擬南芥18srRNA參與隱花色素介導的信號傳導途徑的研究*
2010-03-06 02:59:20汪啟明屠小菊唐冬英趙小英劉選明
湖南大學學報(自然科學版) 2010年12期
汪啟明,屠小菊,唐冬英,趙小英,劉選明?
(1.湖南大學生物學院;化學生物傳感與計量學國家重點實驗室,湖南長沙 410082; 2.湖南農業大學生物科學技術學院,湖南長沙 410128)
擬南芥18srRNA參與隱花色素介導的信號傳導途徑的研究*
汪啟明1,2,屠小菊1,唐冬英1,趙小英1,劉選明1?
(1.湖南大學生物學院;化學生物傳感與計量學國家重點實驗室,湖南長沙 410082; 2.湖南農業大學生物科學技術學院,湖南長沙 410128)
通過篩選以擬南芥藍光受體隱花色素雙突變體cry1cry2為遺傳背景的激活標簽突變體庫,得到一株SCC98-D株系,該突變體具有早開花,下胚軸變短,并具有花瓣數目增加等表型,徹底恢復了cry1cry2的晚開花和長下胚軸表型.通過Tail-PCR的方法克隆得到SCC98-D突變體中激活標簽插入位點的側翼序列,通過對該側翼序列測序發現插入位點在18srRNA的1751bp處,運用RT-PCR方法分析插入位點周圍基因的表達,發現只有18srRNA的表達被激活,由此證實了18srRNA參與隱花色素介導的信號傳導途徑.
擬南芥;隱花色素;激活標簽突變體;18s核糖體RNA
植物的藍光受體對植物的生長發育起著非常重要的調控作用,擬南芥的藍光受體包括隱花色素(cryptochrome)和向光素(phototropin)[1],隱花色素能夠調控擬南芥長日照條件下的開花時間、光形態建成、氣孔開放等一系列的生長發育過程[2-3],現代的植物分子生物學的研究結果發現,擬南芥體內主要發揮功能的隱花色素是CRY 1和CRY2[4-6].
近20年的研究發現,隱花色素能夠通過與常數基因(Constans),光形態建成負調控因子COP1,鈣調蛋白SUB1以及轉錄因子CIB1等蛋白相互作用,將信號傳遞給下游基因從而調控植物的多個生長發育過程[7-11].然而,對藍光受體隱花色素調控下游基因表達的分子機制的了解仍不透徹[12],詳細了解植物的光信號傳遞途徑對于合理利用植物有益基因資源和開發生物能源有著非常重要的意義,所以,研究與隱花色素直接相互作用或者參與隱花色素介導的信號途徑的基因是現代植物分子生物學的研究熱點.
本文以隱花色素雙突變體cry1cry2為實驗材料,通過激活標簽技術構建了T-DNA插入突變體庫.通過篩選具有開花時間提前,植株矮化等表型的SCC突變體(sup press cry1cry2,抑制cry1cry2表型的突變體),并選取突變體庫中表型為顯性的突變體,命名為sup press cry1cry2-dom inant(SCC-D),同時采用 TA IL-PCR(Thermal asymmetric interlaced-PCR,熱不對稱巢式PCR)和RT-PCR方法對突變體進行分析,發掘被激活的基因,為闡明藍光受體隱花色素發揮功能的分子機制奠定基礎.
1 材料與方法
1.1 植物材料
擬南芥野生型(wild-type,W T)為col-4,哥倫比亞生態型;突變體cry1cry2:其遺傳背景為 col-4;突變體SCC98-D是以cry1cry2為受體,通過插入激活標簽篩選分離而來.
1.2 植物培養
擬南芥種子用體積分數為70%酒精處理30 s,然后用體積分數為0.1%升汞滅菌8m in,無菌水沖洗4~5次,均勻播于MS鹽+質量分數0.8%瓊脂的培養基上.4℃處理4 d后,連續照白光12 h,以促進種子萌發,并將植物轉入22℃培養室,分別在長日照白光(16光照,8 h黑暗)和黑暗處理培養后觀察苗期表型,以及在培養土中長日照白光(16光照,8 h黑暗)培養觀察開花時間.本試驗中用的藍光、紅光、遠紅光和白光光源分別為:LED-B(波長為470 nm,半幅寬為30 nm),LED-R(波長為660 nm,半幅寬為20 nm),LED-FR(波長為740 nm,半幅寬為25 nm)和白色熒光燈(飛利普).藍光、紅光和白光的光照強度用Li-250量子光度計測量.遠紅光的強度先用Li-250量子光度計測量,然后根據分光照度計測得的標準曲線進行估計.
1.3 質粒
激活標記載體pSKI015(見圖1)由美國加州大學洛杉磯分校林辰濤教授提供.
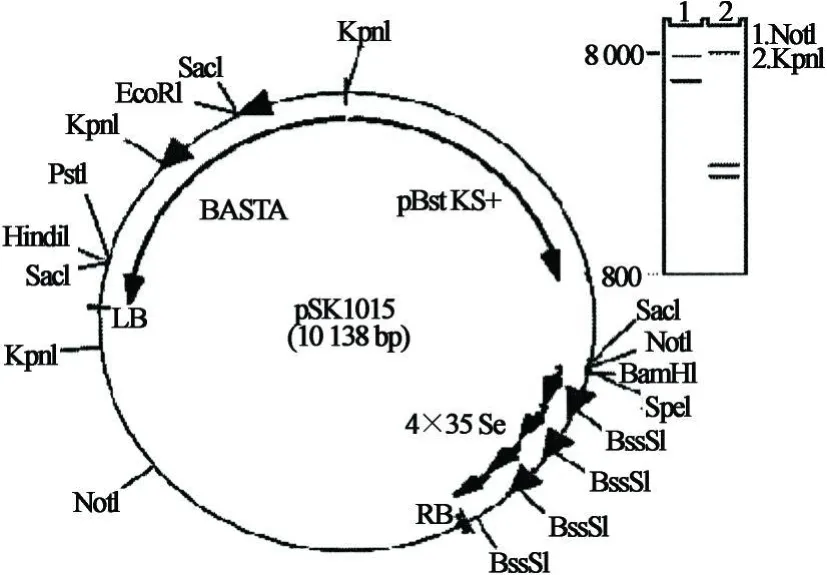
圖1 激活標簽載體pSK I015示意圖Fig.1 Diagram of activation tagging vector pSK I015
1.4擬南芥總DNA提取
液氮充分研磨植物材料,將75 mg新鮮材料樣品中加400μL Buffer AP1,4μL RNase A,充分混勻;在65℃培養10 min,期間上下顛倒離心管2~3次;在130μL Buffer AP2,混勻,冰上放置 5 m in, 14 000 rpm離心5 min;將上清轉入含2 m L收集管的過濾柱內,10 000 r/m in離心2min;將下清轉入新的1.5 m L離心管內,加1.5倍體積的 Buffer AP3/E,顛倒3~4次,混勻;把溶液轉入含2 m L收集管的吸附柱內,10 000 rpm離心1~2 min;如果一次過不完柱,可多次加樣直至過濾完成,倒掉收集管中的液體;加500μL Buffer AW 1,10 000 rpm離心1 m in,棄下清;加 500μL Buffer AW 2,10 000 rpm離心 1 min,棄下清;重復上一步操作一次; 14 000 rpm離心3 min,將離心柱轉移至1.5m L離心管內;加50~100μL Buffer AE至吸附柱濾膜上,室溫靜置 1~5 min,10 000 rpm 離心1 m in;將離心管中DNA溶液電泳檢查后,置于-20℃保存.
1.5 T-DNA插入突變體側翼序列的克隆
TA IL-PCR是一種比較成熟的擴增T-DNA插入側翼序列的技術[13-15].本實驗利用該技術來擴增突變體載體左臂的基因組DNA側翼序列.根據pSKI015載體的序列,TAIL-PCR反應的3個特異性引物如下:LS1 5'-GACAACATGTCGAG-GCTCAGCAGGA-3';LS2 5'-TGGACGTGAATGTAGACACGTCGA-3';LS3 5'-GCTTTCGCCTATAAATACGACGG-3'.其中 LS3離 T-DNA的左臂最近,這3個特異性引物的 Tm值較高,為60℃相鄰.6個簡并引物為AD1:5'-NTCGASTWTSGWGTT-3';AD2:5'-NGTCGASWGA NAWGAA-3';AD3:5'-WGTGNAG-WANCANAGA-3';AD4:5'-AG-WGNAGWANCAWAGG-3';AD5:5'-GNAGT-GWSANCAA-GA-3';AD6:5'-STTGNTASTNCTNTGC-3'.這6個簡并引物的 Tm值較低,均為 45℃左右. TAIL-PCR的擴增條件為第1輪PCR:4℃,2 min;93℃,1 min;95℃,1 min;(94℃,30 s;62℃, 1min;72℃,2.5 min)5個循環;94℃,30 s;25℃, 3min;升溫0.2 ℃/s,升至72 ℃(約需4 min);72℃,2.5min;(94℃,10 s;68℃,1m in;72℃,2.5 min;94℃,10 s;68℃,1 min;72℃,2.5 m in;94℃,10 s;44℃,1 min;72℃,2.5 min)15個循環;72℃,5min.第2輪PCR以第1輪的產物稀釋50倍后取1μL為模板,擴增條件為4℃2m in;(94℃,10 s;64℃,1 min;72℃,2.5 min;94℃,10 s; 64℃,1 min;72℃,2.5 min;94℃,10 s;44℃,1 min;72℃,2.5 min)12個循環;72℃,5min.第3輪PCR以第2輪的產物稀釋50倍后取1μL為模板,擴增條件為4℃,2 min;(94℃,15 s;44℃,1 min;72℃,2.5 min)20個循環;72℃,5min.3輪PCR的反應體系均為20μL.
1.6 總RNA提取及cDNA第1鏈的合成
1)采用安比奧公司生產的 RNA提取試劑盒,提取總RNA;
2)按照試劑說明書在總RNA溶液中加適量的Rnase(無DNase I)和10×Buffer,37 ℃水浴30min;
3)加水使溶液體積達500μL,然后加等體積氯仿混勻;
4)4℃,12,000 rpm 離心10m in,取上清,然后加2.5倍體積無水乙醇混勻,-20℃放置30 min;
5)4℃,12000 r/m in離心15min,然后用70%乙醇洗滌沉淀1~2次,真空干燥后將沉淀溶于適量的無RNase水,通過熒光分光光度計,進行樣品濃度和純度的測量,純度合格的RNA樣品-80℃保存備用.
6)合成cDNA第1鏈的逆轉錄反應體系為25 μL,首先在無 RNase的離心管中加約2μg的總RNA和Promega公司0.5μg Olig(dT)15,體系不能超過14μL,混勻,4℃短暫離心,70℃水浴5min后,立即冰浴冷卻.短暫離心后,依次加入10×Buffer 2.5μL,Genview 公司 10 mM dNTPs 1.25μL, MBI公司RNaseA抑制劑25 u,Promega公司MM LV逆轉錄酶 200 u,加無RNase的水至25μL, 42℃反應 60m in后,70℃水浴10 min使酶失活,即得cDNA第1鏈.
1.7 半定量RT-PCR
半定量RT-PCR(Sem i-quantitative PCR)引物設計參照文獻介紹的方法[16-17],根據半定量PCR產物長度限定在300~700 bp范圍內以及PCR產物序列盡可能靠近目的基因序列的3'端的要求,引物的設計盡可能滿足以下幾個條件:引物最適宜長度為20~22 bp;最適宜退火溫度為60~62℃;G+ C含量最好大于50%.根據上述要求,試驗中采用Primer Premier 5.0軟件對目的基因的PCR引物進行了設計,然后由上海生工生物技術有限公司合成,引物序列如表1所示.
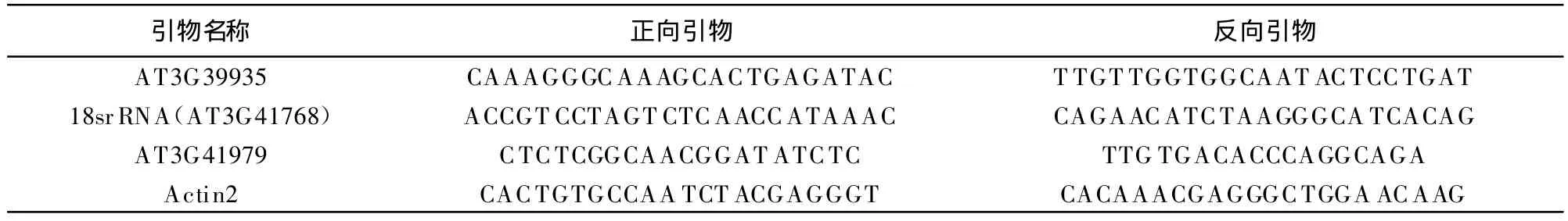
表1 半定量PCR所用引物的序列Tab.1 Primers used in sem i RT-PCR
2 結果與分析
2.1 突變體SCC98-D的表型分析
通過篩選突變體庫得到抑制cry1cry2表型的顯性突變體(SCCD).本實驗以突變體SCC98-D為研究對象,通過對其表型觀察,發現在藍光培養條件下該突變體與野生型相比,具有下胚軸變短(見圖2)并且長日照條件下成花時間提前(見圖3),花器官數目增加的表型(見圖4).
為對突變體SCC98-D的表型變化進行定量分析,實驗選取培養7 d的SCC98-D,WT和cry1cry2為材料,研究了不同光質光以及同一光質的不同光照強度下突變體SCC98-D下胚軸長度的變化,結果如圖2所示:在白光或者藍光下突變體SCC98-D的下胚軸比母本cry1cry2的要短,很好地抑制其遺傳表型,恢復到野生型Col-4的表型,但是暗培養和紅光培養條件下三者之間則無差異.
觀察上述材料在長日照條件下生長的開花時間,并記錄開花時間的差異.見圖3,結果發現:突變體SCC98-D在長日照條件下開花時間提前,只有19 d就抽苔,比野生型Col-4的始花時間還要提前,而在短日照條件下始花時間無差異.由此說明,突變體SCC98-D 完全抑制了cry1cry2的開花表型,該突變體中被激活的基因極有可能是CRY信號傳導途徑的下游基因.
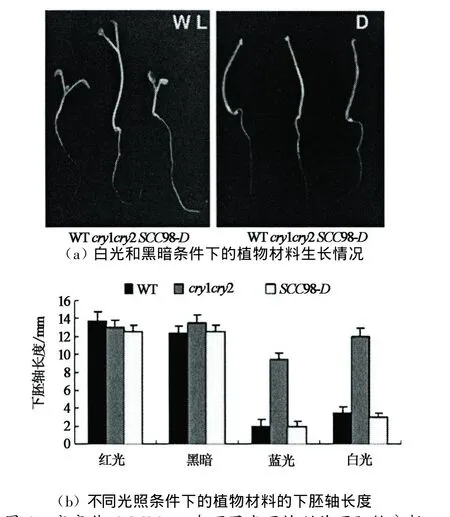
圖2 突變體SCC98-D在不同光下的幼苗下胚軸分析Fig.2 Hypocotyl lengths of SCC98-D mutant seedlings in different lights
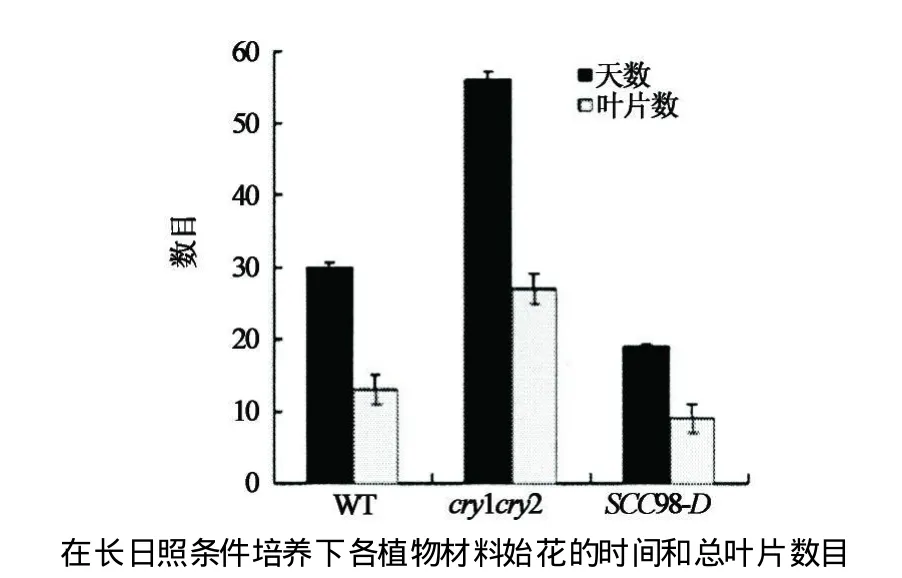
圖3 突變體SCC98-D在長日照(LD)下早開花Fig.3 SCC98-D mutant early flow er under LD
觀察花的發育情況,結果如圖4所示:突變體cry1cry2的花器官的發育正常,與擬南芥野生型的數目和排列方式都一致;突變體SCC98-D花器官的數目增加,排列方式也不是對生,并且每朵花花瓣數目的增加并不一致,從5到9都有,但是大多集中在5枚花瓣.這種花器官的異常發育與突變體dbb1a的花器官突變表型又存在差異,突變體SCC98-D花器官的數目只有增加,而突變體dbb1a的花器官的數目有增加也有減少.突變體SCC98-D的表型說明,其體內被激活的基因可能參與隱花色素介導的光對擬南芥下胚軸伸長的抑制和光周期調控植物開花的信號傳導,是CRY的下游基因.
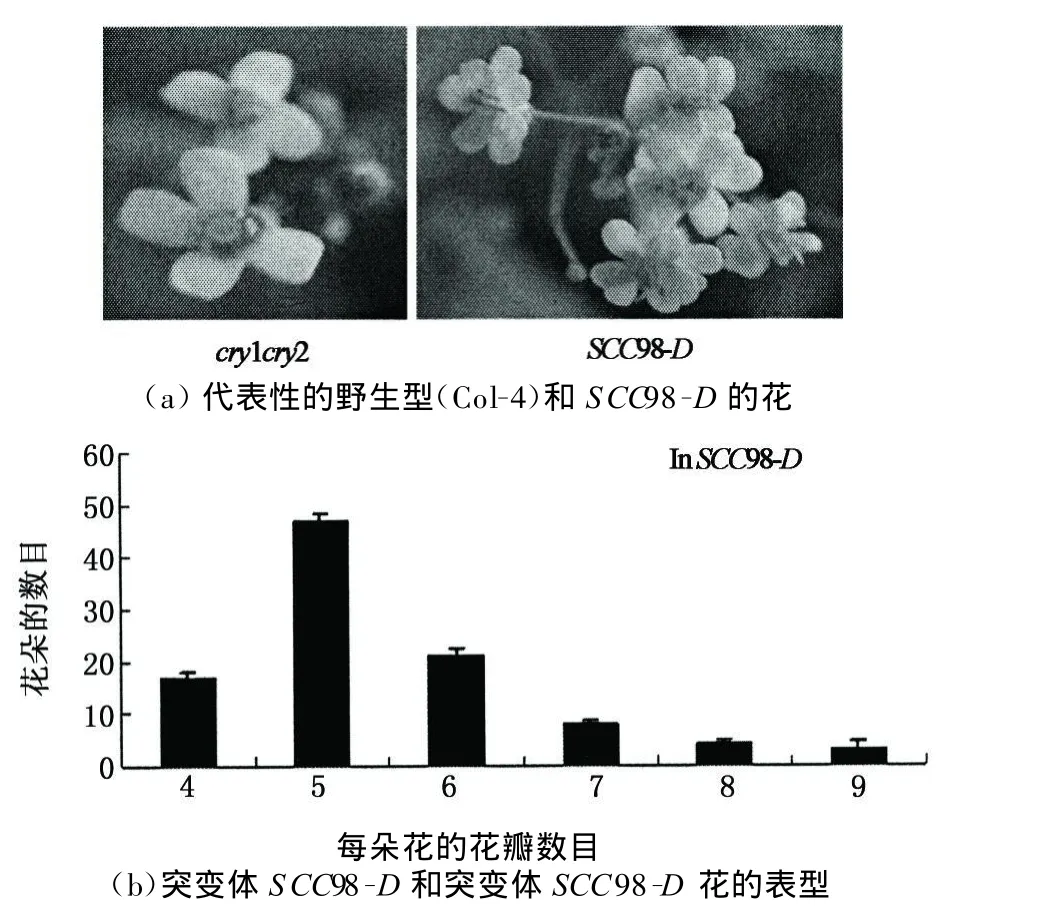
圖4 野生型(CoL-4)和突變體SCC98-D花的表型Fig.4 Flow er o f W T(Col-4)and SCC98-D
2.2 突變體SCC98-D中T-DNA插入位點及被激活基因的確定
為確定突變體SCC98-D的T-DNA插入位點位,采用TA IL-PCR方法克隆出T-DNA插入位點側翼序列,圖5(a)是TAIL-PCR產物的電泳檢測結果,泳道 M 是分子 m arker;泳道 1,4,7分別為SCC91-D,SCC101-D和SCC98-D的總DNA為模板的TAIL-PCR的第1輪PCR產物,泳道 2,5,8分別為對應的TAIL-PCR的第2輪PCR產物,泳道3,6,9分別為對應的TAIL-PCR的第3輪PCR產物.由圖4(a)可見,突變體SCC98-D為模板的TAIL-PCR的第3輪PCR產物有電泳條帶,只略小于其第2輪PCR產物.該結果說明,擴增得到突變體SCC98-D的T-DNA插入位點的側翼片段.
接著通過將克隆片段測序,并將克隆片段與擬南芥基因組序列比對發現突變體SCC98-D中的TDNA插入位點位于18srRNA(18srRNA)的3'端第1 751 bp處.為進一步驗證T-DNA插入位點位,通過PSKI015載體上的特異序列與18srRNA序列設計上的引物,對各種材料的總DNA進行PCR檢測,結果如圖5(b)所示:泳道 M 是分子 marker,泳道1~3分別是以 SCC98-D,W T(Col-4)和cry1cry2的總PCR鑒定電泳,以突變體SCC98-D的DNA為模板的PCR產物中有1個1 000 bp左右的電泳條帶,其與目的片段大小相同;而野生型和cry1cry2中均未擴增出目的條帶.由此進一步證明突變體SCC98-D的T-DNA插在18srRNA的3'端第1 751 bp處,其T-DNA插入位點模式圖如圖6 (a)所示.
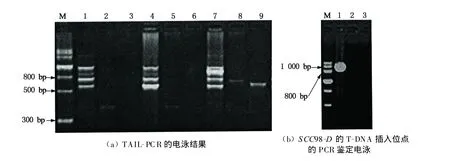
圖5 SCCD突變體插入位點側翼序列的獲得Fig.5 Acquisition of f lanking genom ic sequence o f inserted site of SCCD mutants
為進一步挖掘突變體SCC98-D中被激活的基因,本實驗通過RT-PCR方法分析插入位點相鄰的基因在野生型Col-4,cry1cry2和SCC98-D中的表達差異,確定被激活標簽PSKI015激活的基因.但是,由于18srRNA是18srRNA,相鄰的基因非常少,大多是無功能的核酸序列,而僅有的基因也是是轉坐子基因或沒有具體功能的假基因,所以只選擇檢測A t3g3995,A t3g41979和18srRNA這3個基因.結果如圖6(b)所示:SCC98-D中T-DNA插入位點所在的18srRNA基因在野生型中有表達,但在cry1cry2中表達量很少,而在SCC98-D中表達量很高;各突變體中A t3g3995和A t3g41979基因的表達量均無明顯差異.該結果說明:突變體SCC98-D所具有的表型可能是由于18srRNA基因表達被激活而引起的.
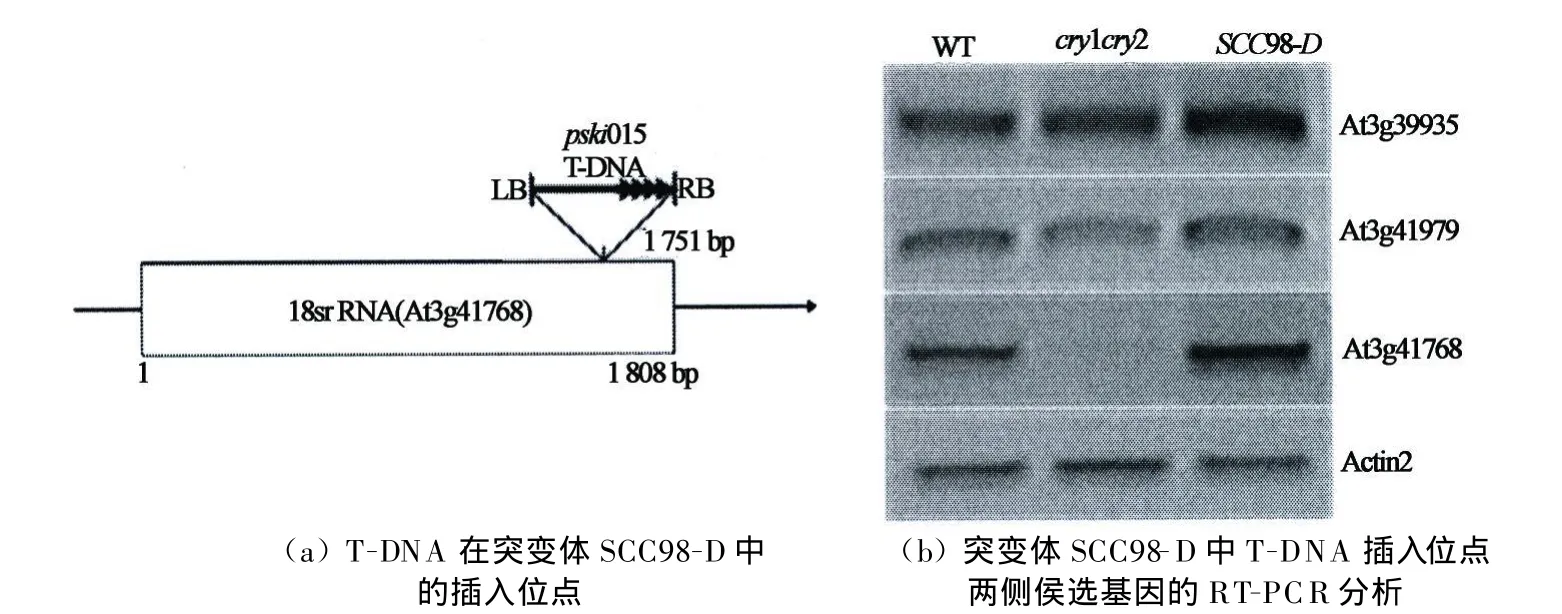
圖6 突變體SCC98-D的T-DNA插入位點及被激活基因分析Fig.6 The T-DNA insertion location and the actived gene in the SCC98-D mutants
3 討 論
18srRNA是核糖體小亞基的組成部分,參與到植物體內蛋白質的翻譯.此外,生物體中的某些基因的5'端非翻譯區結合可以與18srRNA所包含的核糖體內進入位點(internal ribosome entry sites IRES)相結合,從而導致這些特異基因進行不依靠帽子結構翻譯(Cap-independent translation).這種翻譯機制可以幫助生物體在受到脅迫或緊急狀態下得到需要的蛋白[18].通過分析18srRNA基因的互補序列,發現其1 750~1 765 bp區域為核糖體內進入位點,并且與受光誘導表達的鋅指蛋白DBB1a基因5'端非翻譯區中的一段序列一致.
此外,通過酵母雙雜交實驗發現隱花色素CRY2能與真核翻譯的啟始因子eif4E相互作用(未發表實驗結果),而eif4E在真核生物蛋白翻譯復合體中的關鍵蛋白,負責識別m RNA的帽子結構,并且在不依靠帽子結構的蛋白翻譯過程也發揮著關鍵作用[19].因此推測,突變體SCC98-D抑制cry1cry2表型的功能可能與18srRNA基因介導的DBB1a基因表達相關.
本研究發現SCCD 98-1突變體能完全恢復cry1cry2的表型,且其中的18srRNA表達量上升,由此推測18srRNA可能通過參與不依靠帽子結構的蛋白翻譯過程,調控光信號途徑蛋白的表達,從而介入隱花色素介導的信號傳遞途徑.然而植物藍光信號傳導途徑非常復雜,而調控方式也極具多樣性,因此18srRNA、隱花色素、eif4E以及 DBB1a之間詳細的調控機制將有待于進一步研究,為全面揭示藍光受體隱花色素介導的信號傳遞途徑提供理論基礎.
[1] CHEN M,CHORY J,FANKHAUSERC.Ligh t signal transduction in high t plan ts[J].Annu Rev Genet,2004,38(1):87 -117.
[2] LIN C.Plant blue-light recep tors[J].T rend s Plan tSci,2000, 5(2):337-342.
[3] TERZAGH IW B,CASHMORE A R.Light-regulated transcription[J].Annual Review of Plant Physiology and Plant M olecular Biology,1995,46(2):445-474.
[4] LIN C,YANG H,GUO H,etal.Enhancement of blue-light sensitivity ofarabidopsis seedingsby ab lue light recepto r cryptoch rome 2[J].Proc Natl Acad Sci USA,1998,95(26):2686 -2690.
[5] LIN C.Blue ligh t recep tors and signal transduction[J].Plant Cell,2002,14(S1):207-225.
[6] ZHAO XY,YU XH,FOO E,et al.A study of gibberellin homeostasis and cryp tochrome-mediated blue ligh t inhibition of hypocotyls elongation[J].Plant Phy siology,2007,145(8): 106-118.
[7] GUO H,YANG H,MOCKLER T C,et al.Regu lation of flowering time by A rabidopsis photo receptors[J].Science, 1998,279(2):1360-1363.
[8] DENG X,MA TSUIM,WEIN,eta l.COP1,an A rabidop sis regulatory gene,encodes a protein with both a zinc-binding m otif and a Gβhomologous dom ain[J].Cell,1992,71(4): 791-801.
[9] LIU LJ,ZHANG YC,LIQH,eta l.COP1-mediated ubiquitination of constans is implicated in cryptoch rom e regulation of flow ering in A rabidopsis[J].Plant Cell,2008,20(9):292-306.
[10]LIU H T,YU XH,LI KW,et al.Photoexcited CRY 2 interactsw ith CIB1 to regulate transcription and floral Initiation in A rabidopsis[J].Science,2008,322(5907):1535-1539.
[11]YU XH,SA YEGH R,MAYMON M,etal.Formation of nuclear bodies of A rabidop sis CRY 2 in response to blue light is associated w ith itsb lue ligh t-dependent degradation[J].Plant Cell,2009,21(1):118-130.
[12]LIZ R,W AKA I S,FISCHER B B,et a l.Sensing and responding to excess ligh t[J].Annu Rev Plant Biol,2009,60 (9):239-260.
[13]LIU Y,M ITSUKAWA N,OOSUM I,et al.Efficient isolation and m apping of arabidop sis thaliana T-DNA insert junctions by thermal asymmetric in terlaced PCR[J].Plant J, 1995,8(13):457-463.
[14]LIU Y,CHEN Y.High-efficiency thermalasymm etric interlaced PCR for am plification of unknow n flanking sequen ces [J].Biotechniques,2007,43(5):649-654.
[15]王愛榮,吳智芳,張麗麗,等.影響擬南芥轉化效率和激活標簽丟失的因素分析[J].福建農林大學學報:自然科學版,2006,35 (3):298-302.
WANG A R,WU Z F,ZAHGN L L,etal.Analy sisof factors affecting transformation efficiency and lossof enhancer in generation of activation-tagged m utan ts of arabidopsis thaliana [J].Jou rnal of Fujian Agricultu re and Forestry University: Natu ral Scien ce Edution,2006,35(3):298-302.(In Chinese)
[16]SCOFIELD S,DEWITTE W,M urray J A H.The KNOX gene SHOOT MERISTEML-ESS is required for the development of reproductivem eristematic tissues in A ra-bidopsis[J]. Plant Jou rnal,2007,50(5):767-781.
[17]CARLES CC,CHOFFNES-INADA D,REV ILLE K,et a l. U LTRAPETALA 1 en-codes a SANND dom ain putative transcriptional regu lator that con trolsshoot and floralmeristem activity in A rabidopsis[J].Developmen t,2005,132(11):897-911.
[18]VANDERHAEGHEN R,DECLERCQ R,KARIM IM,eta l. Leader sequenceof a plant ribosomalp rotein genew ith com plementarity to the 18S rRNA triggers in vitro cap-independent translation[J].FEBS LETTERS,2006,580(11):2630-2636.
[19]KATRIEN V D K,RUDIi B,DIRK I,eta l.T ranslation control of eukaryotic gene ex pression[J].In forma H eathcare, 2009,44(4):143-168.
A Study of 18srRNA Invovled in Cryptochrome-mediated Signaling in A rabidopsis
WANG Qi-ming1,2,TU Xiao-ju1,TANG Dong-ying1,ZHAO Xiao-ying1,LIU Xuan-ming1?
(1.State Key Laboratory forChemical Biosensing and Chemometrics,Hunan Univ,Changsha,Hunan 410082,China; 2.School of Biological Science and Technology,Hunan Agricultural Univ,Changsha,Hunan 410128,China)
SCC98-D was screened from an activation-tagging mutant library in the background of blue light-recep tor cryp tochrome doub lemutant cry1cry2 in A rabidopsis,which was early flowering,short hypocotyl and excess petalnum ber,and then the phenotype of late flowering and long hypocoty l in cry1cry2 was suppressed.The flanking sequence of inserted sitewas cloned w ith TAIL-PCR.The sequenced result of flanking sequence has shown that the T-DNA w as inserted in 1751 bp of 18srRNA.A fterwards,a transcriptional analysisof the gene around inserted sitewith RT-PCR demonstrated that the activated genewas 18srRNA.This study has identified that 18srRNA is involved in Cry-mediated signaling.
arabidopsis;cryptochrome;activation-tagging mutant;18srRNA
O613.52
A
1674-2974(2010)12-0066-06 *
2010-06-21
國家自然科學基金資助項目(30770200),教育部高校博士點基金資助項目(755228001)
汪啟明(1982-),男,湖南衡陽人,湖南大學博士,湖南農業大學講師
?通訊聯系人,E-mail:sw_x lm@hnu.cn
