延長鹽單胞菌生產依克多因高密度發酵工藝優化
2025-02-26 00:00:00張悅劉昊劉瑋余君雄肖揚郇晶偉顏誠旭莊英萍王澤建
華東理工大學學報(自然科學版) 2025年1期


關鍵詞:延長鹽單胞菌;合成培養基;高細胞密度培養;依克多因;鹽沖擊
依克多因(Ectoine,1,4,5,6-四氫-2-甲基-4-羧酸)為天冬氨酸的衍生物,是嗜鹽菌在極端環境中合成的產物。依克多因可以提高膜脂表面的水合作用力,維持細胞新陳代謝所需的胞質空間和正常細胞形態[1-5],除此以外該物質在皮膚細胞的保護和穩定方面具有良好功效[6]。目前,依克多因作為一種高價值的化合物被廣泛應用在化妝品、醫藥和食品等領域,它可以通過化學合成和生物發酵的方法得到,前體的高成本及合成過程中可能帶來的環境污染使得化學方法在工業化生產中不具優勢[7]。因此,生物發酵生產依克多因已成為主要生產方式,其中延長鹽單胞菌(Halomonaselongata)是主要的工業生產菌株[8-10]。延長鹽單胞菌H.elongataDSM2581T屬于中度嗜鹽菌,能夠在高鹽環境下快速生長,原因在于胞內積累大量的依克多因來維持細胞內外滲透壓平衡[11-13]。
許多研究致力于優化生物合成的方法,設計了許多提高依克多因產量的方法,如“細菌擠奶”工藝[14]。同時,代謝工程也為提高依克多因產量作出了貢獻,如Ning等[15]構建了在低鹽濃度下高效生產依克多因的大腸桿菌菌株。基因操控下的羥化酶的缺失,也會減少依克多因的羥基化而提高依克多因的產量[16]。如何有效地控制微生物的生長和代謝產物的生成,提高發酵生產水平是生產下游環節的重點。發酵過程的優化依然是突破生產瓶頸的關鍵,Salman[17]和Chen等[18]都通過優化培養條件成功地提高了淀粉酶和依克多因的生產水平。利用能夠精準調控合成培養基,高效快速地生產質量穩定的依克多因,是實現依克多因低成本生產的途徑之一。
依克多因的詳細生物合成機制迄今還未完全闡明,研究其中的合成機制及調節關鍵節點具有巨大的商業價值。高細胞密度發酵(HighCellDensityFermentation,HCDF)在工業生產上是用來增加產量的常用方法[19]。利用合成培養基實現細胞的高細胞密度發酵,兼顧高鹽濃度培養基可以刺激依克多因生物合成的特點[20-21],建立H.elongataDSM2581T的高密度培養有效提升依克多因產量的新工藝,為其工業化規模的低成本生產奠定理論基礎。
本研究考察并優化了影響H.elongataDSM2581T依克多因發酵的全合成培養基的營養成分及添加濃度,并在5L發酵罐水平考察了H.elongata在全合成培養基中的發酵代謝情況;基于優化后的合成培養基協同鹽脅迫工藝對延長鹽單胞菌進行高細胞密度培養,最終獲得高于已報道的天然微生物生產依克多因的產量,并且獲得高生產速率的發酵效果。
1實驗部分
1.1原料和試劑
1.1.1出發菌株 實驗菌種延長鹽單胞菌H.DSM2581T,屬于中度嗜鹽菌和革蘭氏陰性細菌,由中國科學院微生物所提供。
1.1.2培養基與培養條件
(1)菌種活化培養基(g/L):蛋白胨10.00、酵母粉5.00、氯化鈉80.00、瓊脂16.00,pH7.00±0.10。
(2)種子培養基(g/L):酵母粉5.00、蛋白胨10.00、氯化鈉80.00、pH7.00±0.10。
(3)M63培養基按照Fatollahi等[22]描述的方法配制;PTM1微量元素溶液按照Charoenrat等[23]等描述的方法配制。
1.2測試與表征
1.2.1實驗儀器 SBA-40E型生物傳感分析儀(山東省科學院生物研究所);pH計(梅特勒-托利多);GZX-9420MBE型鼓風烘箱(華連醫療器械);5L生物反應器(上海國強生化工程裝備有限公司);LC-2010A型高效液相色譜(日本島津公司);YXQ-LB-100SII型壓力蒸汽滅菌器(BOXUN);FA1004型電子天平(上海良平儀器分析儀表有限公司);ZWYC-2933型智城恒溫培養振蕩器(上海智城分析儀器制造有限公司);VARIOSKANLUX型酶標儀(賽默飛世爾科技公司)。
1.2.2檢測方法
(1)生物量檢測方法:利用酶標儀在低振蕩、600nm波長下測量吸光度值(OD600);菌體干重(DryCellWeight,DCW):發酵液離心棄上清,取沉淀于90℃烘箱中烘干至恒重,稱重,將細胞干重與菌液吸光度值作線性擬合,得DCW=0.425×OD600。
(2)葡萄糖濃度檢測方法:發酵液離心取上清進行適當稀釋,經0.22μm水系膜過濾后由生物分析傳感器測量。
(3)依克多因含量檢測方法:取發酵液經超純水稀釋3倍后,加入玻璃研磨珠在冷凍研磨機中于65Hz條件下研磨30min。取出后經過12000r/min轉速下離心5min去沉淀,上清經0.22μm水系膜過濾保存至液相小瓶,待測。依克多因通過島津HPLC檢測,使用SB-C18色譜柱(250mm×4.6mm,5μm,Agilent公司),流動相為水∶乙腈(體積比為98∶2),恒定流速0.5mL/min,柱溫30℃,檢測器為SPD-20AV,紫外檢測波長210nm[24]。
1.2.3動力學計算 菌體生長動力學可以將復雜的細胞代謝過程用簡明的方式展現出來[25-26],本文采用式(1)擬合H.elongata的生長動力學。
其中,X、X0和Xm分別代表菌體質量濃度、初始菌體質量濃度和最大菌體質量濃度(g/L),μm代表最大比生長率(h?1),t代表發酵時間(h)。
1.2.4統計分析 采用DesignExpert8.06軟件設計Plackett-Burman實驗,CentralCompositeDesign采用Origin2021軟件進行數據分析及繪圖,統計學分析采用SPSS的ANOVATuckypost-hoc檢驗。
1.3實驗步驟
1.3.1搖瓶培養 將種子液按10%的接種量接入裝有45mL培養基的250mL擋板搖瓶中,于37℃、225r/min培養12h。
1.3.2反應器培養 實驗采用5L體系發酵罐,裝液量為3L。高壓濕熱滅菌后按10%的接種量進行接種。通氣量為1vvm,轉速為600r/min,罐壓控制為0.05MPa,溫度為37℃。發酵過程每間隔1h取樣,檢測生物量、依克多因含量和殘糖濃度等參數。
2結果與討論
2.1合成培養基優選
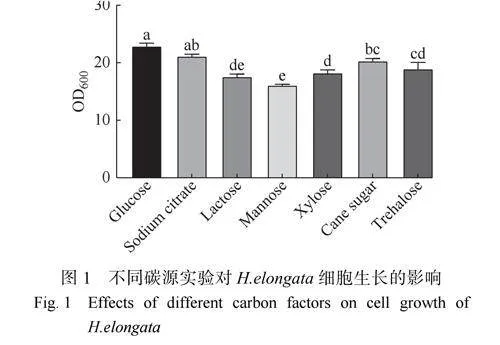
2.1.1碳源篩選實驗 碳源是細菌生長和代謝過程中的重要物質基礎,同時也是生物合成、維持生命活動的能量來源[27]。不同的碳源對微生物生長代謝的影響不同,易利用的碳源可以促進微生物的生長和代謝。本研究考察了不同碳源種類(葡萄糖、檸檬酸鈉、乳糖、甘露糖、木糖、蔗糖、海藻糖)對H.elongataDSM2581T生長的影響,利用多工位反應器進行碳源篩選實驗,培養20h后測定生物量,結果如圖1所示。在不同碳源下延長鹽單胞菌H.elongata均可以生長,說明H.elongata具有廣泛的碳源底物譜,胞內存在利用多種碳源的酶系。在以葡萄糖為碳源的培養基中獲得的生物量最多,其次是檸檬酸鈉,可見兩種物質都是有利于細胞生長的碳源。而甘露糖、乳糖為碳源的培養基中生物量明顯低于其他組別。
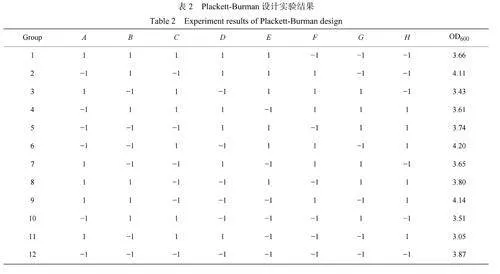
2.1.2Plackett-Burman實驗設計與結果分析 有研究[28]發現在較高鹽濃度(gt;10g/L)下H.elongataDSM2581T的產物合成為生長相關型。基于此發現,在優選營養成分過程中,當Plackett-Burman實驗設計時選擇以生物量為指標,在基礎培養基(葡萄糖10g/L;PTM11mL/L;氯化鈉60g/L)上利用Design-Expert8.06Trial軟件包統計學工具箱實施n=12的Plackett-Burman實驗設計,考察了合成培養基中的檸檬酸鈉、氯化銨、磷酸氫二鈉、硫酸鈉、氯化鉀、硫酸鎂、硫酸鋅、硫酸錳8個成分對延長鹽單胞菌生長的影響。利用250mL搖瓶培養10h后測定OD600,實驗設計因素水平及編碼見表1,實驗結果見表2,評價指標為生物量。每組設置3組平行。
經軟件分析得到各培養基成分對生物量(Y)影響的擬合方程,Y=3.73?0.091A+0.21B+0.085C?0.041D+0.088E?0.090F?0.13G?0.035H。因為p值為0.0212(lt;0.05),表明對于生物量評分所建立的回歸方程顯著。氯化銨的p值0.0046lt;0.01,表明氯化銨是影響生物量的極顯著因素。研究指出氮源對提高生物量和依克多因生產率具有重要意義[20]。檸檬酸鈉、硫酸錳、硫酸鋅、硫酸鎂的p值小于0.05,表明這些物質也是影響細胞生長的顯著因素。Plackett-Burma實驗結果統計分析后確定了以上幾種培養基成分的最佳添加質量濃度為:硫酸鈉2.50g/L,七水合硫酸鎂3.00g/L,硫酸鋅0.25g/L,硫酸錳0.09g/L,十二水磷酸氫二鈉7.50g/L。
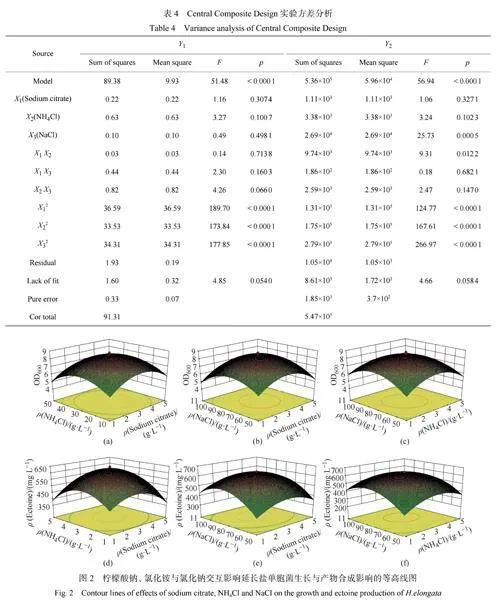
2.1.3CentralCompositeDesign實驗設計與結果分析
研究表明鹽濃度是影響依克多因合成的重要因素[29-30]。由本實驗篩選出的影響延長鹽單胞菌生長顯著的因素:檸檬酸鈉(X1)和氯化銨(X2),將其與促進依克多因合成的主要因素氯化鈉(X3)相結合。通過中心組合實驗考察這3種因素對延長鹽單胞菌生長(生物量Y1)和依克多因合成(依克多因產量Y2)的影響。實驗設置3組平行,各因素及水平的實驗設計見表3,分析結果見表4。為了預測最佳點,運用DesignExpert8.06軟件對試驗數據進行方差分析和二次多項回歸擬合,得到以生長和產物合成為指標的多元二次回歸方程:Y1=8.48+0.13X1?0.21X2?0.084X3?0.059X1X2+0.24X2X3?0.32X2X3?1.59X12?1.53X22?1.54X32;Y2=620.99+9.02X1+15.74X2+44.39X3+34.89X1X2?4.82X1X3?17.98X2X3?95.16X12?110.30X22?139.20X32。以Y1為響應值的模型決定系數R2=0.9789,校正決定系數R2Adj=0.9599,plt;0.0001,模型高度顯著。以Y2為響應值的模型決定系數R2=0.9809,校正決定系數R2Adj=0.9636,plt;0.0001,模型高度顯著。X12、X22、X32的p均小于0.0001,說明因素X1、X2、X3對生物量和依克多因產量的影響都極顯著,因素X3對依克多因產量影響極顯著,X1X2兩兩相交效應對依克多因產量影響顯著。
各因素的響應面等值線圖如圖2所示,3個響應面的等高線中心都在設定的范圍內,表明在設計的因素水平內存在最優設計條件。由圖2可知,三維響應曲面呈弧形,檸檬酸鈉、氯化銨、氯化鈉呈兩兩間交互作用,分別固定檸檬酸鈉、氯化銨、氯化鈉,生物量和依克多因產量隨著另外兩個因素取值的增大先升高后降低,曲面頂點即為生物量和依克多因產量最高點及其對應的最優因素添加質量濃度。當氯化鈉處于中心水平時,檸檬酸鈉和氯化銨對依克多因產量影響較大,響應面坡度較陡。
由以上實驗得到的最優化添加質量濃度為:檸檬酸鈉3g/L,氯化銨3g/L,氯化鈉80g/L。經過搖瓶實驗驗證,在最優條件下培養基中得到的生物量OD600和依克多因產量分別為8.48和618.70mg/L,與預測值一致,證明該模型較為合理。
2.2延長鹽單胞菌產依克多因的發酵工藝優化
2.2.1批發酵工藝 在5L罐的發酵中,對優化后的合成培養基(HE9)進行延長鹽單胞菌的發酵過程代謝參數考察,結果如圖3所示。在添加葡萄糖20g/L的條件下菌體OD600最高可達到26.60,依克多因產量為2.00g/L,對比搖瓶發酵結果均有明顯的提高。利用Logistic方程擬合菌體的生長動力學模型,結果表明在優化的全合成培養基HE9中延長鹽單胞菌的μm可達到0.49h?1。有研究表明H.elongataDSM2581T在鹽質量濃度為80g/L的復合培養基條件下μm為0.65h?1[28],約為HE9培養基所達到μm的1.3倍。
從整個發酵過程來看,菌體生長經歷短暫的延滯期后進入菌體快速生長的指數期,依克多因進入快速合成階段與菌體生長的對數生長期重合,這與之前報道的H.elongataDSM2581T在氯化鈉質量濃度為80g/L時依克多因合成是一個高度耦合生長過程的結果一致[28]。碳源的變化情況表現為:發酵前5h碳源消耗較慢,僅僅消耗了17.50%的葡萄糖;隨著細胞生長進入對數生長期,葡萄糖消耗速度顯著增加,pH開始快速下降,通過氨水調節pH值在6.5左右至發酵結束。CO2釋放率(Carbon-dioxideEscapeRate,CER)總體呈現與菌體生長的一致趨勢。隨著分批培養葡萄糖的耗盡,菌體生長和依克多因的合成出現停滯。
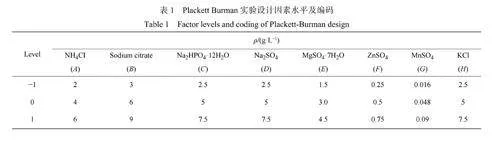
2.2.2分批補料發酵 為了進一步提升菌體的生物量和依克多因的合成,在優化的培養基基礎上研究了連續流加葡萄糖的延長鹽單胞菌高密度培養工藝。發酵培養7h后流加葡萄糖溶液,利用發酵液殘糖濃度反饋調整葡萄糖溶液流加速率;利用氨水進行pH調控和氮元素補充,發酵過程各曲線如圖4所示。發酵培養34h時,OD600最高可達到191,依克多因產量達到11.05g/L(圖4(a)),菌體氧攝取速率(OxygenUptakeRate,OUR)最高達到220mmol/(L·h)(圖4(b))。與批式發酵相比,最高OD600和依克多因產量分別為批發酵結果的7.18倍和5.53倍。發酵中期觀察到菌體生長緩慢、OUR持平的現象,在發酵20h時補充2g/L的磷酸氫二鉀,隨后菌體生長速率和OUR增加速率明顯加快。磷酸鹽含量顯著影響工程大腸桿菌和嗜鹽生物的生物量和依克多因產量[18,31],說明這段生長的緩慢期可能是由于HE9培養基中磷元素不足導致細胞生長受限。對不同發酵時期的生理代謝參數(表5)進行分析發現,發酵后期依克多因產率明顯下降的現象(圖4(c)),遠低于理論最大產率[32]。補料過程中持續葡萄糖和氨水的補入會導致鹽濃度的逐步下降,從而影響依克多因的合成,這可能是導致發酵后期比產率低的重要原因。
2.2.3鹽梯度脅迫促進依克多因合成的優化控制工藝 為改善高密度培養發酵后期出現的菌體生長緩慢和依克多因轉化率低的問題,進一步對HE9合成培養基中磷源濃度進行優化,并與鹽梯度沖擊控制相結合,以達到促進依克多因的快速生成。在發酵過程的指數期(8h)進行鹽梯度沖擊實驗,結果如圖5、表6所示。在指數生長中期利用一次性補加的方式將發酵液中氯化鈉的質量濃度從80g/L補充到130g/L(圖5(a))。OUR在鹽沖擊后有短暫的下降,隨后維持著快速增加的趨勢,最高達到209.84mmol/(L·h)(圖5(b))。發酵13h后OD600達到169.3,依克多因產量為20g/L。與優化前的工藝相比,依克多因產量提高了81%。依克多因合成速率在進行鹽梯度沖擊(8h)后顯著增加(圖5(d)),最大達到了5.5g/(L·h)。圖5(c)所示的依克多因比產率的結果也表明在鹽梯度沖擊的過程中,發酵10h時的比產率(0.086g/(g·h))顯著高于分批補料發酵最高的比產率(0.016g/(g·h)),說明鹽梯度沖擊能夠有效地促進依克多因的合成,而且優化后發酵時間顯著縮短。
本研究中發現在搖瓶中生產依克多因的最佳鹽質量濃度為80g/L。有研究利用動力學模型對不同鹽濃度下H.elongataDSM2581T菌體生長和依克多因生物合成進行擬合發現:當氯化鈉質量濃度低于80g/L時,H.elongataDSM2581T的依克多因的生物合成速率隨著氯化鈉質量濃度的增加而增加[33]。但過高的鹽濃度超過了細胞的耐受水平,反而抑制細胞生長[28,34]。因此本研究利用不同鹽濃度的培養基分階段進行依克多因生物合成:在前期,80g/L的鹽質量濃度可以給細胞提供良好的生長環境,當細胞生長到一定階段采取提高鹽質量濃度的方式進入刺激依克多因合成階段。
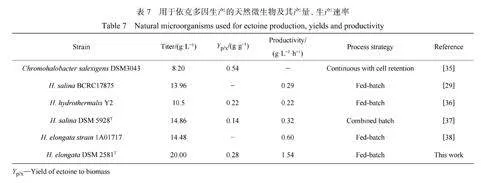
在合成培養基上結合連續補料,鹽梯度沖擊工藝進行高密度細胞密度發酵,可以獲得OD600為169.3,依克多因產量為20g/L的結果。與目前已報道的嗜鹽菌底盤的依克多因生產結果(表7)相比,本研究優化后的依克多因的產量和生產速率與之相比具有極大的優勢。發酵產量達到了20g/L,比其他嗜鹽菌的最高發酵水平(14.86g/L)[37]提高了34.5%。尤其是依克多因的合成速率達到了1.54g/(L·h),遠高于文獻中的報道水平。
3結論
本研究首先通過單因素實驗研究了不同碳源對H.elongataDSM2581T菌體生長的影響,結果顯示葡萄糖是最佳碳源。通過Plackett-Burman考察了培養基中各組分的濃度對生物量的影響,結果表明顯著因素為檸檬酸鈉和氯化銨。利用CentralCompositeDesign進一步優化了培養基,最終確定檸檬酸鈉3g/L,氯化銨3g/L,氯化鈉80g/L。基于優化后的全合成培養基,進行連續補料和指數中期鹽梯度式沖擊的發酵工藝研究,最終獲得的細胞量的OD600約為169.3,依克多因產量和依克多因合成速率分別達到20g/L和1.54g/(L·h)。這說明在該合成培養基上聯合連續補料和指數期鹽梯度沖擊進行高密度培養的可行性,為高效合成依克多因的發酵工藝提供了數據支撐。
