24個香菇菌株的鑒定與分類研究
2009-01-15 09:03:36謝雪迎任鵬飛朱常香宮志遠(yuǎn)
山東農(nóng)業(yè)科學(xué) 2009年12期
關(guān)鍵詞:分類
謝雪迎 任鵬飛 朱常香 宮志遠(yuǎn)
摘 要:采用拮抗試驗和酯酶同工酶譜分析兩種生物學(xué)方法對24個香菇菌株進行鑒定和分類,構(gòu)建樹狀圖。結(jié)果表明:拮抗試驗受外界環(huán)境影響大,可用于香菇菌株的初步分類;酯酶同工酶譜帶豐富,可以完成香菇菌株的鑒定,在聚類圖譜0.84的水平上24個香菇菌株可以分為10大類。
關(guān)鍵詞:香菇;分類;拮抗;酯酶同工酶
中圖分類號:S646.1+20.37 文獻標(biāo)識號:A 文章編號:1001-4942(2009)12-0036-04
香菇(Lentinula edoeds)是世界上最主要的食用菇類栽培品種之一,在食用菌的產(chǎn)量構(gòu)成中占有重要地位,1994~1995年世界食用菇類總產(chǎn)量達(dá)490×104 t,其中香菇占15%[1]。由于其味美質(zhì)嫩和具有增強人體免疫功能的特殊作用,香菇已成為全球性備受歡迎的功能食品。
我國是世界香菇栽培的起源地,從發(fā)明最原始的“砍花法”以來,迄今已有1 500多年的栽培歷史[2]。我國擁有世界上最豐富的香菇種質(zhì)資源,在香菇生產(chǎn)的三大要素(菌種、培養(yǎng)料、栽培管理)中,菌種是最重要的基礎(chǔ)生產(chǎn)資料,是影響香菇產(chǎn)量及質(zhì)量構(gòu)成的關(guān)鍵因子。我國香菇生產(chǎn)快速發(fā)展的主要原因之一,就是引進和選育了一大批優(yōu)質(zhì)、高產(chǎn)、抗逆性強的菌種。由于目前香菇菌株同名異物和同物異名現(xiàn)象嚴(yán)重,故對香菇菌株進行有效的分類和鑒定就成了香菇新品種選育的基礎(chǔ)和前提。本試驗對24個香菇菌種通過拮抗和酯酶同工酶試驗進行了分類和鑒定,旨在為后來的選育工作提供依據(jù)[3]。
1 材料與方法
1.1 供試菌株
供試的香菇菌株見表1。
1.2 試驗方法
1.2.1 拮抗試驗 挑取大米粒大小的兩菌株種塊于90 mm培養(yǎng)皿內(nèi)對峙培養(yǎng),種塊間距1.5~2.0 cm。每個菌株重復(fù)3次,于25℃培養(yǎng)20 d,觀察結(jié)果并拍照。
1.2.2 菌絲體培養(yǎng) 參考林芳燦的方法[4],將參試菌株在相同條件下接種于PDY液體培養(yǎng)基中進行淺層培養(yǎng),培養(yǎng)量以各自可以做3個平行試驗為準(zhǔn)。
1.2.3 同工酶分析 收集供試菌株菌絲體各3份,液氮研磨。將研磨后的樣品在4℃、10 000 r/min離心10 min,取其上清液。將上清液、蔗糖溶液和溴酚藍(lán)指示液按5∶1∶1的體積比混合作為電泳樣品。經(jīng)SDS-PAGE電泳,取分離膠染色,拍照。
1.2.4 試驗結(jié)果的觀察和統(tǒng)計分析 觀察樣品酶帶的位置,計算每條酶帶的相對遷移率(Rf)。Rf按下列公式計算:
Rfn=d璶/d
式中:Rfn為條帶n的相對遷移率;d璶為條帶n的遷移距離(cm);d為溴酚藍(lán)的遷移距離(cm)。
同時將凝膠照相存圖。根據(jù)菌株每條酶帶的出現(xiàn)與否,計算各菌株間的遺傳相似系數(shù)。
2 結(jié)果與分析
2.1 拮抗試驗
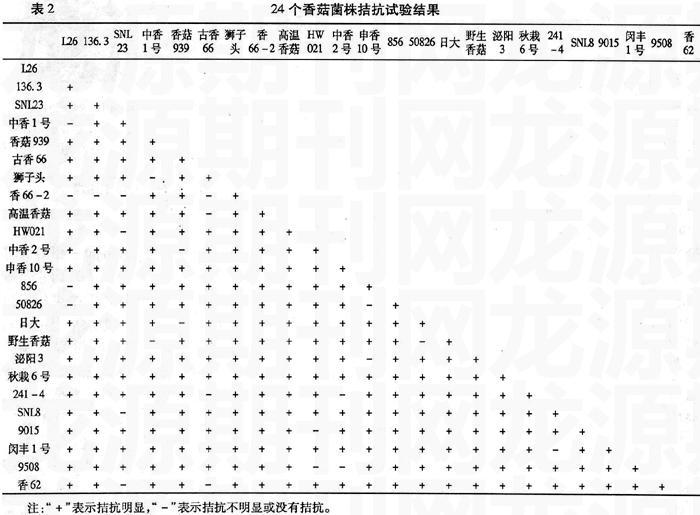
24個香菇菌株共設(shè)計拮抗組合276組,拮抗現(xiàn)象不明顯的有25組,占9.1%,其中L26與中香1號、香66-2、856、50826沒有拮抗,親緣關(guān)系較近;SNL23與HW021、SNL8、香62親緣關(guān)系較近;中香1號與獅子頭、野生香菇親緣關(guān)系較近;香939與中香2號、日大親緣關(guān)系較近。其它組合的11個菌株拮抗現(xiàn)象比較明顯。因而拮抗試驗結(jié)果可將24個香菇菌株初步分為14類(圖1、表2)。
2.2 酯酶同工酶譜分析
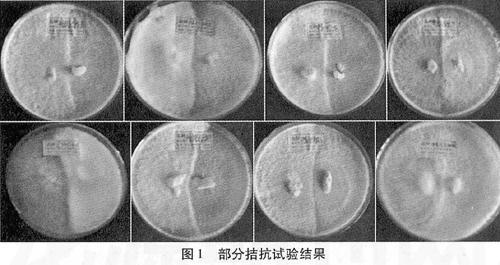
提取24個香菇菌株的酯酶同工酶進行聚丙烯凝膠電泳,共得到73條譜帶,各菌株譜帶1~5條不等,相對遷移率介于0.29~0.92之間(見表3)。各菌株間譜帶的深度和寬度有明顯差異,寬且深的譜帶說明酶含量高,活性大;窄且淺的表明酶含量低,活性小。根據(jù)譜帶的深淺和寬窄,可將所有譜帶分為四級:一級酶帶色深帶寬;二級酶帶色較深而寬;三級酶帶色較淺而窄;四級酶帶色淺而窄(圖2)。

2.3 聚類分析
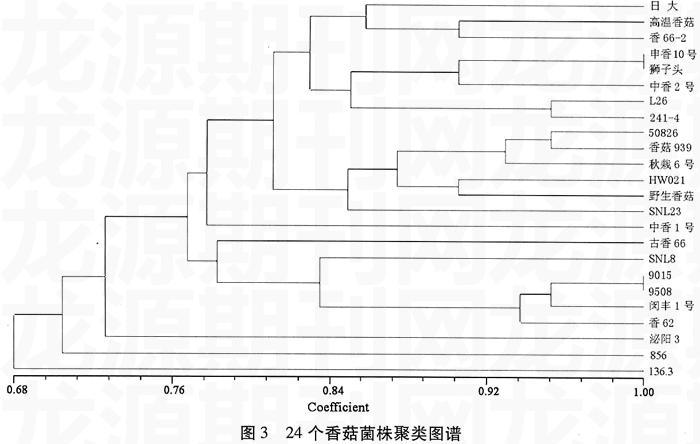
根據(jù)酯酶同工酶電泳結(jié)果,將同一相對遷移率位置上有條帶記為1,無條帶記為0,構(gòu)建矩陣并用NTSYS進行分析[5]。聚類分析結(jié)果(圖3)表明,所有參試菌株在0.68的相似水平上可以歸為一類,在1.00水平上歸為22類,變異范圍為0.68~1.00。在1.00水平上,9015和9508、獅子頭和申香10號相似系數(shù)很高,可認(rèn)為是一個菌株。在0.84水平上分為10大類,其中136.3、856、泌陽3、SNL8、古香66、中香1號各成一類;香62、閔豐1號、9508和9015歸為一類;SNL23、野生香菇、HW021、秋栽6號、香菇939、50826歸為一類;日大、高溫香菇、香66-2歸為一類;申香10號、獅子頭、中香2號、L26、241-4歸為一類。
3 結(jié)論與討論
與生化標(biāo)記、分子生物學(xué)標(biāo)記相比,拮抗試驗結(jié)果主觀性強,菌絲生長狀況受外界環(huán)境的影響很大,往往使觀察結(jié)果與其它鑒定手段相悖。本試驗中,在沒有拮抗線的試驗組合中,也能觀察出菌絲生長狀況的明顯不同,這說明其菌株遺傳背景差異還是有的,只不過沒有在平板上顯現(xiàn)出來而已。所以拮抗試驗只能用于菌株的初步分類。
在生化標(biāo)記中,酯酶同工酶的穩(wěn)定性最好,是目前菌種鑒定的常用方法。但酯酶同工酶的測定對培養(yǎng)條件的一致性要求較高,并且不同菌株、不同發(fā)育時期、不同部位的酶譜也不一致[6]。故所有處理必須具備相同的培養(yǎng)條件和提取條件,才能保證酶譜分析的客觀性。本試驗中,香菇共同性譜帶不清楚,可能與提取條件有關(guān)。
聚類分析通過酶譜條帶的有無構(gòu)建矩陣,并利用NTSYS軟件繪制樹狀圖、計算相關(guān)系數(shù)。但在多大的遺傳系數(shù)上進行分類最為合理,需要進一步的研究。本試驗的遺傳系數(shù)介于0.68~1.00之間,所取的分類點為0.84,是兩者的平均數(shù)。
參 考 文 獻:
[1] Chang shu-ting.Mushroon Biology and Mushroon Products[M].Chinese University press,1997,1-10.
[2] 楊新美.中國食用菌栽培學(xué)[M]. 北京:中國農(nóng)業(yè)出版社,1988,292-294.
[3] 呂作舟,劉巧云.我國香菇育種的回顧與展望[J].中國食用菌,1996,15(1):5-6.
[4] 林芳燦,高國琪.香菇主要數(shù)量性狀遺傳率及相關(guān)性研究[J].華中農(nóng)業(yè)大學(xué)學(xué)報,1993,12(1):27-30.
[5] 蓋均鎰.試驗統(tǒng)計方法[M].北京:中國農(nóng)業(yè)出版社,2000,35-46
[6] 王 鐳,張 婤,潘迎捷,等.利用方差分析和聚類分析法進行香菇種質(zhì)資源評價[J].食用菌學(xué)報,1997,4(3):21-24.
猜你喜歡
西北民族大學(xué)學(xué)報(自然科學(xué)版)(2021年4期)2021-12-29 02:54:24
數(shù)學(xué)小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
小聰仔(科普版)(2020年12期)2021-01-18 09:16:52
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
學(xué)生天地(2019年32期)2019-08-25 08:55:22
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2019年4期)2019-05-20 10:06:32
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2018年6期)2018-06-26 08:36:06
小天使·一年級語數(shù)英綜合(2017年11期)2017-12-05 18:49:56
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
