不同性狀的基因型與環境互作(genotype by environment interaction, G×E)效應已成為各國研究的一個重點[14-18]。G×E即在不同的環境下相同基因型的表現會有所差別,會導致育種進展出現差異性[19-20]。G×E效應可以通過雙性狀模型進行檢測,通過將同一性狀在不同環境(地區)的表現視為不同性狀,若遺傳相關估計值顯著小于1(Plt;0.05)則視為存在G×E效應[17,21-23]。在Interbull(https://interbull.org/)的跨國遺傳評估中,不同國家之間的繁殖性狀存在著中等或較強程度的G×E。有研究表明,同一國家因養殖模式的不同(例如傳統與有機牧場的區別)會導致產奶性狀和繁殖性狀呈現不同程度的G×E[23-26]。由此可見,隨著一個國家養殖模式的多樣化跨地區遺傳評估的開展,為實現不同情景下育種進展的最大化,G×E在奶牛育種中越顯重要[27-29]。
本研究對全國6個地區中國荷斯坦牛群的繁殖性狀進行了詳細質控和遺傳參數估計,并檢測了繁殖性狀各地區之間的G×E效應。旨在為我國奶牛繁殖性狀的系統性評估方案提供參考、為制定區域性和全國性的育種方案打下基礎。
1 材料與方法
1.1 試驗數據
本研究所使用的原始出生及產犢記錄來源于中國奶業協會數據庫,共包含北京、天津、上海、河北、河南和山東6個地區的2 064個規模化奶牛場。數據收集時間為2005年1月至2022年2月,共保留出生記錄2 351 795條和產犢記錄4 895 587條。系譜數據根據中國奶牛數據中心(https://www.holstein.org.cn)數據庫、牧場牛只出生記錄和產犢記錄進行整理追溯,最終包含2 972 604頭個體,其中有25 711頭公牛。本研究主要對2個重要繁殖性狀進行分析:初產日齡(age at first calving, AFC, d):牛只初次產犢的日齡;產犢間隔(calving interval, CI, d):成母牛連續兩次產犢的間隔天數。根據數據分布情況,本研究保留了1~7胎的數據用于計算CI。
1.2 數據質控及分組
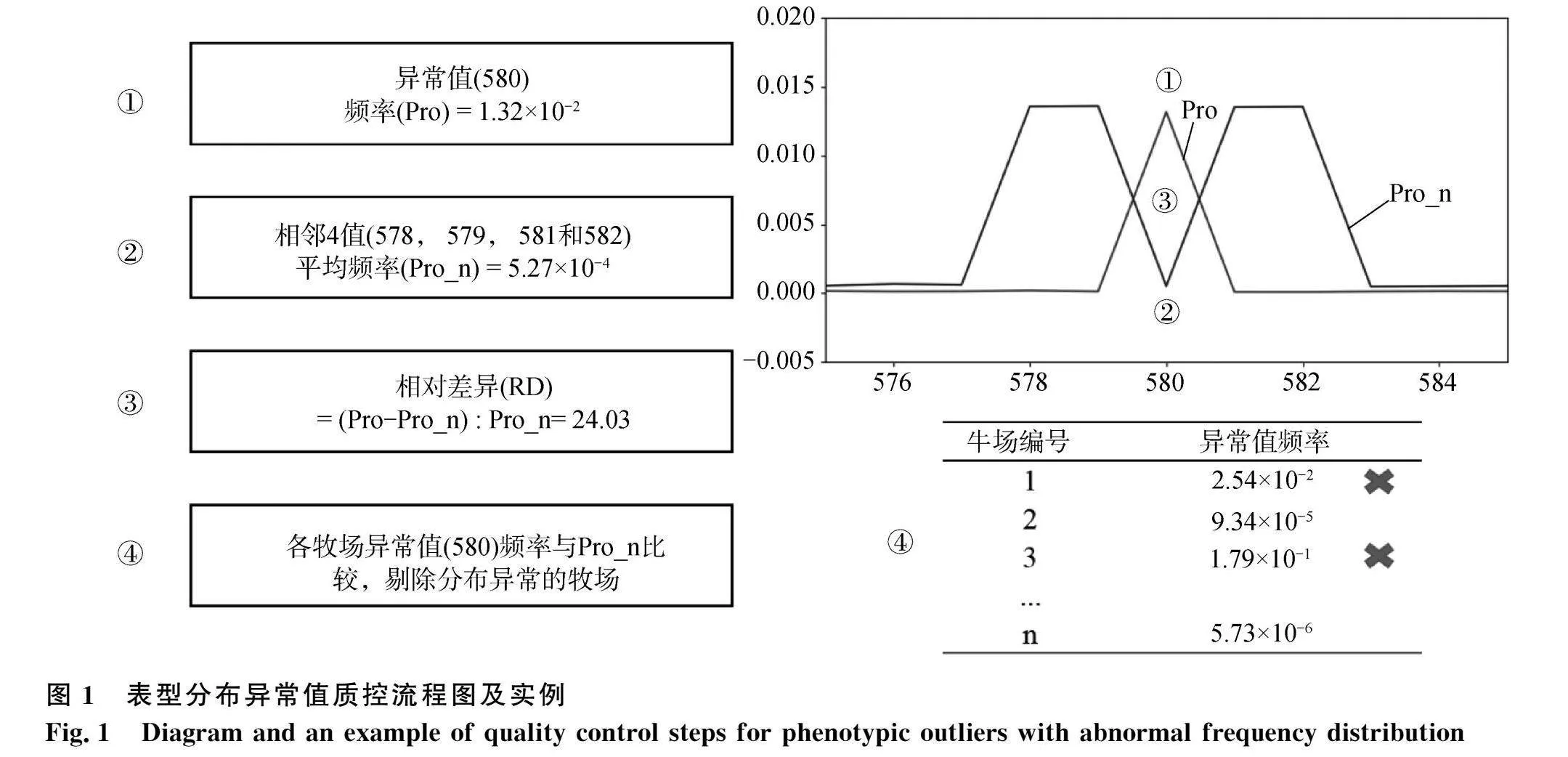
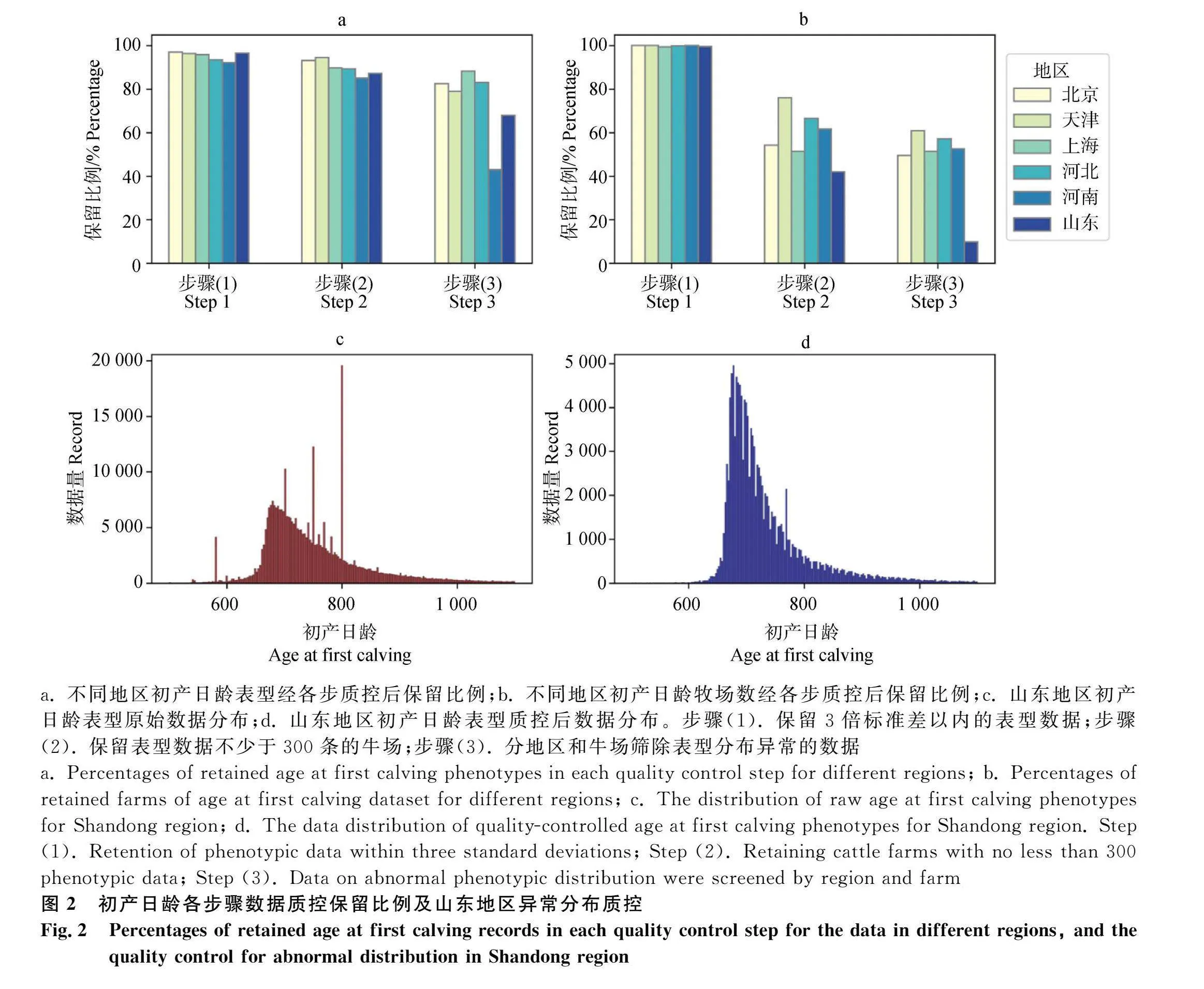
各繁殖性狀表型質量控制包含3步:1)保留3倍標準差以內的表型數據;2)保留表型數據不少于300條的牛場;3)分地區和牛場篩除表型分布異常的數據。根據原始數據分布,研究發現部分數據分布存在異常峰值,因此在第“3)”步中引入異常峰值的判定篩除(圖1)。異常值的判定基于該值頻率(pobability, Pro;①)和該值相鄰4個值的平均頻率(probability nearby, Pro_n;②)之間的相對差異(relative difference, RD;③)。如果該值的RD大于0(即Pro高于Pro_n;④),則判定測試地區的性狀表型在該值的分布存在異常峰值。隨后計算地區中各牛場表型異常值的頻率,若基于牛場數據所得的表型異常值頻率高于基于地區數據的Pro_n,則判定測試牛場的性狀表型分布異常,并剔除該牛場的所有數據。例如,表格內1號和3號牛場異常值頻率(2.54×10-2和1.79×10-1)高于Pro_n(5.27×10-4),因此剔除這些牛場的所有數據。若存在多個異常值,則取各異常值判定結果的合集剔除相應牧場。
質控完成后,對于任一性狀,本研究將所得數據分為3組用于后續分析:(a)所有數據(1個數據集);(b)各地區數據(共6個數據集);(c)兩兩地區合并數據(共15個數據集)。為探究公牛遺傳聯系對G×E檢測的影響,本研究進一步統計了各個地區之間母牛群體的父親重合比例,并篩選出這些父親的女兒表型,再對數據進一步質控,對AFC和CI分別對應保留出生場-年-季效應或前次產犢場-年-季效應不少于10條的記錄。該數據組記為(d),共包含15個數據集。
1.3 統計模型
對于數據(a)和(b),本研究使用單性狀動物(重復力)模型計算全國及各地區不同性狀方差組分及遺傳力,模型如下所示:
y=Xb+Za+Wc+e
其中,y是表型值向量;X、Z和W分別是固定效應、加性遺傳效應和永久環境效應(AFC不含此矩陣)的關聯矩陣;b是固定效應向量(對于AFC為個體出生場-年-季效應;對于CI為個體前次產犢場-年-季效應和胎次效應);a是隨機加性遺傳效應向量;c是隨機永久環境效應向量(AFC不含此向量);e是隨機殘差向量。其中,a~N(0,Aσ2a),A矩陣為基于系譜構建的親緣關系矩陣,σ2a為加性遺傳方差;c服從分布c~N(0,Iσ2pe),I為單位矩陣,σ2pe為永久環境方差;e服從分布e~N(0,Iσ2e),σ2e為殘差方差。
為評估同一性狀不同地區的G×E效應,本研究針對數據(c)和(d)使用雙性狀動物(重復力)模型估計兩兩地區之間的遺傳相關,模型如下所示:
y1
y2=X10
0X2b1
b2+Z10
0Z2a1
a2+
W10
0W2c1
c2+e1
e2
其中效應均與單性狀模型一致,下標指示不同地區。各效應服從的分布為:a1a2~N0,Aσ2a1σa1a2σa1a2σ2a2,c1c2~N0,Iσ2pe100σ2pe2,以及e1e2~N0,Iσ2e100σ2e2。其中,σa1a2為兩地區間的加性遺傳協方差,兩地區間的永久環境協方差及殘差協方差假設為0。
本研究所有計算均基于BLUPF90軟件[30]的airemlf90模塊,采用平均信息約束最大似然法(average information restricted maximum likelihood, AIREML)估計各性狀的方差組分。AFC的遺傳力計算公式為:h2=σ2aσ2a+σ2e;CI的遺傳力計算公式為:h2=σ2aσ2a+σ2pe+σ2e;CI的重復力計算公式為:r2=σ2a+σ2peσ2a+σ2pe+σ2e;兩地區間的遺傳相關計算公式為:rg=σa1a2σ2a1×σ2a2。遺傳力及相關系數的標準誤依據泰勒展開式進行計算[31]。遺傳相關顯著的標準為相關系數與0存在顯著差異(Plt;0.05),G×E效應的判定標準為同一性狀在不同地區之間的遺傳相關顯著小于1(Plt;0.05)[23]。
2 結 果
2.1 表型數據質控
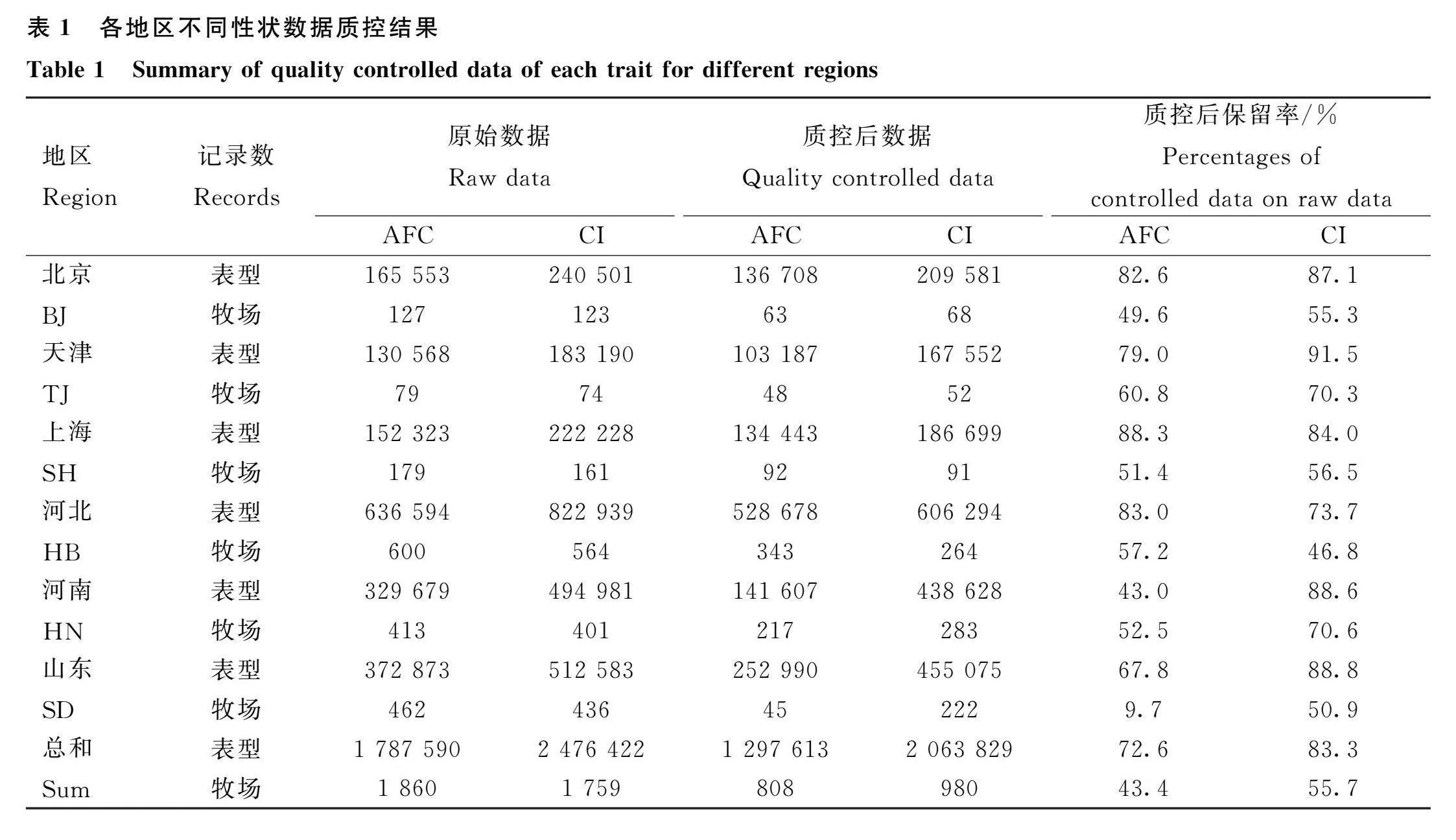
經過數據質控后,不同性狀各地區表型數據量及牧場數見表1。對于AFC,各地區經質控后的表型數據占原始數據的比例總體均值為72.6%,各地區中數據保留率最低為河南(43.0%),最高為上海(88.3%)。質控后,牧場數量總體保留率為43.4%,山東地區牧場保留率(9.7%)大幅低于其他地區(49.6%~60.8%)。對于CI,質控后表型數據及牧場的總體保留率分別為88.3%和55.7%。其中,河北地區的表型和牧場保留率最低,分別為73.7%和46.8%。
因山東地區的AFC數據質控后牛場保留率極低,圖2c-d展示了山東地區AFC異常分布質控的過程,可見質控步驟(3)篩除了山東地區大量的牧場。通過檢查原始數據分布,發現該地區AFC出現了多個分布異常的峰值(例如AFC=800)。經過質控后,數據分布與整體數據及其他地區數據較為一致。其他地區的數據未出現大量牧場或表型被篩除的情況,且各性狀表型分布較為一致。
2.2 各地區中國荷斯坦牛繁殖性狀遺傳參數
針對研究所選2個繁殖性狀,不同地區性狀表型的描述性統計、方差組分、遺傳力(標準誤)及重復力(標準誤)估計值如表2所示。本研究所估計的不同性狀不同地區的遺傳力和重復力均顯著大于0。結果表明,AFC的遺傳力普遍高于CI,且地區間差異較大,最低為上海地區(0.06),而最高為河北地區(0.40)。其差異主要來源于加性遺傳方差,上海地區AFC的加性方差(242.41)遠小于河北地區(2 366.40),但殘差方差則較為一致(3 619.70~3 820.40)。不同數據集CI的方差組分及遺傳參數的估計值皆較為一致。對于CI,遺傳力范圍為0.02~0.04,重復力范圍為0.06~0.08,加性遺傳方差的范圍為72.05(上海)~141.34(河北),殘差方差則在2 304.90(河南)~3 800.60(北京)之間。
2.3 不同地區繁殖性狀之間基因環境互作
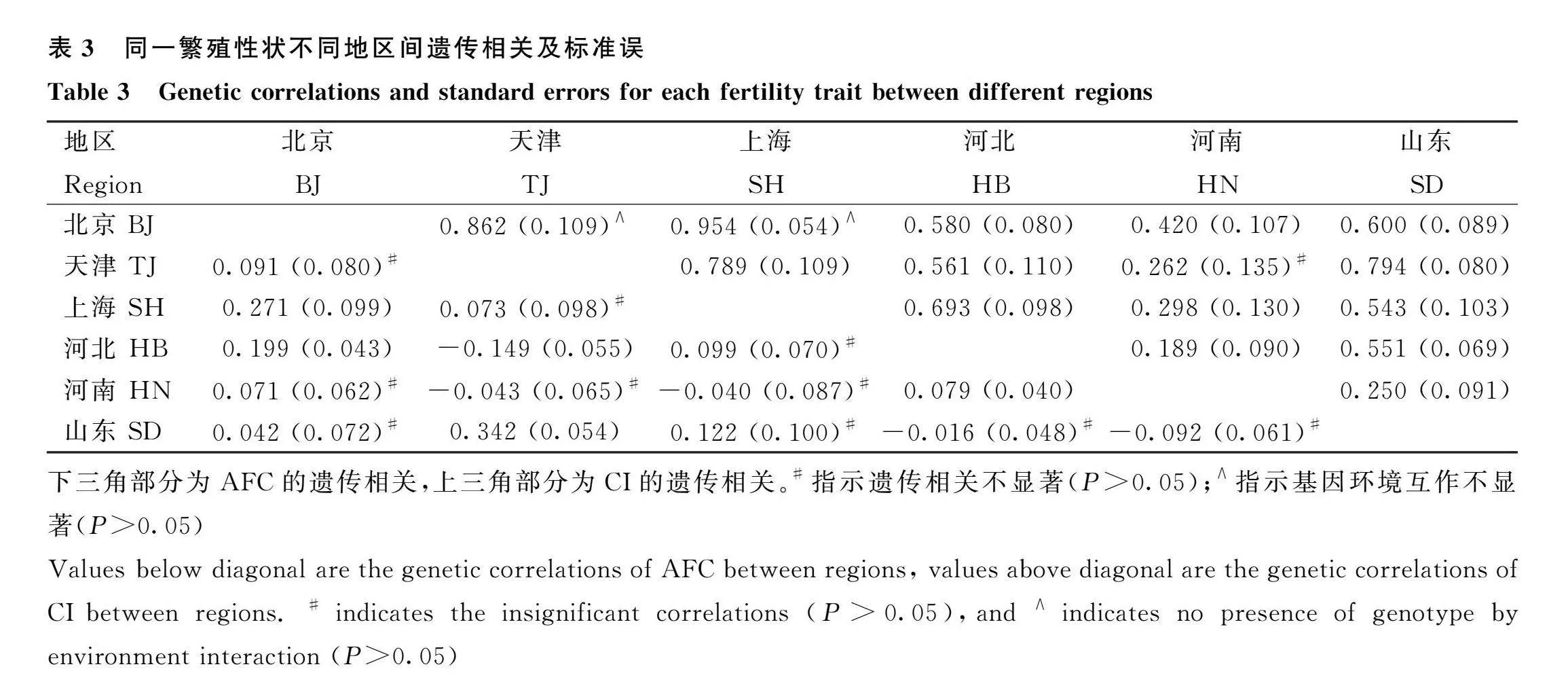
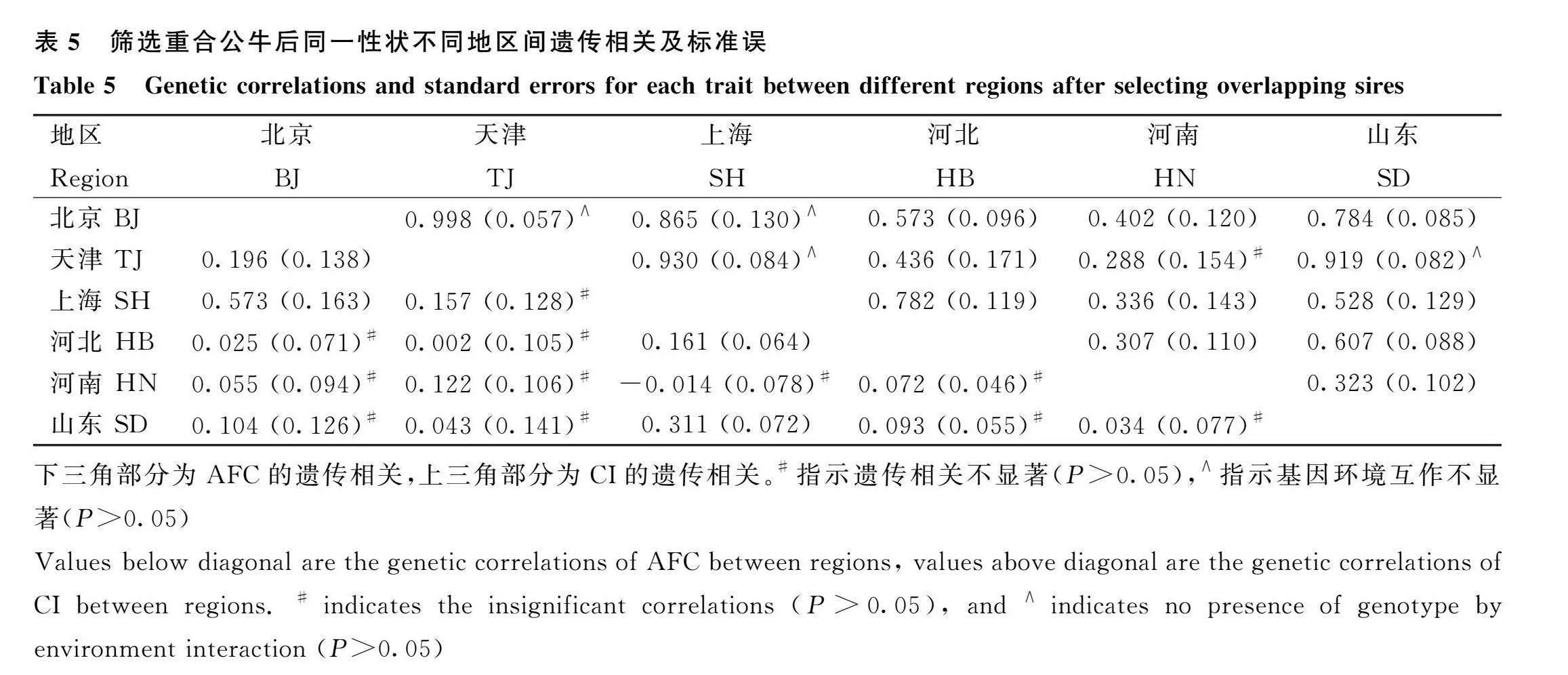
通過雙性狀模型所得同一性狀不同地區之間的遺傳相關及G×E效應的估計結果見表3。結果表明,AFC地區之間的遺傳相關水平較低,范圍為-0.149(天津-河北)~0.342(天津-山東),且有10個地區之間的遺傳相關估計值不顯著。對于CI,各地區之間遺傳相關差異較大,最低為0.189(河北-河南),最高為0.954(北京-上海),其余地區之間多呈中等遺傳相關,且僅有天津和河南之間的遺傳相關估計值不顯著。根據G×E效應的檢測標準,僅有北京-天津及北京-上海地區之間的CI性狀不存在顯著的G×E效應,其余地區之間的繁殖性狀皆檢測出顯著的G×E效應。
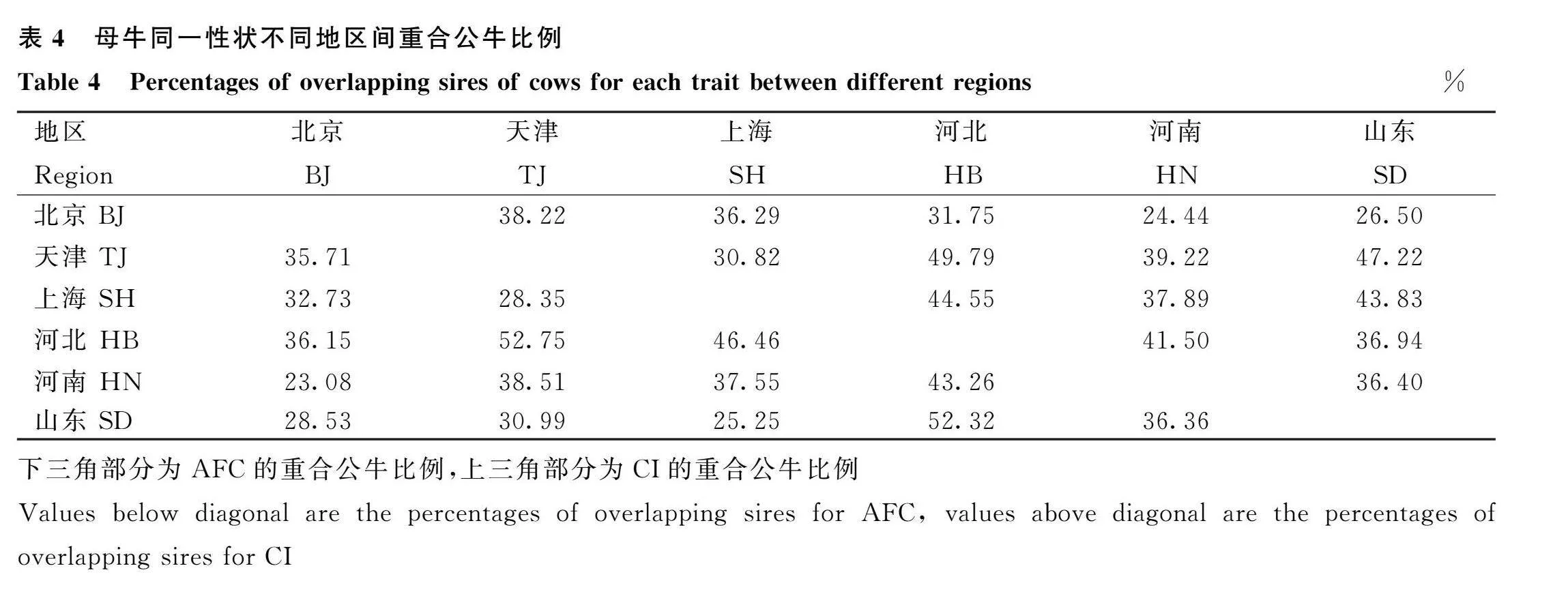
各地區之間母牛同一性狀重合父親的比例見表4,結果表明所研究母牛群體的父親在不同地區之間差異較大。對于AFC,母牛的父親重合比例范圍在23.08%(北京-河南)~52.75%(天津-河北)之間;對于CI,范圍則在24.44%(北京-河南)~49.79%(天津-河北)之間。經過對重合父親的篩選及進一步質控后,數據組(d)的平均保留率為36.28%,地區之間保留率范圍在20%~55%之間。
基于數據(d)和雙性狀模型所得同一性狀不同地區之間的遺傳相關及G×E效應檢測的結果見表5。對于AFC,遺傳相關不顯著的地區組合數增加至11個;除上海-河南(-0.014)外,其余地區組合的AFC相關系數均大于0。對于CI,僅有天津-河南組合的遺傳相關不顯著;而G×E效應不顯著的結果則新增天津-上海和天津-山東兩個地區組合。相比于數據(c)的結果(表3),各地區組合的遺傳相關估計值及標準誤出現不同程度的變化,此變化在AFC中的幅度較大,在CI中則相對較小。
3 討 論
3.1 繁殖性狀的數據記錄及質控
本研究對全國不同地區的不同牧場繁殖記錄進行了系統整理和嚴格質控。盡管大部分表型數據能夠通過質控,但數據質量合格的牧場占比較小,即表型數據量較小的牧場都未能通過質控。此外,各地區數據經質控后,AFC表型的保留比例要明顯小于CI的保留比例(表1)。本研究中,篩除數據量最多的步驟為第(3)步(篩除各場表型分布異常的數據),這說明大部分牧場的繁殖記錄準確性未能達到質控要求。以山東地區的AFC表型為例,數據記錄異常可能是因為牧場管理軟件自動填充(圖2,AFC=800異常峰值)。本研究中質控過程基于前人相關研究[6,32-34],國內外學者也強調了原始數據整理是繁殖性狀準確評估的挑戰之一[6,32-34]。因此,在針對繁殖性狀進行遺傳評估前,應當謹慎、細致地對原始數據進行質控,避免因數據記錄不準確而導致的評估錯誤。
3.2 各地區繁殖性狀遺傳參數差異
本研究評估了中國不同地區荷斯坦母牛AFC和CI的遺傳參數,為其余母牛繁殖性狀在全國范圍內的評估打下了基礎。本研究中,不同地區荷斯坦牛AFC和CI的群體均值雖然有一定差異,但皆與其他國家荷斯坦牛群體的報道相近,且性狀的遺傳力估計值也與前人報道相一致[35-37]。國內相同地區的其他研究表明,北京地區荷斯坦牛的初產日齡為781.11±83.32天,而上海地區為758.73±57.10天[6],可見本研究的結果處于正常范圍。
本研究的AFC遺傳力呈現出地區差異性。這可能與飼養環境和牛場管理有關。因為牧場可以實行不同策略人為控制母牛首次配種日齡,進而間接控制AFC大小(與首次配種僅相差一個妊娠期長度)便于群體管理[13,38]。管理的差異性也體現在AFC較大的表型標準差(表2)。
本研究中CI的遺傳力估計值較低,且各區域之間差異較小。這是因為CI反映了母牛受胎、妊娠及產犢的能力,導致此性狀易受各類環境影響,殘差方差遠高于加性方差(表2)。但由于同一品種牛只妊娠期長度較為一致,且妊娠期占該性狀比例較大,無法人為調控。因此CI在不同地區(牧場)間的差異較小(表2)。這也與其他國內外的報道一致[9-10,39]。
3.3 中國荷斯坦牛繁殖性狀的基因環境互作
本研究的目的是檢測不同地區之間中國荷斯坦牛同一繁殖性狀的G×E效應。雙性狀模型結果顯示,全國不同地區的繁殖性狀表現有所差異。對于AFC,大多數分組之間的遺傳相關不顯著,但所有分組之間都檢測到了G×E效應。而對于CI,大多數分組之間的遺傳相關顯著且檢測到了G×E效應。國內對繁殖性狀G×E的研究較少,陳紫薇等[37]依據配種季節對寧夏地區荷斯坦青年牛繁殖性狀的G×E效應進行了檢測,結果表明多個繁殖性狀在不同季節間存在G×E效應(遺傳相關小于0.8)。國外的研究中,加拿大的研究將傳統牧場和放牧牧場作為不同牧場模式進行CI的G×E效應檢測,其遺傳相關為0.64,但由于其標準誤較大,所以二者的G×E效應并不顯著[40]。而Liu等[23]基于雙性狀模型,對丹麥地區傳統牧場和有機牧場之間繁殖性狀的G×E進行了檢測,發現繁殖性狀存在顯著的G×E效應,其雙性狀遺傳相關取值范圍為0.607~0.957。
根據上述文獻可知,使用雙性狀對荷斯坦牛繁殖性狀進行G×E檢測結果差異較大,以往研究也表明不同環境下同一繁殖性狀的遺傳相關取值較廣,但大部分研究所估計的遺傳相關值均在0.500以上,與本研究中CI的結果相一致。對于AFC,之前研究[6]對全國數據評估時也發現AFC在不同地區之間的遺傳相關系數較低,并且提出了原因可能是所用系譜數據未能建立較準確的遺傳關系。因為在其他研究中,系譜不同群體中也有較高重合率,如劉澳星對丹麥荷斯坦牛的研究中,有機牧場的母牛父親是傳統牧場母牛父親的子集[23]。本研究也對各地區組合的表型數據篩選了重合的母牛父親重新評估(表5)。該結果與未篩選父親的數據相近,但AFC各地區之間的遺傳相關系數仍不一致。因此,AFC在地區間較低的遺傳相關可能由于牧場管理方案及地區差異的因素混雜,導致雙性狀模型未能在不同分組下得到較為一致的遺傳相關系數。此外,本研究中各地區之間母牛父親的重合率較低(表4),經過篩選后的小部分表型數據仍可得到相似估計結果。這也說明清晰的系譜關系能夠保證雙性狀模型估計的準確性。
本研究檢測到的繁殖性狀G×E效應也為我國奶牛育種規劃提供了思路。基于本研究結果,我國在制定奶牛選育的育種目標時,也需考慮跨地區導致的G×E效應,以期在不同環境、管理條件下達到最快的遺傳選擇進展。
4 結 論
本研究以全國不同地區的中國荷斯坦牛為對象,對初產日齡和產犢間隔進行了遺傳分析和G×E效應檢測。研究發現,繁殖性狀的異常表型值能夠通過精確質控篩除,相同繁殖性狀在不同地區的遺傳表現呈現異質性,且奶牛繁殖性狀在我國不同地區廣泛存在G×E效應。因此,選育我國奶牛的繁殖性狀時,應考慮G×E效應的影響,以便在不同地區都能快速提高繁殖性狀的遺傳水平。
參考文獻(References):
[1] BRITT J H.Enhanced reproduction and its economic implications[J].J Dairy Sci,1985,68(6):1585-1592.
[2] LUCY M C.Reproductive loss in high-producing dairy cattle:where will it end?[J].J Dairy Sci,2001,84(6):1277-1293.
[3] SCHMIDTMANN C,THALLER G,KARGO M,et al.Derivation of economic values for German dairy breeds by means of a bio-economic model-with special emphasis on functional traits[J].J Dairy Sci,2021,104(3):3144-3157.
[4] SAMARAWEERA A M,VAN DER WERF J H J,BOERNER V,et al.Economic values for production,fertility and mastitis traits for temperate dairy cattle breeds in tropical Sri Lanka[J].J Anim Breed Genet,2022,139(3):330-341.
[5] MIGLIOR F,FLEMING A,MALCHIODI F,et al.A 100-Year Review:identification and genetic selection of economically important traits in dairy cattle[J].J Dairy Sci,2017,100(12):10251-10271.
[6] 師 睿.中國荷斯坦牛繁殖性狀的基因組預測效果比較[D].北京:中國農業大學,2019.
SHI R.Estimation of genetic parameters and genome-wide association study for female reproductive traits in Chinese Holstein population[D].Beijing:China Agricultural University,2019.(in Chinese)
[7] 劉澳星,郭 剛,王雅春,等.中國荷斯坦牛初產日齡遺傳評估及全基因組關聯分析[J].畜牧獸醫學報,2015,46(3):373-381.
LIU A X,GUO G,WANG Y C,et al.Genetic analysis and genome wide association studies for age at first calving in Chinese Holsteins[J].Acta Veterinaria et Zootechnica Sinica,2015,46(3):373-381.(in Chinese)
[8] 李 欣,周靖航,溫 萬,等.寧夏地區荷斯坦奶牛遺傳參數估計[J].中國畜牧獸醫,2017,44(6):1754-1761.
LI X,ZHOU J H,WEN W,et al.The estimation of genetic parameters of Holstein dairy cows in Ningxia Region[J].China Animal Husbandry amp; Veterinary Medicine,2017,44(6):1754-1761.(in Chinese)
[9] ZHU K,LI T W,LIU D Y,et al.Estimation of genetic parameters for fertility traits in Chinese Holstein of South China[J].Front Genet,2024,14:1288375.
[10] GHAVI HOSSEIN-ZADEH N.Genetic parameters and trends for calving interval in the first three lactations of Iranian Holsteins[J].Trop Anim Health Prod,2011,43(6):1111-1115.
[11] ELAHI TORSHIZI M.Effects of season and age at first calving on genetic and phenotypic characteristics of lactation curve parameters in Holstein cows[J].J Anim Sci Technol,2016,58(1):8.
[12] OJANGO J M,POLLOTT G E.Genetics of milk yield and fertility traits in Holstein-Friesian cattle on large-scale Kenyan farms[J].J Anim Sci,2001,79(7):1742-1750.
[13] PIRLO G,MIGLIOR F,SPERONI M.Effect of age at first calving on production traits and on difference between milk yield returns and rearing costs in Italian Holsteins[J].J Dairy Sci,2000,83(3):603-608.
[14] WATERS D L,CLARK S A,MOGHADDAR N,et al.Genomic analysis of the slope of the reaction norm for body weight in Australian sheep[J].Genet Sel Evol,2022,54(1):40.
[15] MCWHORTER T M,SARGOLZAEI M,SATTLER C G,et al.Single-step genomic predictions for heat tolerance of production yields in US Holsteins and Jerseys[J].J Dairy Sci,2023,106(11):7861-7879.
[16] FREITAS P H F,JOHNSON J S,TIEZZI F,et al.Genomic predictions and GWAS for heat tolerance in pigs based on reaction norm models with performance records and data from public weather stations considering alternative temperature thresholds[J].J Anim Breed Genet,2024,141(3):257-277.
[17] SONG H L,ZHANG Q,DING X D.The superiority of multi-trait models with genotype-by-environment interactions in a limited number of environments for genomic prediction in pigs[J].J Anim Sci Biotechnol,2020,11:88.
[18] SONG H L,ZHANG Q,MISZTAL I,et al.Genomic prediction of growth traits for pigs in the presence of genotype by environment interactions using single-step genomic reaction norm model[J].J Anim Breed Genet,2020,137(6):523-534.
[19] FALCONER D S,MACKAY T F C.Introduction to quantitative genetics[M].4th ed.Harlow,Essex:Longman,1996.
[20] DUCROCQ V,CADET A,PATRY C,et al.Two approaches to account for genotype-by-environment interactions for production traits and age at first calving in South African Holstein cattle[J].Genet Sel Evol,2022,54(1):43.
[21] SUNDBERG T,RYDHMER L,FIKSE W F,et al.Genotype by environment interaction of Swedish dairy cows in organic and conventional production systems[J].Acta Agric Scand Sect A-Anim Sci,2010,60(2):65-73.
[22] ISMAEL A,STRANDBERG E,BERGLUND B,et al.Genotype by environment interaction for activity-based estrus traits in relation to production level for Danish Holstein[J].J Dairy Sci,2016,99(12):9834-9844.
[23] LIU A,SU G,H GLUND J,et al.Genotype by environment interaction for female fertility traits under conventional and organic production systems in Danish Holsteins[J].J Dairy Sci,2019,102(9):8134-8147.
[24] ZHANG Z,KARGO M,LIU A X,et al.Genotype-by-environment interaction of fertility traits in Danish Holstein cattle using a single-step genomic reaction norm model[J].Heredity (Edinb),2019,123(2):202-214.
[25] CHERUIYOT E K,NGUYEN T T T,HAILE-MARIAM M,et al.Genotype-by-environment (temperature-humidity) interaction of milk production traits in Australian Holstein cattle[J].J Dairy Sci,2020,103(3):2460-2476.
[26] SIGDEL A,LIU L,ABDOLLAHI-ARPANAHI R,et al.Genetic dissection of reproductive performance of dairy cows under heat stress[J].Anim Genet,2020,51(4):511-520.
[27] DEKKERS J C M.Multiple trait breeding programs with genotype-by-environment interactions based on reaction norms,with application to genetic improvement of disease resilience[J].Genet Sel Evol,2021,53(1):93.
[28] BENGTSSON C,THOMASEN J R,KARGO M,et al.Emphasis on resilience in dairy cattle breeding:possibilities and consequences[J].J Dairy Sci,2022,105(9):7588-7599.
[29] CAO L,LIU H M,MULDER H A,et al.Genomic breeding programs realize larger benefits by cooperation in the presence of genotype×environment interaction than conventional breeding programs[J].Front Genet,2020,11:251.
[30] MISZTAL I.Complex models,more data:simpler programming?[C]//Proceedings of the Computational Cattle Breeding ′99 Workshop.Tuusula,1999:33.
[31] SU G,LUND M S,SORENSEN D.Selection for litter size at day five to improve litter size at weaning and piglet survival rate[J].J Anim Sci,2007,85(6):1385-1392.
[32] LIU A X,LUND M S,WANG Y C,et al.Variance components and correlations of female fertility traits in Chinese Holstein population[J].J Anim Sci Biotechnol,2017,8:56.
[33] SHI R,BRITO L F,LIU A X,et al.Genotype-by-environment interaction in Holstein heifer fertility traits using single-step genomic reaction norm models[J].BMC Genomics,2021,22(1):193.
[34] WELLER J I,EZRA E,GERSHONI M.Genetic and genomic analysis of age at first insemination in Israeli dairy cattle[J].J Dairy Sci,2022,105(6):5192-5205.
[35] VEERKAMP R F,BEERDA B.Genetics and genomics to improve fertility in high producing dairy cows[J].Theriogenology,2007,68 Suppl 1:S266-S273.
[36] MUUTTORANTA K,TYRISEV "A M,M NTYSAARI E A,et al.Genetic parameters for female fertility in Nordic Holstein and Red Cattle dairy breeds[J].J Dairy Sci,2019,102(9):8184-8196.
[37] 陳紫薇,師 睿,羅漢鵬,等.寧夏地區荷斯坦牛青年牛繁殖性狀遺傳參數估計[J].畜牧獸醫學報,2021,52(2):344-351.
CHEN Z W,SHI R,LUO H P,et al.Estimation of genetic parameters of reproductive traits of Holstein heifers in Ningxia[J].Acta Veterinaria et Zootechnica Sinica,2021,52(2):344-351.(in Chinese)
[38] MOTA L F M,LOPES F B,FERNANDES JNIOR G A,et al.Genome-wide scan highlights the role of candidate genes on phenotypic plasticity for age at first calving in Nellore heifers[J].Sci Rep,2020,10(1):6481.
[39] 任小麗,栗敏杰,白雪利,等.中國荷斯坦牛青年初產年齡和成年母牛產犢間隔遺傳參數估計[J].中國畜牧雜志,2019,55(3):45-49.
REN X L,LI M J,BAI X L,et al.Genetic parameters for age at first calving of heifer and calving interval of cow in Chinese Holstein[J].Chinese Journal of Animal Science,2019,55(3):45-49.(in Chinese)
[40] KEARNEY J F,SCHUTZ M M,BOETTCHER P J.Genotype×environment interaction for grazing vs. confinement.II.Health and reproduction traits[J].J Dairy Sci,2004,87(2):510-516.
(編輯 郭云雁)
