衡陽盆地紫色土丘陵坡地主要植物群落自然恢復演替進程中種群生態位動態
2010-05-07 11:11:46鄒冬生李建國陳盛彬陳志陽
水土保持通報 2010年4期
楊 寧,鄒冬生,李建國,陳盛彬,陳志陽
(1.湖南農業大學生命科學學院,湖南長沙410128;2.湖南環境生物職業技術學院園林系,湖南衡陽421005)
生態位(niche)又稱生態龕,最早是由美國生態學家Grinnell于1917年給予定義:恰好被一種或一個亞種所占據的最后單元[1]。早期生態位的研究多集中于動物學領域,至20世紀70年代末才被廣泛運用于植物學的群落結構、種間關系、生物多樣性、物種進化等方面,成為解釋自然群落中種間共存與競爭機制的基本理論之一[2-3]。然而生態位又是極其抽象模糊的概念,一些學者曾就某一側面進行了空間生態位、營養生態位和功能生態位等的定義。而Hutchinson把生態位看作一個生物單元生存條件的總體集合,將其拓展為既包括生物的空間位置及其在生物群落中的功能地位,又包括生物在環境中的位置,即所謂的“n維超體積生態位”[4],這樣不僅豐富了生態位的概念與內涵,而且為生態位的定量計測提供了可能。尤其近年借助于一些數學方法,實施有效降維,使得生態位的理論與計測均出現了新的飛躍[5-6]。目前關于植物群落生態位的研究,大多側重于對某一群落主要物種或優勢物種的研究[7-9],或對不同立地條件下群落生態位的比較研究[10-11]。對于動態研究,如處于演替過程中的群落種群生態位動態研究極少,僅是沙地環境下植被次生演替過程中的種群生態位的研究[12]。而對于衡陽盆地紫色土丘陵坡地的植物群落的恢復演替進程中種群生態位的動態變化還未見報到。
以衡陽盆地紫色土丘陵坡地為代表的紫色土地區被認為是湖南省環境最為惡劣的地區之一,植被恢復重建難度較大,長期以來,該區域的植被恢復倍受重視。由于紫色土有機質含量和氮的含量較低,滲透性較差,加上紫色土顏色深吸熱性強,蒸發量大,以及區域內水、熱等不利因素的影響,致使紫色土丘崗地區水土流失嚴重,植被稀疏,基巖裸露,有的區域幾乎沒有土壤發育層,生態環境十分惡劣,植被恢復十分困難[13]。為此,本研究通過群落調查,從生態位的角度分析群落演替過程中的結構變化、種群動態、種間關系以及物種的環境適應等特性,旨在為衡陽盆地紫色土丘陵坡地的生態恢復重建提供指導和借鑒。
1 研究區概況及研究方法
1.1 研究區概況
衡陽盆地位于湖南省中南部,湘江中游,位于東徑110°32′16″—113°16′32″,北緯 26°07′05″—27°28′24″,地處湖南省凹形面的軸帶部分,周圍環繞著古老宕層形成的斷續環帶的嶺脊山地,構成典型的盆地地形。
該區屬亞熱帶季風濕潤氣候,年平均氣溫18℃;極端最高氣溫40.5℃,極端最低氣溫-7.9℃,年平均降雨量1 325 mm,年平均蒸發量1 426.5 mm。平均相對濕度80%,全年無霜期286 d。地貌類型以丘崗為主。
衡陽盆地紫色土面積有1.625×105hm2,呈網狀集中分布于衡陽盆地中部海拔60~200 m的地帶,東起衡東縣霞流、大浦,西至祁東縣過水坪,北至衡陽縣演陂、渣江,南達常寧市官嶺、東山和耒陽市遙田、市爐一帶,以衡南、衡陽兩縣面積最大。
1.2 研究方法
1.2.1 樣地的選擇及群落調查 于2007年4月下旬至5月下旬,結合當地的記載資料,采用“空間序列代替時間序列”的方法進行群落階段的劃分,群落演替的初始條件均為撂荒地,選取4類代表不同演替階段的樣地(表1),進行樣方調查,選取的樣方面積為20 m×20 m,在每個樣方內的四角和中心分別設置5個4 m×4 m和1 m×1 m的小樣方來調查灌木植物和草本植物,并記錄樣方內物種的種類、頻度、均高、蓋度以及密度,計算其重要值。每類樣地設樣方50個,其取樣方200個。調查結果取均值后,進行生態位分析。

表1 各類樣地狀況
1.2.2 分析方法 將群落調查的每個樣方視為多種資源的綜合狀態,這樣樣方數即為資源梯度的數目。以種群重要值指標計測其生態位寬度與之間的生態位重疊,其中,重要值VI=(相對蓋度+相對密度+相對高度+相對頻度)/4。選用重要值指標是因為它是多項指標的綜合,既能較好地體現物種對環境資源的利用效率,又可避免因各物種個體大小差異所帶來的誤差[10]。
生態位寬度是物種對環境資源利用多樣性的一種測度,反映不同物種的生態適應幅度,也可表示物種利用或趨于利用所有可利用資源狀態而減少種內個體相遇的程度[14],或生態轉化性的倒數[15]。此測算方法也較多,主要有 Levins生態位寬度、Simpson生態位寬度以及Shannon—Wiener生態位寬度,效果均較好。本研究采用廣為應用的Levins生態位寬度[16],測算公式為:
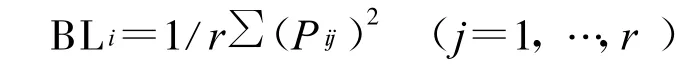
式中 :BLi——物種i的 Levins生態位寬度;Pij——物種i對第j資源梯度級的利用占它對全部資源利用的百分率,Pij=nij/Ni,而 Ni=∑nij,nij— —物種i在資源梯度級j的數量特征值(如蓋度、重要值、密度等),本研究為種 i在第j樣方的重要值;r——資源等級數,本研究為樣方數。
將演替系列看作各演替階段和不同演替時間群落類型組成的多維空間,則種群在整個演替系列的Levins生態位總寬度BLt[17]。
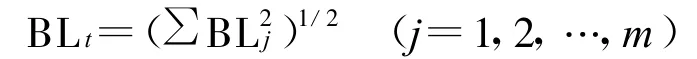
式中:BLj——表示植物種群在第 j個演替階段或第j個群落類型的Levins生態位寬度;m——組成演替系列的群落演替階段或群落類型的數目。
生態位重疊為物種在其與生態因子聯系上的相似性,反映不同物種對生態條件要求的相似程度[5]。植物群落研究中,生態位重疊是種群對相同資源的共同利用,或是共有的生態空間資源區域[18]。采用的測算方法主要有Pianka生態位重疊和Petraitis生態位重疊等。本研究采用Pianka生態位重疊[19],該指數簡單、對稱,能明確反映物種在資源利用上的重疊,測算公式為:

式中:Oik——物種i和物種k的生態位重疊值;nij,nkj——種i和種k在資源梯度級j的數量特征,本研究中種i和種k在樣方中的重要值;r——樣方數。
2 結果與分析
2.1 生態位寬度
通過調查與測算,計算出了衡陽盆地紫色土丘陵坡地植物群落演替不同階段內物種及其生態位寬度以及每一種群在整個演替系列中生態位總寬度(表2)。
由表2可知,演替初期(Ⅰ),由于其生境惡劣(干旱、缺水,土層淺薄,基巖顯露),物種極其稀少。以適合于繁殖力強、抗干旱與瘠薄能力強的狗尾草的生態位最高并占有絕對優勢,為群落的優勢種群;當演替進入中前期Ⅱ時,一些新物種隨生境改善而侵入,并占據一定的生態位,狗尾草種群也進一步擴展,其生態位寬度達到演替系列中的最高值,仍為群落的單優種群,整個群落對資源的利用也有所提高。
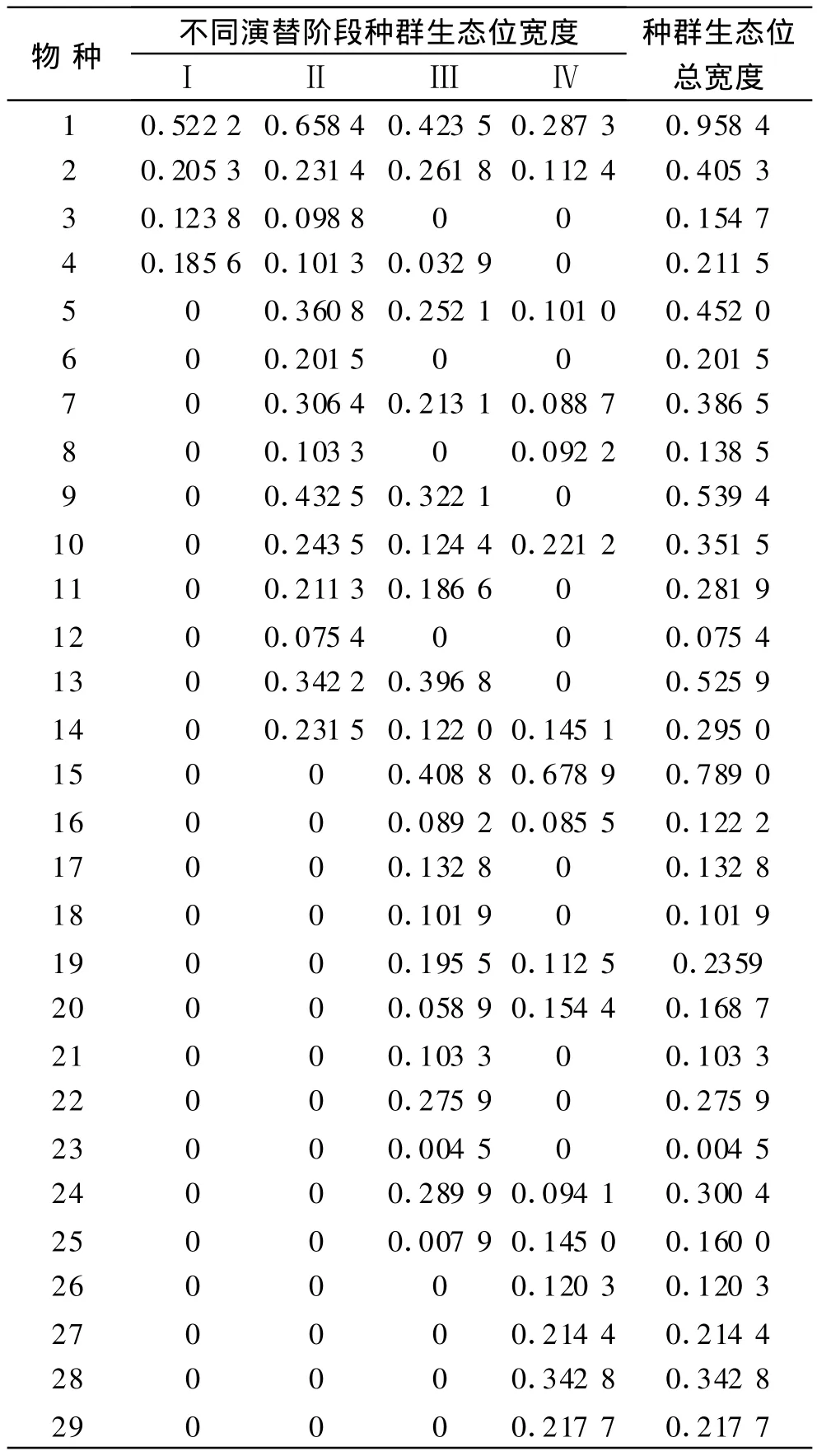
表2 衡陽盆地紫色土丘陵坡地不同演替階段種群生態位寬度
當演替進入中后期Ⅲ時,物種數達到最高,對資源的需求最強,種間競爭加劇,引起各種群生態位的進一步變化。盡管此時狗尾草的生態位仍最高,但已明顯收縮,已與牡荊和一支黃花(Solidago canadensis)等種群的生態位相當,其在群落中的絕對優勢地位喪失,群落也由以前的單優種群落演化為共優種群落;進入演替后期Ⅳ,物種數稍有回落,狗尾草生態位仍在進一步縮小。
相反,牡荊生態位進一步擴展,最終占據絕對優勢,群落的優勢種再一次轉化為單優種群,完成了優勢種的更迭,群落類型也隨之改變,由原來的狗尾草群落演化為牡荊群落。
在群落演替的過程中,隨著生境條件的改變,一些物種的生態位寬度值也隨之發生改變,從而導致了它們在整個演替過程中的興衰。
2.2 生態位重疊
群落中復雜的生態關系使各種群的生態位通常不是表現為離散的,而總是傾向于分享其它種群的基礎生態位部分,結果導致兩個或更多的種群對某些資源的共同需求,使不同種群的生態位處于不同程度的重疊狀態[2]。衡陽盆地紫色土丘陵坡地植物群落演替不同階段內各種群間生態位重疊及不同階段群落生態位重疊總均值見表3—7。
演替初期(Ⅰ),盡管生態環境惡劣,但物種稀少,資源相對充分,各種群可相對獨立地享用環境資源,因而生態位重疊相對較小。其中狗尾草與須芒草兩物種重疊在群落內最高,表明它們在同一資源位上出現的頻率較多,生態相似性較大。而馬鞭草與野菊花盡管生態位的寬度最低,但其生態位重疊值處于第二位,在野外的群落調查中,發現它們常有同時出現的現象。因此,并非生態位重疊大的種群其生態位寬度值就一定高。以上這些特點說明種群在適應和利用生態位資源方面具有趨同式分離特性。
演替中前期(Ⅱ),種群間的生態位重疊分離明顯,如狗尾草與夏枯草、須芒草、一支黃花、蒲公英等種群間生態位重疊較高,這是因為優勢種狗尾草與這些種群均具有較寬的生態位,在群落的分布上較多地重疊與混生;而另有某些種群如野菊花,本身生態位較窄,與其它多數種群間的生態位重疊極低,甚至為0,表明這些物種在此時的生境里生態幅窄小,對資源利用能力弱小。
演替進入物種最為豐富的中后期(Ⅲ),生態位重疊分離現象更為明顯。此時,狗尾草與牡荊構成群落的共優群落,整個群落內優勢種群間、優勢種群與其它種群間以及其它種群間大多生態位重疊均較高,特別是優勢種狗尾草與牡荊間的生態位重疊值高達0.74,表明群落中的大多數種群在資源利用上具有極大的相似性。重疊增高必將引起種群間競爭加劇,當可利用資源不足時,競爭必將更加激烈,使得一些種群生態位降低,與其它種群間的生態位重疊進一步降低。
演替進入后期(Ⅳ),作為單優種群的牡荊,盡管生態位較寬,但與大多數種群間生態位重疊的并不高,這可能是牡荊強大有競爭力排斥其它物種與其共生、分享資源的原故,它與狗尾草的生態位重疊值有所降低,但仍有0.60,這是因為它們二者具有利用資源上的相似性。另外,與演替中后期(Ⅲ)相比,由于生境條件與群落組成結構的改變,狗尾草與別的植物的生態位重疊出現了升降有別的不同變化趨勢。
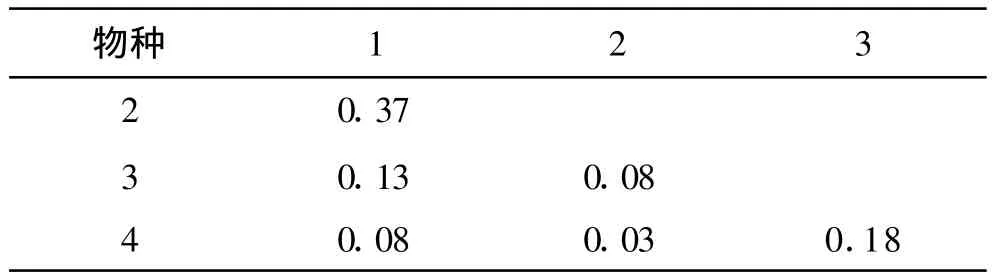
表3 衡陽盆地紫色土丘陵坡地演替初期(Ⅰ)種群間生態位重疊
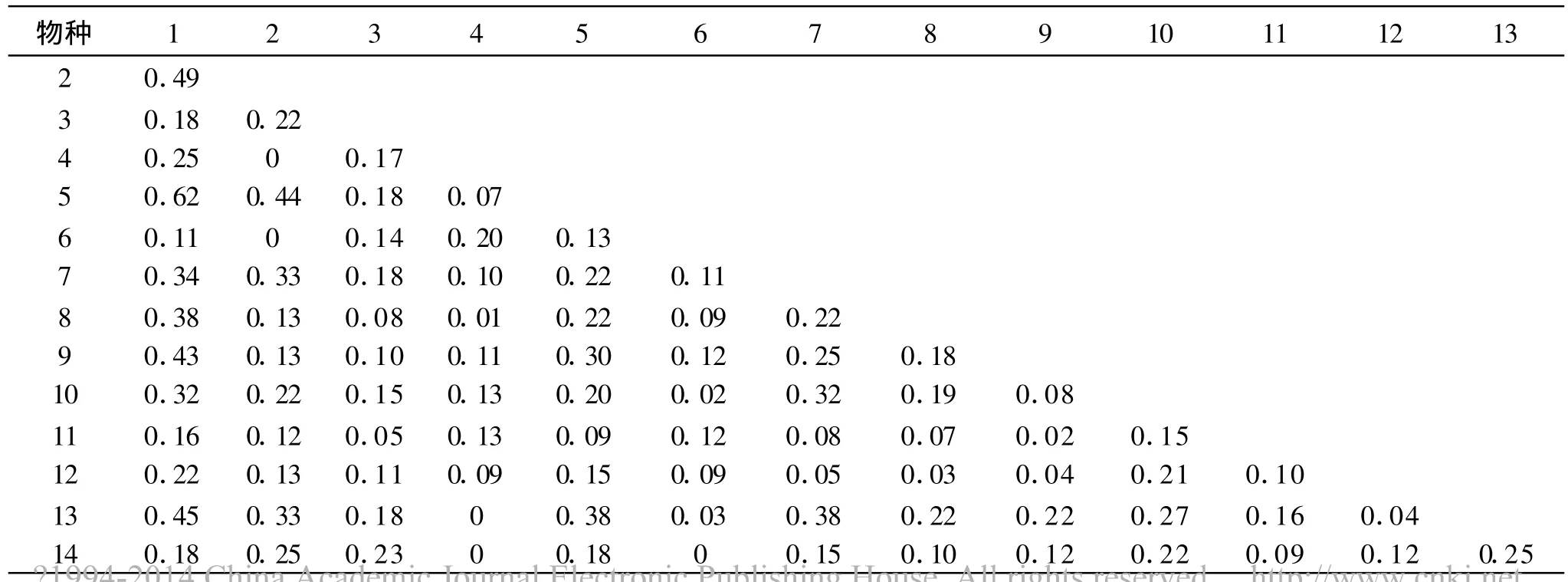
表4 衡陽盆地紫色土丘陵坡地演替中前期(Ⅱ)種群間生態位重疊
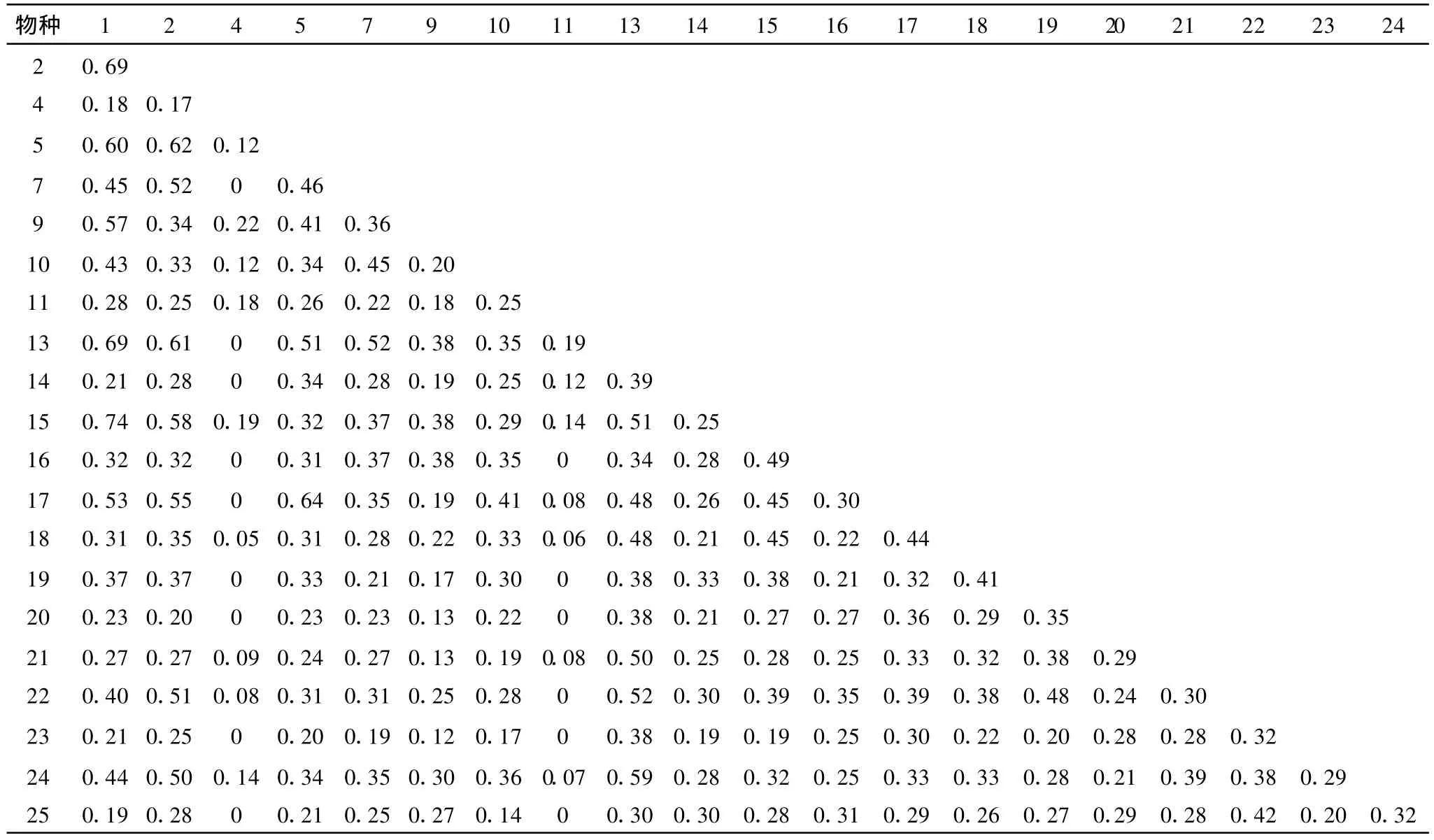
表5 衡陽盆地紫色土丘陵坡地演替中后期(Ⅲ)種群間生態位重疊

表6 衡陽盆地紫色土丘陵坡地演替后期(Ⅳ)種群間生態位重疊
由表7可知,不同階段群落的生態位重疊平均值前期隨演替逐漸增加,中后期(Ⅲ)達最大,后期(Ⅳ)稍有回落,這是群落內種內競爭和種間競爭共同作用的結果。
結合表2—6可知,演替初期,群落內物種較少,植物平均占有的資源相對豐富(特別是光資源),各物種均能獨立地占據各自的生態位,使之表現為分離狀態,因而群落總體平均生態位重疊較小,此時群落內的競爭更多地表現為種內競爭,使各種群生態位呈擴張趨勢,加之新物種隨著群落的演替而不斷侵入,占據并拓展各自生態位。這樣,在有限的生境容量下,種群間必將出現較多的生態位重疊,致使平均生態位重疊上升。因此,至群落物種最豐的演替中后期,平均生態位重疊最大,群落內的競爭也最為激烈。而此時的競爭則更多地表現為種間競爭,激烈的競爭既淘汰一些物種,又對存留物種的生態位進行優化,使之分離。因此,演替后期群落的平均生態位重疊出現回落,競爭也有所減弱,各物種從而能較為協調、充分地分享環境資源,群落趨于平衡和穩定,這也是群落演替趨于成熟的特征之一。事實上,群落演替過程中,平均生態位重疊呈“小—大—小”的動態變化,正是生態學中的“種內競爭促使不同物種的生態位重疊,種間競爭促使不同物種的生態位分離”的理論在群落演替中的體現[2]。

表7 研究區不同演替階段群落內種群間生態位重疊均值
3 結論與討論
(1)生態位越寬,物種的特化程度越低,該物種越傾向于一種泛化種;相反,物種的生態位越窄,該物種特化度越高,更傾向于一種特化種。通常情況下,泛化種較特化種具有更強的競爭力,當可利用資源不足時,這種競爭優勢更為明顯。然而泛化種和特化種的劃分并非絕對的,二者隨環境的變化可以發生轉換。在群落演替過程中,可根據種群生態位大小及其動態變化來確定泛化種與特化種以及它們在不同演替階段的轉化。如夏枯草等由演化中前期的泛化種轉化為后期的特化種,女貞等由演替中后期的特化種轉化為后期的泛化種。
在群落演替種群不斷更替的過程中,優勢種群的更替及其生態位變化更為重要。衡陽盆地紫色土丘陵坡地是一個開放的生態系統,自然的風力、動物與人類的活動為多種植物的侵入提供了載體和媒介,然而惡劣的初始環境(干旱、缺水;土層淺薄,基巖顯露),對侵入植物進行了嚴格的篩選。只有那些生態位較寬、適應力較強的先鋒種才能成功定居,繼而開始群落的演替。而狗尾草下正是憑借其特有的生態學特征成為群落的先鋒種群乃至優勢種群。它耐瘠薄、耐干旱、繁殖力強,較好地適應了演替初期的生態環境,成為群落的優勢種群。隨著演替的進行,狗尾草生長、繁殖,占據空間越來越大,形成草丘并連成片,至演替的中前期,發育為成熟的狗尾草群落。此時,狗尾草群落在整個演替階段中生態位最高,為群落的單優種群。隨著演替的進一步推進,其生態環境有所改善,另一種植物牡荊進入群落,其強大的繁殖力、競爭力和資源利用能力使其很快占有較寬的生態位,與狗尾草一起構成群落的共優群落。至演替的后期,環境的改變使得狗尾草的生態位進一步降低,牡荊的生態位卻持續擴張,成為群落的單優種群。而其它一些物種在演替過程中是否出現及出現量的多少也是它們各自生態位變化的結果。
(2)研究衡陽盆地紫色土丘陵坡地主要植物群落自然恢復演替進程中種群生態位動態,將其植物群落劃分為4個不同的演替階段,其中,與演替中后期(Ⅲ)對應的牡荊+狗尾草群落物種最豐,生態位重疊最高。事實上,衡陽盆地紫色土丘陵坡地植物群落自然演替過程中的植被類型并不復雜,主要是狗尾草群落和牡荊群落,而牡荊+狗尾草群落只是狗尾草群落和牡荊群落的過渡交錯群落,正是交錯帶特有的生態特性,使該階段物種最多,生態位重疊最高,但它只是群落演替過程中一個短暫的、不穩定的過渡階段,激烈的種間競爭必將使種群生態位出現分離,從而使群落進入一個生態位重疊度降低的穩定群落階段。
(3)生態位的重疊是兩個或更多的物種對相同資源的共同利用,研究種群生態位重疊隨群落演替的變化,可以揭示群落演替過程中種間關系的變化。對生態位重疊大小的解釋往往與種間競爭相聯系。通常認為種間競爭越激烈,生態位重疊值越大。但生態位重疊絕不能與競爭程度等同,還要考慮資源供應狀況和對資源利用的互補性,群落資源豐富時,生態位重疊值大,說明二者共享同一資源,競爭并不激烈。事實上,生態位高度重疊既有可能是物種間有相近的生態特征,也有可能是其對生態因子有互補性要求,甚至二者兼有。相近生態特征的物種,只有在共享資源不足的前提下才產生競爭。如共享資源豐富時,生態位重疊度就不能反映競爭的程度,而只表明兩個種群因生態相似性而占據了相近的生態空間[19-20]。衡陽盆地紫色土丘陵坡地優勢種狗尾草與牡荊的生態特征的相似性,因而在利用資源也具有相似性,當共享資源不足時必將表現出激烈的競爭。然而二者又具有生態特征的差異性,使得它們可以互補性地利用生境因子。
因此,二者生態位高度重疊既有生態特征相似性的原因,也有對生境因子互補性要求的原因,從而使得二者既表現競爭又表現共生的特性,這也可解釋牡荊種群替代狗尾草種群的長期性和不徹底性。因此,對生態位重疊值的解釋要結合群落的資源狀態和群落演替過程加以綜合考慮。
(4)在衡陽盆地紫色土丘陵坡地植被恢復的措施上,應重視進行保護,不同的人為干擾程度,使得植被受到一定的破壞,因此,在人為干擾比較嚴重、生境條件相對有利的區域應實行全面封禁,杜絕人為破壞,使之進行自然演替和恢復。在生境條件相對較差的區域,應堅持因地制宜,用科學的方法進行人工恢復植被,加速其演替過程。
[1] Grinnel J.Geography and evolution[J].Ecology,1924,5:225-229.
[2] 楊寧,鄒冬生,李建國.衡陽盆地紫色土丘陵坡地自然恢復灌叢階段主要種群空間分布格局[J].生態環境學報,2009,18(3):996-1001.
[3] 楊寧,鄒冬生,李建國.衡陽盆地紫色土丘陵坡地植物群落數量分類及物種多樣性研究[J].農業現代化研究,2009,30(5):615-619.
[4] Hutchinson G E.Concluding remarks[J].Cold Spring Harbor,Symp.Quant.Biol.,1957 ,22:415-427.
[5] 王剛,趙松嶺,張鵬云,等.關于生態位定義的探討及生態位重疊計算公式改進的研究[J].生態學報,1984,4(2):119-127.
[6] 黃英姿.生態位理論研究中的數學方法[J].應用生態學報,1994,5(3):331-337.
[7] 譚永欽,張國安,郭爾祥.草坪雜草生態位研究[J].生態學報,2004,24(6):1300-1305.
[8] 鄭元潤.大青溝森林植物群落主要木本植物生態位研究[J].植物生態學報 ,1999 ,23(5):475-479.
[9] 李軍玲,張金屯,郭逍宇.關帝山亞高山灌叢草甸群落優勢種群的生態位研究[J].西北植物學報,2003,23(12):2081-2088.
[10] 胡相明,程積民,萬惠娥,等.黃土丘陵區不同立地條件下植物種群生態位研究[J].草業學報,2006,15(1):29-35.
[11] 劉加珍,陳亞寧,張元明.塔里木河中游植物種群在四種環境梯度上的生態位特征[J].應用生態學報,2004,14(4):549-555.
[12] 張繼義,趙哈林,張銅會,等.科爾沁沙地植物群落恢復演替系列種群生態位動態特征[J].生態學報,2003,23(12):2741-2746.
[13] 楊寧,鄒冬生,李建國.衡陽盆地紫色土丘陵坡地土壤水分變化動態研究[J].水土保持研究,2009,16(6):16-21.
[14] Hurlbert S.The measurement of niche overlap and some relatives[J].Ecology,1968 ,59(1):67-77.
[15] Klopfer P J M.Nichesize and faunal diversity[J].Am.Nat.,1960 ,94:29-300.
[16] Levins R.Evolution in changing environments:Some Theoretical explorations[M].Princeton:Princeton U-niversity Press,1968.
[17] Li D Z,Qin A L,Zang R G.Measure and analysis of dominant wood populations niche in natural secondary forest in east mountains of northest China[J].Quarterly Journal of Chinese Forestry ,1995,28(2):3-12.
[18] Abrams R.Some comments on measuring niche overlap[J].Ecology ,1980,61:44-49.
[19] Pianka E R.The structure of lizard communities[J].Annual Review of Ecology&Systematics,1973 ,4:53-57.
[20] 楊效文,馬繼盛.生態位有關術語的定義及計算公式評述[J].生態學雜志,1992,11(2):44-49.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
保健醫苑(2021年7期)2021-08-13 08:48:02
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
汽車觀察(2018年10期)2018-11-06 07:05:26
資源再生(2017年3期)2017-06-01 12:20:59
