核桃青皮堆肥過程中理化性質及微生物群落的變化
2024-12-27 00:00:00郭金鵬楊霞
經濟林研究 2024年4期


關鍵詞:核桃青皮;堆肥;理化性質;微生物群落多樣性
核桃青皮,是核桃Juglansregia果實最外層的綠色果皮。在核桃果實成熟后青皮被剝離,大量的青皮作為廢料被堆放在田地里,既造成資源浪費,又會對環境造成污染。核桃青皮中的成分具有一定的藥用價值和農用價值[1-3],但核桃青皮中有效成分的提取工藝目前尚未市場化和工業化。目前,核桃青皮用于制備有機肥仍然是其較好的處理方式,具有操作方便、節約成本的特點。前人已經探索了核桃青皮與其他原料混合后制備有機肥的方式及其肥效,證明可以利用堆肥發酵后的核桃青皮制備有機肥[4-7]。了解堆肥過程中微生物群落的變化對于優化堆肥過程、提升堆肥質量具有重要意義,但是目前關于核桃青皮發酵過程中微生物群落的變化情況的研究報道較為鮮見。本研究中擬通過采用核桃青皮作為單一原料進行堆肥發酵,并且分析發酵過程中的養分變化和微生物群落變化情況,探討微生物群落變化對青皮理化指標的影響,旨在為挖掘和篩選較好的核桃青皮堆肥菌劑提供一定的理論參考。
1材料與方法
1.1試驗材料
2022年,在貴州省正安縣瑞溪鎮王大巖核桃園收集供試核桃青皮,用粉碎機對晾干的青皮(含水量8.52%)進行粉碎,將粉碎的青皮過篩(孔徑0.25mm)后在室溫條件下保存,待用。在8—9月份進行發酵,環境溫度22~30℃,環境濕度35%~55%。將粉碎后的核桃青皮粉末按照C、N質量比25∶1加入尿素,混合均勻后裝入長方形泡沫盒里進行堆肥發酵。堆肥后每天記錄堆肥的溫度,根據溫度的升降情況,在堆肥第1、4、11、17、21天對堆肥進行取樣,各階段分別標記為A、B、C、D、E。
1.2試驗方法
1.2.1樣品理化指標測定
采用文獻[8]中的方法測定堆肥的pH值、全氮含量、全磷含量、有效磷含量、有機質含量、全鉀含量、速效鉀含量,采用文獻[9]中的方法測定堆肥的水解氮含量,采用文獻[10]中的方法測定堆肥的腐殖酸含量、含水率和電導率(EC值)。
1.2.2樣品掃描電鏡觀察
將樣品在干燥器中干燥48h后,用庫賽姆(COXEM)EM-30臺式掃描電鏡對樣品進行觀察,記錄樣品的降解情況。
1.2.3樣品微生物群落多樣性分析
用細菌16srRNA通用引物338F(5′-ACTCCTACGGGAGGCAGGAG-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)對樣品中細菌的保守區域進行擴增。使用真菌18srRNA通用引物ssu0817F(5′-TTAGCATGGAATAATRRAATAGGA-3′)和1196R(5′-TCTGGACCTGGTGAGTTTCC-3′)對樣品中真菌的保守區域進行擴增。用上海美吉生物公司的IlluminaMiSeq/NovaSeq平臺對PCR擴增產物進行雙端測序。過濾處理測序結果后得到優化序列,對序列進行去嵌合體處理,在97%的相似度水平上將序列進行OTU聚類,利用R語言分析不同階段的樣品在不同分類水平上的群落組成及相對豐度。
采用Chao、Shannon、Ace、Simpson指數進行Alpha多樣性分析,來反映樣本中微生物群落的豐富度和多樣性[8]。采用主成分分析(PCA)法解釋Beta多樣性指數,來反映樣品間物種群落構成的差異性。
1.2.4數據分析
將試驗數據導入Excel2007軟件進行統計,用SPSS19.0軟件進行單因素方差分析,用R語言繪圖;采用Spearman法分析堆肥的理化指標與微生物多樣性的相關性;利用KEGG數據庫[11]和MetaCyc數據庫[12]分別對堆肥過程中的細菌群落和真菌群落進行富集和功能注釋。
2結果與分析
2.1堆肥過程中核桃青皮理化指標的變化
2.1.1基本理化指標的變化
核桃青皮堆肥過程中基本理化指標變化如圖1所示。由圖1可以看出,堆肥3d后溫度高于50℃,堆肥4d后溫度達峰值62.3℃,50℃以上的溫度可以維持6d,持續高溫可以殺滅病原微生物,且能更好地腐熟肥料。堆肥的溫度也是堆體中微生物活性的體現,微生物的活動越劇烈,產生的熱量越大,堆肥的溫度峰值越高,高溫持續的時間越長。由圖1可以看出,堆肥的水分含量隨著時間的延長逐漸下降,是溫度升高導致了部分水分蒸發,且堆肥中微生物的活動也消耗了部分水分。由圖1可以看出,核桃青皮初始的pH值在中性偏酸范圍,但經堆肥后逐漸變為偏堿性,說明核桃青皮被降解后,其中的偏堿性物質被釋放出來。由圖1可以看出,隨著時間延長,堆肥的腐殖酸含量逐漸增加,在堆肥21d后達到10.38%。由圖1可以看出,隨著時間的延長,堆肥的EC值先升高后降低,在堆肥21d后為57S/m。
2.1.2養分含量的變化
核桃青皮堆肥過程中其養分含量變化情況如表1所示。由表1可以看出,全氮、全鉀、水解氮、有效磷、速效鉀、有機質的含量隨著時間的延長呈現升高趨勢,全磷含量逐漸降低。這可能與堆肥里面的微生物對磷的代謝有關,微生物將青皮中難溶解的磷元素降解成了易溶的離子形式,因此全磷含量降低而有效磷的含量增高。
2.2核桃青皮堆肥降解過程
掃描電鏡下不同階段核桃青皮的降解情況如圖2所示。隨著時間的延長,塊狀的青皮顆粒在微生物的作用下逐漸被降解為更為細小的顆粒,在堆肥21d后大部分塊狀青皮顆粒已經被降解,說明青皮的發酵較為成功。
2.3堆肥過程中核桃青皮微生物群落多樣性分析
2.3.1Alpha多樣性和Beta多樣性
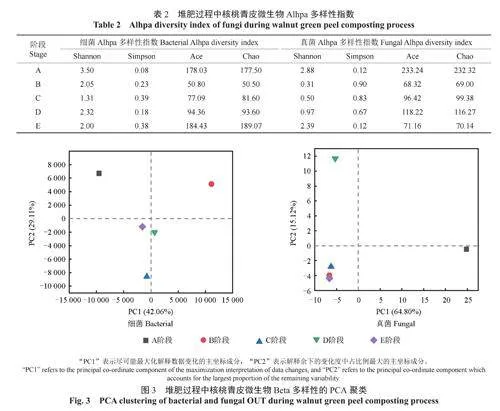
樣品的Alpha多樣性分析結果如表2所示。已知Shannon指數、Ace指數、Chao指數的值與物種多樣性為正相關關系,而Simpson指數的值與物種多樣性為負相關關系[13]。由表2可知:隨著堆肥過程的進行,堆體中細菌群落的Shannon指數、Ace指數、Chao指數均是先降低,在堆肥11d后達最低值,然后隨著時間延長逐漸增大;Simpson指數則是在堆肥11d后達最高值,然后隨著時間的延長而減小。由表2可知:隨著堆肥過程的進行,堆體的真菌群落的Shannon指數、Ace指數、Chao指數先降低后升高,在堆肥4d后達最低值,然后隨著時間延長逐漸增大;Simpson指數則是在堆肥4d后達最高值,然后隨著時間的延長而減小。由此可知,在堆體升溫過程中,隨著溫度的上升,微生物多樣性逐漸減少,說明溫度上升有助于殺滅大部分病原菌。在堆肥初期,嗜溫微生物通常是主導群落,群落多樣性較豐富,但隨著堆體溫度的升高,嗜溫微生物的活性受到嚴重影響,大部分不耐高溫的微生物死亡,群落結構發生了顯著變化,嗜熱微生物取代嗜溫微生物成為主導物種,導致堆體內物種多樣性降低。經歷過高溫后,隨著堆體溫度的下降,嗜熱微生物活動減弱,嗜溫微生物再次活躍起來,微生物多樣性和豐富度得到恢復。
在不同堆肥階段核桃青皮的細菌和真菌Beta多樣性的PCA聚類如圖3所示。由圖3可知:A階段由于是堆肥初期,其細菌多樣性單獨為一組;在B階段堆肥高溫期,堆肥細菌群落以嗜熱細菌為主,因此B階段也單獨為一組;C階段為群落恢復期,細菌多樣性單獨為一組;E階段和D階段由于溫度起伏較小,細菌多樣性更為接近,群落組成也更為相似。由圖3可知,A階段的真菌多樣性單獨為一組,D階段的真菌多樣性單獨為一組,B、C、E3個階段的真菌多樣性聚集在一起,形成一組。Beta多樣性分析結果表明,在不同的堆肥階段微生物群落多樣性不同,且堆肥初期與高溫期的微生物多樣性的差異較大,而溫度較為穩定階段的群落微生物的多樣性較為相似。
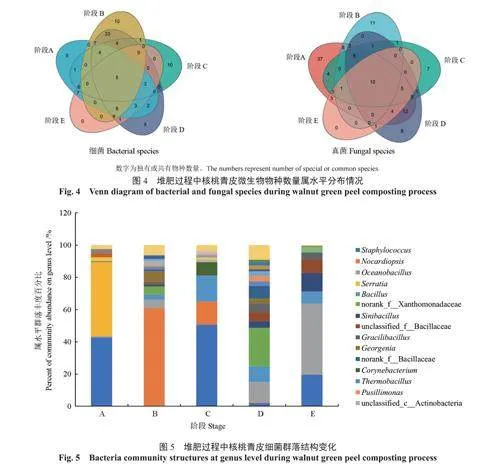
2.3.2各階段屬水平物種數量的比較
對堆肥不同階段的微生物在屬水平進行聚類,來反映不同階段核桃青皮堆肥的微生物群落物種數量的變化。細菌和真菌的聚類結果如圖4所示。由圖4可知,不同階段的細菌在屬水平上的物種數量有所不同,各階段細菌共有的屬有5個,其中芽孢桿菌屬Bacillus占23.74%,葡萄球菌屬Staphylococcus占74.90%,白細菌屬Leucobacter、短桿菌屬Brachybacterium和鏈霉菌屬Streptomyces3個屬共占1.37%。由圖4可知,不同階段的真菌在屬水平上的物種數量有所不同,各階段真菌共有的屬為10個,其中念珠菌屬Candida占40.04%,unclassified_f__Dipodascaceae占25.27%,青霉屬Penicillium占9.04%,曲霉屬Aspergillus占8.63%,Gibellulopsis占6.95%,地霉屬Geotrichum占5.59%,毛白毛孢霉菌屬Cutaneotrichosporon占1.96%,絲衣霉屬Byssochlamys占1.43%,節擔菌屬Wallemia和枝孢屬Cladosporium占1.09%。
2.3.3各階段細菌群落結構及其優勢菌群的變化
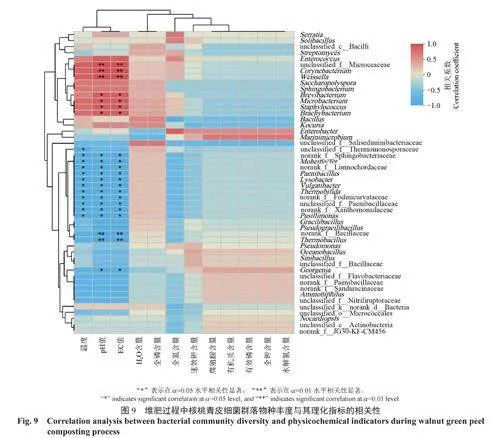
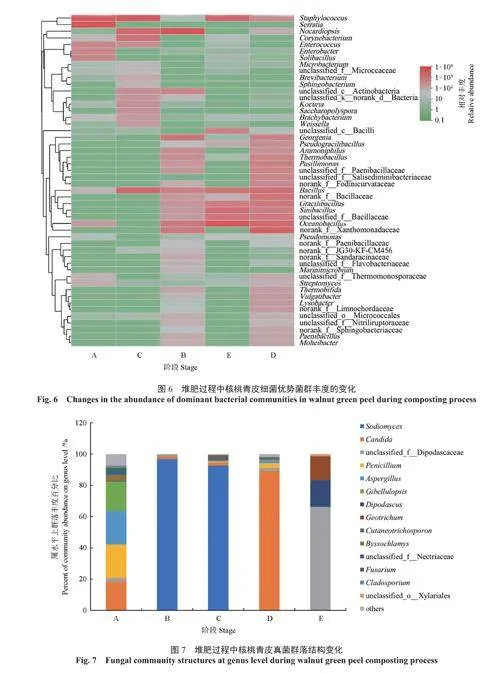
在堆肥不同階段,在屬水平上的核桃青皮細菌群落結構如圖5所示。由圖5可知:在A階段升溫期,優勢菌群為葡萄球菌屬(占42.76%)和沙雷氏菌屬Serratia(占45.83%);在B階段高溫期,優勢菌群為擬諾卡氏屬Nocardiopsis(占60.86%),已知擬諾卡氏屬能夠分解和降解大量的有機化合物,包括脂肪、碳水化合物和蛋白質等,因此此類菌可能在高溫期對核桃青皮中的有機物起到重要的降解作用[14];在C階段降溫期,優勢菌群為葡萄球菌屬(占50.55%)、擬諾卡氏屬(占14.59%)和黃單胞菌屬Xanthomonadaceae(占16.02%);在D階段溫度波動期,優勢菌群為黃單胞菌屬(占23.99%)、海洋芽孢桿菌屬Oceanobacillus(占12.73%)和芽孢桿菌屬Bacillus(占9.58%);在E階段穩定期,優勢菌群為海洋芽孢桿菌屬(占44.25%)、葡萄球菌屬(占19.54%)、華夏芽孢桿菌屬Sinibacillus(占11.46%)和芽孢桿菌屬(占7.48%)。經過一系列的溫度和理化性質變化后,最終芽孢桿菌成為了主要的核桃青皮堆肥菌群。芽孢桿菌是廣泛存在于糞便堆肥中的微生物,可以降解有機物,具有一定的木質纖維素降解活性[15-16]。
不同堆肥階段屬水平上的細菌群落優勢菌群豐度的變化如圖6所示,結果表明,A階段與C階段的細菌多樣性更為相似,B階段、D階段和E階段的細菌群落組成更加相似。
2.3.4各階段真菌群落結構及其優勢菌群的變化
在堆肥不同階段,在屬水平上核桃青皮真菌群落結構如圖7所示。由圖7可知,在堆肥前期,微生物群落數量較多,在屬水平上分布較為均勻,優勢菌群為念珠菌屬(占18.47%)、青霉屬(占21.46%)、曲霉菌屬(占21.39%)和Gibellulopsis(占18.62%),均是在環境中常見的菌群。在B階段溫度升高后,群落數量下降,優勢菌群為Sodiomyces(96.83%);在C階段,隨著溫度降低,真菌群落數量又逐漸增加,但優勢菌群仍是Sodiomyces(92.63%);D階段屬于堆肥后期,但是堆肥溫度仍有一定起伏,優勢菌群轉化為念珠菌屬(89.21%);E階段為堆肥完成期,其優勢菌群轉變成unclassified_f__Dipodascaceae(65.82%)、雙足囊菌屬Dipodascus(17.09%)和Sodiomyces(15.03%)。
不同堆肥階段屬水平上的真菌群落優勢菌群豐度的變化如圖8所示,結果表明A階段與D階段、B階段與C階段的優勢屬組成更加相似,這4個階段與E階段堆肥優勢屬組成存在差異。
2.3.5各階段微生物代謝功能的變化
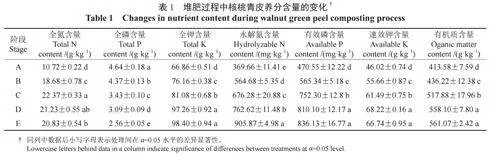
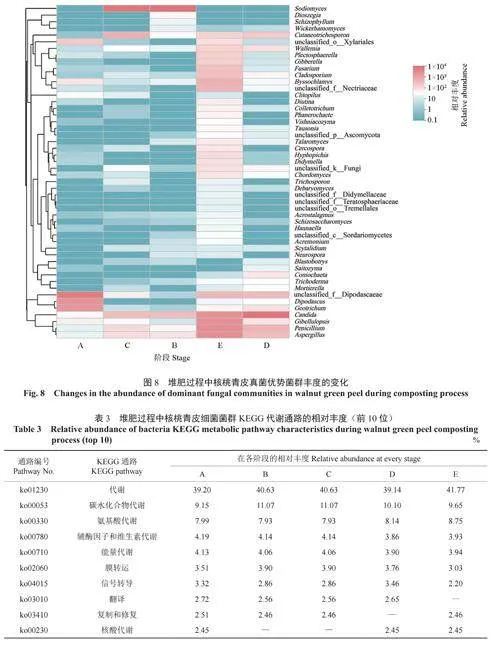
微生物群落多樣性能夠在一定程度上揭示微生物群落的演替規律,但無法解釋堆肥過程中菌群發揮的功能。為了明確微生物群落的代謝功能,將檢測到的微生物序列與相關的代謝數據庫進行比對,并對微生物群落的相關代謝通路進行分析。各階段核桃青皮細菌群落相對豐度排前10位的KEGG代謝通路見表3。由表3可知,在整個堆肥過程中,細菌菌群的代謝通路顯示出較高的豐富性。其中,菌群的代謝過程主要體現在碳水化合物代謝、氨基酸代謝、輔酶因子和維生素代謝、能量代謝及核酸代謝等方面,細菌群落的膜轉運、信號轉導、核酸的翻譯及復制和修復過程在5個階段中的相對豐度也較高,說明在堆肥過程中細菌菌群的生命活動活躍,對核桃青皮中的物質進行了重復的利用和降解。
各階段核桃青皮真菌群落相對豐度排前10位的MetaCyc代謝通路見表4。由表4可知,真菌菌群在5個階段中呼吸作用(Ⅰ和Ⅱ)的相對豐度最大,說明在堆肥過程中,真菌充分吸收了氧氣,生命活動活躍,對青皮堆肥進行了有氧發酵。良好的通氣性有助于維持微生物的活動和代謝,促進有益菌群的繁殖,提高分解有機物的效率。5個階段的真菌群落在脂肪酸代謝相關通路上的相對豐度也較高,例如脂肪酸β-氧化的通路和TCA循環Ⅱ通路對生物體有效地將脂肪酸轉化為能量具有重要作用。在B、C、D、E4個階段中,維持真菌生命活動相關的通路有甘酰乙酸循環、GDP-甘露糖生物合成途徑、D-肌肌醇(1,4,5)-三磷酸生物合成通路、tRNA激活途徑、腺苷核苷酸的新合成途徑和丙酮酸轉異丁醇途徑等,與群落生命活動相關途徑的相對豐度較高,可能是堆肥真菌群落在生存和繁殖過程中活躍的生命活動所導致。
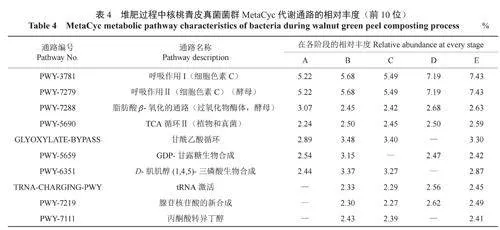
2.4堆肥過程中核桃青皮理化指標與微生物豐度的相關性
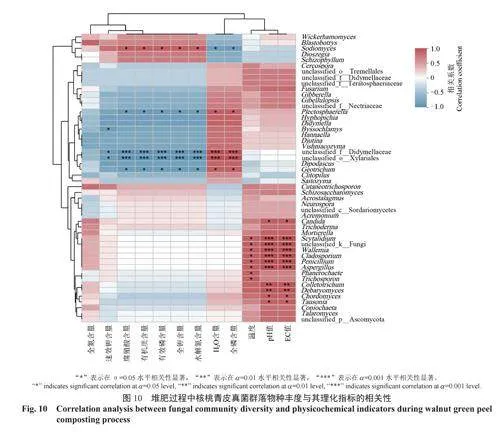
將堆肥過程中核桃青皮的溫度、pH值、全鉀含量、水分含量、EC值、全氮含量、全磷含量、有機質含量、腐殖酸含量、速效鉀含量、水解氮含量、有效磷含量與細菌屬水平物種豐度進行相關性分析,結果如圖9所示。結果表明:堆肥的溫度與unclassified_f__Thermomonosporaceae、norank_f__Sphingobacteriaceae、Moheibacter等12個屬的豐度顯著負相關;pH值與細菌屬水平的相關性和EC值與細菌屬水平的相關性趨勢表現一致,即與unclassified_o__Micrococcales、棒狀桿菌屬Corynebacterium、Weissella的豐度極顯著正相關,與微桿菌屬Microbacterium、短桿菌屬Brevibacterium、葡萄球菌屬、短狀桿菌屬的豐度顯著正相關,與norank_f__Sphingobacteriaceae、Moheibacter、norank_f__Limnochordaceae、類芽孢桿菌Paenibacillus等12個屬的豐度顯著負相關,與norank_f__Bacillaceae和耐熱芽孢桿菌屬Thermobacillus的豐度極顯著負相關;堆肥的其余指標與細菌屬水平上物種豐度無顯著相關關系。
將堆肥過程中核桃青皮的溫度、pH值、全鉀含量、水分含量、EC值、全氮含量、全磷含量、有機質含量、腐殖酸含量、速效鉀含量、水解氮含量、有效磷含量與真菌屬水平物種豐度進行相關性分析,結果如圖10所示。結果表明:堆肥的速效鉀含量與絲衣霉屬、雙足囊菌屬、unclassified_o__Xylariales的豐度顯著負相關;堆肥的腐殖酸含量、有機質含量、有效磷含量、全鉀含量、全氮含量與芽生葡萄孢酵母屬Blastobotrys的豐度顯著正相關,與小不整球殼屬Plectosphaerella和地霉屬的豐度顯著負相關,與unclassified_f__Dipodascaceae、unclassified_o__Xylariales的豐度極顯著負相關;堆肥的水分含量、全磷含量與芽生葡萄孢酵母屬的豐度顯著負相關,與小不整球殼屬和地霉屬的豐度顯著正相關,與unclassified_f__Dipodascaceae、unclassified_o__Xylariales的豐度極顯著正相關;堆肥的溫度與柱霉屬Scytalidium、節擔菌屬、unclassified_k__Fungi等8個屬的豐度顯著正相關;堆肥的pH值和EC值與念珠菌屬、Chordomyces、Tausonia的豐度顯著正相關,與柱霉屬、unclassified_k__Fungi、節擔菌屬等5個屬的豐度極顯著正相關。
3討論與結論
已知核桃青皮中含有較多的多酚類物質,多酚類物質具有一定的抗菌活性[17],僅用核桃青皮作為主要原料發酵時微生物的生命活動可能會被其抑制,且有報道用鮮核桃青皮堆肥時溫度僅能達到30~40℃[18],可能無法達到物料腐熟溫度,所以較多試驗中將核桃青皮與其他輔料進行配比堆肥[4-5]。本研究中,通過對干燥后的核桃青皮進行堆肥處理,證明了僅以核桃青皮為主要原材料進行堆肥,在堆肥4d后溫度可達60℃以上,且50℃以上的溫度可以維持6d,達到了殺滅多數病原菌的溫度要求。理化指標測定結果表明,在堆肥過程中,核桃青皮的全氮和全磷的含量呈現先升高后降低的趨勢,可能是由于微生物的活動消耗了部分氮元素和磷元素,而其全鉀、水解氮、有效磷、速效鉀、腐殖酸、有機質等的含量逐漸升高,說明堆肥中微生物的活動既消耗了一定量的養分,又轉化了青皮中的部分養分,將難溶的物質降解轉化成了生物體易吸收的形式。掃描電鏡下觀察結果表明,在微生物的降解作用下,大顆粒的青皮被降解成了小顆粒的物質。核桃青皮堆肥的養分測定結果顯示,其氮含量、磷含量、鉀含量、有機質含量、腐殖酸含量等指標均符合文獻[19]中的相關要求,且其中鉀含量較高,與之前的報道一致[4],因此,核桃青皮可被單獨作為原料用來生產有機肥。
堆肥過程一般被劃分為4個時期(嗜溫期、嗜熱期、降溫期和腐熟期),4個時期優勢菌群的演替與堆肥發酵效率和堆肥質量密切相關[18]。在核桃青皮堆肥過程中,由于不同階段其溫度的變化,細菌和真菌的群落多樣性均呈現先降低后升高的趨勢。在核桃青皮堆肥的嗜溫期,主要的細菌群落為葡萄球菌屬和沙雷氏菌屬,真菌優勢菌群為念珠菌屬、青霉屬、曲霉菌屬和Gibellulopsis;在嗜熱期,細菌優勢菌群為擬諾卡氏屬,真菌的優勢菌群為Sodiomyces;在降溫期,細菌的優勢菌群為葡萄球菌屬、擬諾卡氏屬和黃單胞菌屬,真菌的優勢菌群為Sodiomyces和念珠菌屬;在腐熟期,細菌的優勢菌群為海洋芽孢桿菌屬Oceanobacillus、葡萄球菌屬、華夏芽孢桿菌屬和芽孢桿菌屬,真菌的優勢菌群為unclassified_f__Dipodascaceae、雙足囊菌屬和Sodiomyces。核桃青皮中的化合物影響了堆肥的pH值,使其偏堿性,部分菌群由于能夠在偏堿的環境下生存繁殖,得以發展為優勢菌群,這些菌群能夠在高溫堿性環境下生存繁殖并且對青皮起到一定的降解作用。
通過對微生物進行KEGG和MetaCyc功能分析,結果表明微生物在代謝過程和生命活動相關途徑上相對豐度較高,說明微生物通過活躍的代謝活動對核桃青皮進行了有效降解[20]。
堆肥的養分指標與微生物群落的相關性分析結果表明,溫度、pH值和EC值對微生物屬水平上物種的相對豐度影響較大,真菌屬水平上物種的相對豐度與堆肥的養分指標(全磷含量、全鉀含量、速效鉀含量、有效磷含量、水解氮含量、有機質含量)有一定的相關性,而細菌屬水平上物種的相對豐度與堆肥的養分指標的相關性不強。目前,相關研究主要聚焦在農業廢棄物還田對土壤養分與微生物群落的影響[21],有關堆肥過程中微生物群落與養分變化的關系的研究報道較為鮮見,本研究結果可以為該領域提供一定的思路。
本研究中對核桃青皮堆肥的優勢菌群進行了分析,研究結果可為明確微生物的降解功能特性提供參考,可為解決核桃青皮等固體廢物的資源化處理問題提供思路。為給高效降解核桃青皮微生物菌劑的研制提供參考,下一步將研究分離各堆肥階段的優勢菌種。
