小麥新品系配合力和遺傳力分析
2009-01-15 09:03:36于海濤閆美徐磊李安飛
山東農(nóng)業(yè)科學(xué) 2009年12期
于海濤 閆 美 徐 磊 李安飛
摘 要:采用8×6不完全雙列雜交,對8個小麥新品系的9個性狀進(jìn)行配合力和遺傳力分析。結(jié)果表明,株高、穗長和穗下節(jié)間長以加性效應(yīng)為主,而結(jié)實小穗數(shù)、總小穗數(shù)、穗粒數(shù)、單株穗數(shù)、單株粒重和百粒重以非加性效應(yīng)為主。遺傳力分析表明,株高、穗長和穗下節(jié)間長廣義遺傳力和狹義遺傳力均較大,可在早代進(jìn)行選擇,其它性狀早代選擇效果不佳。
關(guān)鍵詞:小麥; 品種(系); 配合力; 遺傳力
中圖分類號:S512.103.2 文獻(xiàn)標(biāo)識號:A 文章編號:1001-4942(2009)12-0024-05Analysis
小麥?zhǔn)俏覈牡诙蠹Z食作物,選育新品種(系)提高小麥產(chǎn)量對我國農(nóng)業(yè)的發(fā)展具有重大的作用[1]。配合力和遺傳力是雜交育種和雜種優(yōu)勢利用中親本選配的重要指標(biāo),對于評價親本、正確選擇和提高育種效率都具有重要的指導(dǎo)意義。前人在小麥配合力研究中已經(jīng)做了大量的工作,對早熟、粒重[2,3]、沉淀值[4]、冠層溫度[5]、穗部性狀[6]和產(chǎn)量性狀[7~10]等均進(jìn)行了分析,為深入了解各個性狀的遺傳規(guī)律和指導(dǎo)育種工作積累了豐富的資料。本試驗選用了山東農(nóng)業(yè)大學(xué)農(nóng)學(xué)院新選育的8個新品系進(jìn)行配合力和遺傳力分析,以了解其配合力和遺傳力,明確其作為親本在育種中的價值。
1 材料與方法
1.1 試驗材料與設(shè)計
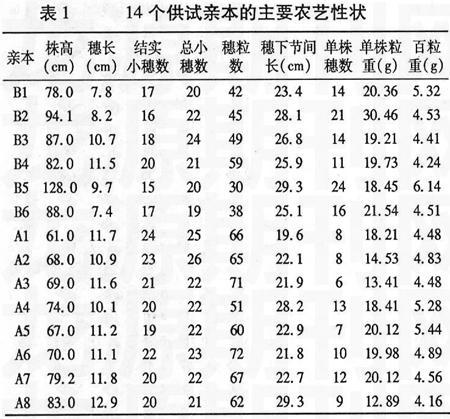
試驗于2006~2007年在山東農(nóng)業(yè)大學(xué)農(nóng)學(xué)實驗站進(jìn)行。以山東農(nóng)業(yè)大學(xué)選育的530001(A1)、530065(A2)、530170(A3)、530311(A4)、530319(A5)、530351(A6)、530385(A7)、530639(A8)共8個矮稈、抗病、大穗、早熟種質(zhì)系為母本組,以濟(jì)麥20(B1)、藁城8901(B2)、山農(nóng)919(B3)、山農(nóng)5536(B4)、輝縣紅(B5)、山農(nóng)990525(B6)6個品種(系)為父本組,組配8×6不完全雙列雜交。供試材料的農(nóng)藝性狀情況見表1。試驗采用隨機(jī)區(qū)組設(shè)計,重復(fù)2次,每組合F1和親本種植2行,行長1.55 m,行距25 cm,粒距6.7 cm,人工點播。栽培管理措施按常規(guī)進(jìn)行。
1.2 性狀調(diào)查
于成熟前各組合隨機(jī)取20個單莖,調(diào)查株高、穗長、結(jié)實小穗數(shù)、穗粒數(shù)、總小穗數(shù)、穗下節(jié)間長。完熟時每組合收獲10個單株考察單株穗數(shù)、單株粒重并計算百粒重。
1.3 統(tǒng)計分析
采用劉來福的不完全雙列雜交分析方法進(jìn)行配合力模型分析[11],使用Excel和DPS軟件進(jìn)行方差分析[12]。
2 結(jié)果與分析
2.1 各性狀配合力的方差分析
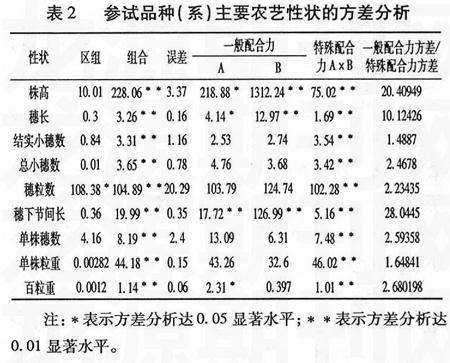
對不完全雙列雜交組合的9個性狀進(jìn)行方差分析,結(jié)果表明:供試材料株高、穗長、結(jié)實小穗數(shù)、總小穗數(shù)、穗粒數(shù)、穗下節(jié)間長、單株穗數(shù)、單株粒重、百粒重9個性狀在F1組合間存在極顯著差異(表2)。因此可以對這9個性狀進(jìn)行配合力分析。
由表2可知,株高、穗長、穗下節(jié)間長和百粒重4個性狀中除父本百粒重外其它的一般配合力差異達(dá)到極顯著或顯著水平,同時它們的特殊配合力差異均達(dá)到極顯著水平,表明這4個農(nóng)藝性狀的遺傳同時受基因加性和非加性效應(yīng)的共同作用。結(jié)實小穗數(shù)、總小穗數(shù)、穗粒數(shù)、單株穗數(shù)和單株粒重的一般配合力差異并不顯著,而特殊配合力差異達(dá)到極顯著,表明這5個性狀主要由非加性效應(yīng)作用。由一般配合力和特殊配合力的方差比可以看出,株高、穗長、穗下節(jié)間長的一般配合力方差大幅度超過特殊配合力方差,而其余6個性狀的一般配合力方差略大于特殊配合力方差。表明株高、穗長和穗下節(jié)間長3個性狀的遺傳方差中,加性效應(yīng)占主導(dǎo)地位;其余6個性狀的的遺傳方差中加性和非加性效應(yīng)同等重要。
2.2 一般配合力效應(yīng)
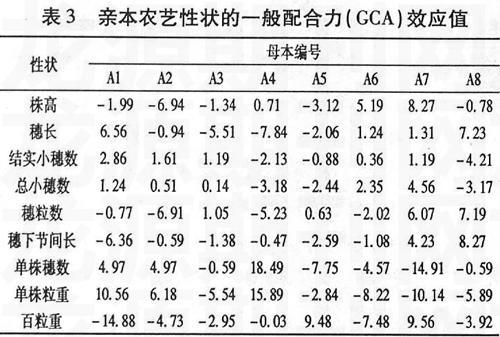
一般配合力是對基因加性效應(yīng)的度量,某個親本某個性狀一般配合力效應(yīng)值大,表明親本的加性基因效應(yīng)高,向后代傳遞能力強(qiáng),易于穩(wěn)定遺傳和固定選取一般配合力高的親本,可望該親本在所配制的雜交組合中對該性狀產(chǎn)生較大的影響。從表3中可以看出,同一親本的不同性狀或同一性狀的不同親本的一般配合力效應(yīng)值差別很大。就株高而言,母本組8個親本中A1、A2、A3、A5、A8都有降稈作用,A2的負(fù)效應(yīng)值最大,降稈作用明顯,利于選育矮稈高產(chǎn)組合。就穗長而言,A8的配合力最大,其次是A1,利于選育大穗型組合。結(jié)實小穗數(shù)的一般配合力A1最高,A8的負(fù)效應(yīng)值最大。總小穗數(shù)的一般配合力A7最大。穗粒數(shù)的一般配合力A8最大,其次為A7。單株穗數(shù)和單株粒重的一般配合力效應(yīng)值A(chǔ)4均最高,A1、A2次之。百粒重的一般配合力A7最高,A5次之。
2.3 特殊配合力效應(yīng)
特殊配合力主要是受基因非加性效應(yīng)控制,即基因的顯性和上位性效應(yīng)。從表4中可以看出,同一性狀不同組合間差異較大,且各個性狀的變異范圍較大。就株高而言其變異范圍為-20.13~17.40,A5B5組合株高SGA最低,降稈效果明顯;穗長的變異范圍-14.36~8.96,A4B5組合穗長SGA最高;結(jié)實小穗數(shù)的變異范圍-12.86~9.63,A2B5和A8B5組合SGA最高;總小穗數(shù)的變異范圍-12.47~8.79,A5B2組合的SGA最高;穗粒數(shù)的變異范圍-22.40~18.42,A2B3組合的SGA最高;穗下節(jié)間長的變異范圍-16.87~8.45,組合A5B1的SGA最高;單株穗數(shù)的變異范圍-37.32~74.62,其變異范圍最大,組合A4B2的SGA最高,并且該組合株高SGA為負(fù)效應(yīng),其它性狀除百粒重外均有較大的正效應(yīng);單株粒重的變異范圍-38.97~30.42,組合A4B4的SGA最高;百粒重的變異范圍-26.56~32.53,組合A1B2的SGA最高。 從表4還可看出,A4B4組合各個產(chǎn)量性狀的特殊配合力雖然都不是最高,但是單株粒重、單株穗數(shù)和百粒重的特殊配合力均為正效應(yīng),并且存在一定的降稈作用;該組合實際單株產(chǎn)量雖然不是最高,但是相對來說也較優(yōu)(25.54 g)。其余組合某一性狀也有很好的表現(xiàn),在選配組合時可以考慮它們某一性狀的特殊配合力。
2.4 配合力基因型方差和遺傳力的估計
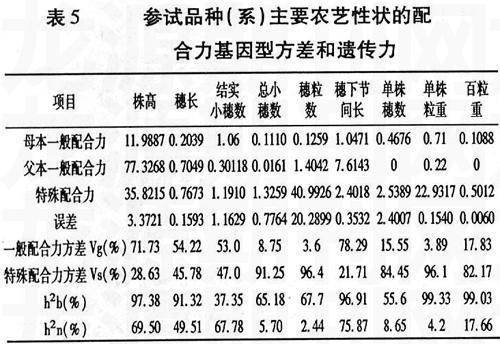
遺傳力是表示數(shù)量性狀遺傳過程中遺傳決定與環(huán)境影響相對重要性的數(shù)量指標(biāo),是重要的遺傳和育種參數(shù)。進(jìn)一步分析各性狀的配合力基因型方差和遺傳力(表5)可以看出,株高、穗長、結(jié)實小穗數(shù)和穗下節(jié)間長的Vg(%)大于Vs(%),

說明這5個性狀的加性作用相對于非加性作用更重要,而其它5個性狀的Vs(%)遠(yuǎn)遠(yuǎn)大于Vg(%),說明在這5個性狀中,非加性成分(顯性和上位性)更為重要。廣義遺傳率(h2b):單株粒重>百粒重>株高>穗下節(jié)間長>穗長>穗粒數(shù)>總小穗數(shù)>單株穗數(shù)>結(jié)實小穗數(shù);狹義遺傳率(h2n):穗下節(jié)間長>株高>結(jié)實小穗數(shù)>穗長>百粒重>單株穗數(shù)>總小穗數(shù)>單株粒重>穗粒數(shù)。9個性狀中有8個的廣義遺傳率大于50%,其中株高、穗長、穗下節(jié)間長、單株粒重和百粒重皆在90%以上,表明株高、穗長、穗下節(jié)間長、單株粒重和百粒重這5個性狀受環(huán)境的影響較小。株高、穗下節(jié)間長的狹義遺傳率為69.5%和75.87%,可在早期進(jìn)行選擇。穗長的狹義遺傳率中等,宜在F4代進(jìn)行選擇。結(jié)實小穗數(shù)、總小穗數(shù)、穗粒數(shù)、單株穗數(shù)、單株粒重和百粒重的廣義遺傳率較大,但狹義遺傳率較低,說明加性遺傳效應(yīng)較小,顯性效應(yīng)和上位性效應(yīng)較大,應(yīng)該適當(dāng)提高選擇的代數(shù)。
3 討論與結(jié)論
對新育成的優(yōu)良品系進(jìn)行配合力和遺傳力分析可以幫助我們深入了解新品系各個性狀的遺傳規(guī)律,同時為以后利用這些品系提供理論依據(jù)。前人在不同地區(qū)對不同的優(yōu)良品系進(jìn)行了大量的配合力和遺傳力分析,因為所選材料的不同導(dǎo)致其結(jié)果存在較大差異。歐俊梅等(2000)[13]認(rèn)為,株高、穗長、穗粒數(shù)和千粒重為加性和非加性效應(yīng)共同作用,主要受加性基因控制。何良榮等(2001)[8]研究表明,株高、每穗小穗數(shù)以加性效應(yīng)為主。龐紅喜等(2000)[9]則發(fā)現(xiàn)株高、穗長、每穗小穗數(shù)、穗粒數(shù)和千粒重的加性、顯性效應(yīng)均重要。曹亞萍等(2000)[10]指出,株高受基因加性、非加性和母體效應(yīng)的共同作用,以加性效應(yīng)占優(yōu)勢;千粒重取決于加性和非加性效應(yīng),穗粒數(shù)由加性效應(yīng)控制,而穗粒重存在明顯的核質(zhì)互作。姚金保等(2003)[3]認(rèn)為千粒重符合加性—顯性模型,以加性效應(yīng)為主。
本試驗選用山東農(nóng)業(yè)大學(xué)新選育的抗病、高產(chǎn)新品系作為母本,通過配合力和遺傳力分析評價其在育種中的應(yīng)用價值,為以后的育種工作奠定基礎(chǔ)。研究表明,在產(chǎn)量性狀方面母本的一般配合力要大于父本即母本對于后代產(chǎn)量的影響要大于父本;在株高方面,A1、A2、A3、A5和A8均具有較好的降稈作用,且A1、A2、A3和A5親本本身株高均低于70 cm,利于選育矮稈、高產(chǎn)品種。A1(530001)親本的穗長GCA較大,結(jié)實小穗數(shù)的GCA最大,可以作為大穗、多小穗型親本。A2株高的GCA最低,因此可以作為矮源親本。A4單株穗數(shù)和單株粒重的GCA均最高,可以作為多穗、高產(chǎn)親本。A5、 A6和A7其它性狀的GCA表現(xiàn)一般,但它們的百粒重配合力很高,可以作為大粒型親本。A8在穗粒數(shù)上GCA最大,可以作為多穗材料。
通過試驗發(fā)現(xiàn),株高、穗長、結(jié)實小穗數(shù)和穗下節(jié)間長這4個性狀由加性效應(yīng)和非加性效應(yīng)共同作用,其中以加性效應(yīng)為主,這和前人的研究基本一致。同時發(fā)現(xiàn)總小穗數(shù)、穗粒數(shù)、單株粒重、單株穗數(shù)和百粒重5個性狀主要受非加性效應(yīng)影響,其中總小穗數(shù)、穗粒數(shù)和單株粒重3個性狀非加性效應(yīng)占遺傳方差的90%以上。非加性效應(yīng)包括顯性效應(yīng)和上位性效應(yīng),其中顯性效應(yīng)是不能夠穩(wěn)定遺傳到后代的,而研究表明上位性效應(yīng)在遺傳效應(yīng)中具有重要地位并且在自花授粉作物中可以穩(wěn)定遺傳。如果上位性存在,將會給育種選擇帶來額外收益[14]。因此為了進(jìn)一步明確本試驗新品系的利用價值,需要研究上位性在這些性狀中的作用。
參 考 文 獻(xiàn):
[1] 田紀(jì)春. 超級小麥的概念、育種目標(biāo)和任務(wù)[J].山東農(nóng)業(yè)科學(xué),2004,5 :18 - 21.
[2] 曹亞萍,范紹強(qiáng),張 娟,等.冬小麥BYDV材料粒重的配合力與遺傳模型分析[J].山西農(nóng)業(yè)科學(xué),2002,30(1):17-20.
[3] 姚金保,王書文,姚國才,等.普通小麥千粒重的配合力與遺傳模型分析[J].南京農(nóng)學(xué)專報,2003,19(1):52-54.
[4] 張彩英,謝令琴,常文鎖,等.冬小麥品種資源沉降值配合力及其它性狀相關(guān)性研究[J].中國農(nóng)學(xué)通報,2001,17(4):15-17.
[5] 隋新霞,李根英,吳祥云,等.不同穗型冬小麥主要農(nóng)藝性狀配合力分析[J].山東農(nóng)業(yè)科學(xué),2004,2:20-22.
[6] 王 瑞,寧 錕,王 怡,等.普通小麥穗部性狀的配合力與遺傳模型分析[J].西北農(nóng)業(yè)學(xué)報,1996,5(1):1-5.
[7] 陳集賢,郜和臣,黃相國.十一個春小麥品種產(chǎn)量性狀的配合力與遺傳力分析[J].作物學(xué)報,1981,7(3):201-207.
[8] 何良榮,杜 娟,鄒春梅,等.大穗小麥產(chǎn)量性狀的配合力與遺傳力分析[J].新疆農(nóng)業(yè)科學(xué),2001,38(1):16-17.
[9] 龐紅喜,亢福仁,王成社,等.大穗型小麥主要農(nóng)藝性狀配合力分析[J].西北農(nóng)業(yè)學(xué)報,2000,9(2):78-81.
[10]曹亞萍,張明儀,喬合心,等.冬小麥產(chǎn)量性狀的配合力及遺傳規(guī)律研究[J].山西農(nóng)業(yè)科學(xué),2000,28(2):7-11.
[11]劉來福編著.作物數(shù)量遺傳[M].北京:農(nóng)業(yè)出版社,1982,211-281.
[12]唐啟義,馮明光.實用統(tǒng)計分析及其DPS數(shù)據(jù)處理系統(tǒng)[M].北京:科學(xué)出版社,2002.
[13]歐俊梅,李生容,杜小英,等.5個矮稈小麥品種(系)主要農(nóng)藝性狀的配合力和遺傳力分析[J].綿陽經(jīng)濟(jì)技術(shù)高等專科學(xué)校學(xué)報,2000,17(2):1-4.
[14]張文英,程君奇,朱 軍,等.上位性及其在遺傳育種中的應(yīng)用[J].生物信息學(xué),2004,2(2):39-43.
[15]Cheverud J M, Routman E J. Epistasis and its contribution to genetic variance components [J]. Genetics, 1995, 139: 1455-1461.
