4種結縷草光合特性及葉綠素含量的比較研究
2010-05-13 08:10:52李高揚李建龍
草原與草坪 2010年1期
鄧 蕾,李高揚,王 艷,李建龍,姜 濤
(1.南京大學生命科學院草學中心,江蘇南京 210093;2.連云港曲情生物科學技術有限公司,江蘇連云港 222000)
草坪具有凈化空氣、調節空氣溫度、消減噪音、保持水土、美化環境的功能,在城市生態系統中起著重要作用。由于園林綠化廣泛使用的草坪草大多具有不耐蔭、適應性差等特性,在高大喬灌木或大型建筑物群等弱光照環境條件下生長較差,而且在冬季弱光條件下枯黃期長,生長緩慢。為了解決處于植物群落最下層草坪草的適應性問題,篩選出質優、適應性強的耐蔭性草坪草已成為目前重要的研究課題。
結縷草屬(Zoysia)植物在我國分布廣泛,遺傳多樣性豐富,而且具有低維護、耐鹽堿、耐干旱、耐踐踏等特點,成為重要的草坪草種質[1]。有關結縷草的特性研究集中在抗寒性、抗旱性和抗鹽性等方面[2-6]。但是結縷草在弱光蔭蔽環境的生理生態表現的報道很少。
以結縷草不同種為試驗材料,測定并分析了光合作用中的幾個重要參數以及葉綠素含量,探討結縷草屬草坪草的光適應性以及對弱光的捕捉利用能力,為結縷草在弱光蔭蔽條件下的推廣和應用以及在室內弱光下生長的微型草坪[7]的選種提供理論依據。
1 材料和方法
1.1 試驗地概況
田間試驗在南京大學浦口校區試驗基地進行。南京地處長江中下游平原,N 31°14′~ 32°37′,E 118°22′~119°14′,南京屬北亞熱帶季風氣候區,四季分明 ,雨水充沛,光能資源充足,年平均溫為15.7℃,最高氣溫43℃,最低氣溫-16.9℃,最熱月平均溫度28.1℃,最冷月平均溫度 -2.1℃。年均降水117 d,降水量為1 106.5 mm,最大平均濕度 81%。最大風速19.8 m/s。土壤最大凍結深度0.09 m。夏季主導風向東南、東風,冬季主導風向東北、東風。無霜期237 d。每年6月下旬到7月中旬為梅雨季節。
1.2 試驗材料
試驗用溝葉結縷草(Zoysia matrella)原始材料來自浙江麗水學院,中華結縷草(Zoysia sinica)、日本結縷草(Zoysia japonica)和細葉結縷草(Zoysia tenui folia)來自南京農業大學草坪實驗地。于2006年7月下旬在南京大學浦口校區草坪基地進行栽種,2007年5月上旬重新移栽種植。
1.3 試驗方法
1.3.1 葉綠素含量測定 采用SPAD-502葉綠素儀對4種結縷草葉片進行測定,測量葉片在兩種波長(650nm和940 nm)的光學濃度差來確定葉片當前葉綠素的相對數量。每個結縷草種重復測定15次,最后取其平均值。
1.3.2 光合參數的測定 野外試驗于2007年9月16日至17日進行,2日均為晴朗天氣,全天平均溫度為25℃,午間可達30℃。每天測量時間為上午8:00時至下午18:00。測量時分別選取草坪叢頂端以下第3~5片完整展開葉進行光合參數及光響應曲線的測定,測定3次[8]。
光合參數的測定使用 LT-6400便攜式光合作用系統 ,開放式氣路,空氣流速為500 μ mol/s。光合生理參數及環境參數的測定包括凈光合速率(Pn)、蒸騰速率(Tr)、光合有效輻射(PAR)、空氣溫度(Ta)、空氣CO2濃度(Ca)、大氣相對濕度(R H)等。
光響應曲線的測定使用Li-6400-02B內置式紅藍光源。測量時間 9:00至11:30,光強為2 000、1 500、1 000、500、200、100、50、20 和0 μ mol/(m2? s),測定時每一光強下停留200 s,測量前使用人工光源進行5~10 min的光合誘導。
1.4 統計分析
采用Excel對數據進行處理和應用SPSS分析和擬合光響應曲線。將獲得的不同光強梯度的凈光合速率根據Prioul及Chartier[9]的模型進行非線性回歸擬合得到Pn-PAR曲線,具體模型為:
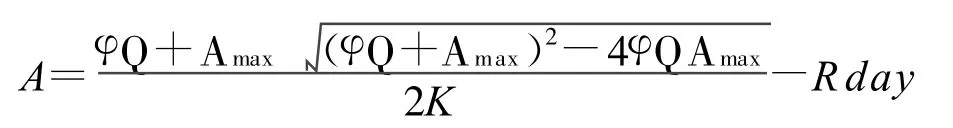
其中A:凈光合速率(Pn),Amax:最大凈光合速率(Pnmax),φ:表觀量子效率,k:曲角,Rday:暗呼吸速率 ,Q:光合有效輻射(PAR)。由該模型計算出最大光合速率(Pnmax)、表觀量子效率(AQY)與暗呼吸速率(Rday),并根據擬合后的光合曲線方程計算出光補償點(LCP)、光飽和點(LSP)[10]。
2 結果與分析
2.1 結縷草不同種葉片的葉綠素含量測定
葉綠素是植物進行光合作用的催化劑、植物綠色的源泉,草坪草品種不同,葉片葉綠素含量有差異,綠度也有所不同,葉綠素含量越高,顏色越深[11]。葉綠素含量的高低是反映植物光合能力的重要指標之一。日本結縷草葉綠素含量最大,達到34.5 mg/g;其次為中華結縷草和溝葉結縷草,為32.7和30.5 mg/g;細葉結縷草最小,為22.7 mg/g(圖1),此值分別比日本結縷草低11.63%,5.25%和34.09%。葉綠素含量在日本結縷草,溝葉結縷草和中華結縷草之間沒有顯著差異(P>0.05),但這3個品種與細葉結縷草之間卻有顯著差異(P<0.05)。

圖1 結縷草不同種葉片葉綠素含量Fig.1 The chlorophyll contents of leaf of 4 Zoysia species
葉綠素含量的高低代表了葉片捕捉和利用光能的能力,同時也影響光能在葉綠體中的分配。日本結縷草和中華結縷草的葉綠素含量較高,因此它們對光能的利用能力以及對弱光的捕捉能力更強;溝葉結縷草稍差一些,細葉結縷草利用光能的能力最差。
2.2 結縷草不同種葉片的實測凈同化速率
植物的光合作用經常受到外界環境條件和內部因素的影響而發生變化,凈同化速率是表示光合作用變化的指標之一。實測凈同化速率反映了測量瞬間各種參數的實際狀況,為擬合值提供了基礎與參考。4種結縷草的凈同化速率隨著光合有效輻射的增強出現了先升后降的變化,日本結縷草,中華結縷草和溝葉結縷草的峰值均在光合有效輻射為1 000 μ mol/(m2?s)(圖2)出現,繼續增強光合有效輻射,反而會降低凈同化速率。說明過強的光合有效輻射會抑制光合作用。而細葉結縷草在200 μ mol/(m2?s)時出現最大凈同化速率,而且其值始終低于其他3種結縷草。
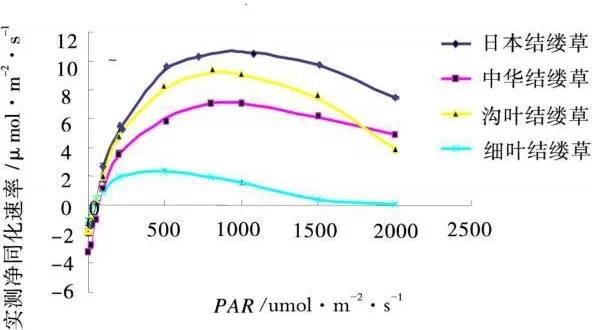
圖2 結縷草不同種葉片的實測凈同化速率Fig.2 The net assimilation rate in leaf of 4 Zoysia species
2.3 不同結縷草品種葉片的擬合凈同化速率
擬合凈同化速率可排除測量的隨機性以及環境、人為等因素,以獲得較為穩定的數據。同時通過數據擬合,計算出Pnmax、AQY與Rday,并根據擬合后的光合曲線方程計算出LCP、LSP。
植物LCP和LSP的高低直接反映了植物對弱光的利用能力,是植物耐蔭性評價的重要指標。表1表明,LSP在日本結縷草,溝葉結縷草和中華結縷草之間沒有顯著差異(P>0.05),但這3個品種與細葉結縷草之間卻有顯著差異(P<0.05),并且細葉結縷草LSP較低;而LCP在日本結縷草和溝葉結縷草之間沒有顯著差異(P>0.05),在中華結縷草和細葉結縷草之間也沒有顯著差異(P>0.05),但是中華結縷草和細葉結縷草卻顯著高于日本結縷草和溝葉結縷草,這種差異可能與結縷草的不同起源有關。
Rday是指黑暗中進行的氣體交換速率,較低的暗呼吸速率可減少呼吸作用對光合產物的消耗[12]。中華結縷草的暗呼吸速率為3.222 μ mol/(m2?s),顯著高于其他3種結縷草(P<0.05),而溝葉結縷草和日本結縷草之間無顯著差異(P<0.05),細葉結縷草最小,為1.00 μ mol/(m2?s),顯著低于其他3種結縷草(P<0.05)。反映植物對低光利用效率的表觀量子效率(AQY)在日本結縷草,溝葉結縷草和細葉結縷草之間沒有顯著差異(P>0.05),但是中華結縷草顯著高于其他3種結縷草(P<0.05)。
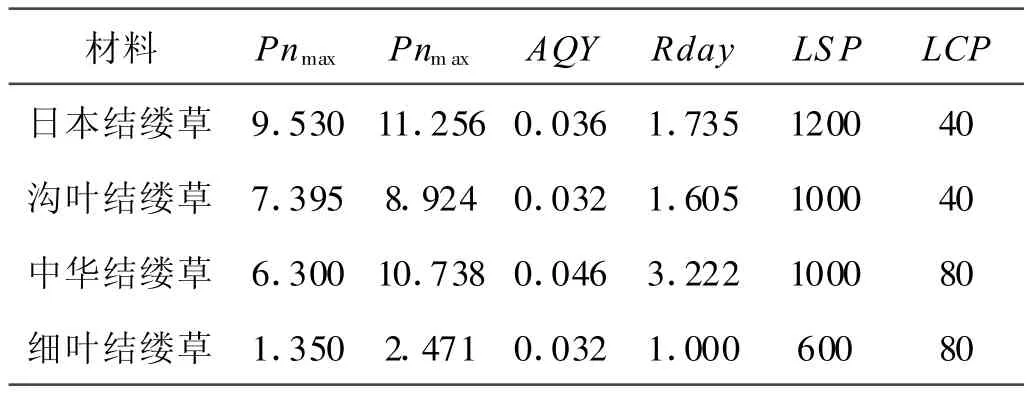
表1 結縷草不同種葉片的擬合凈同化速率Table 1 The estimated net assimilation rate of leaf of 4 Zoysia species μ mol/(m2?s)
2.4 結縷草不同種葉片的光合特征分析
光合作用是植物形成生物產量的生理代謝基礎。光合速率是表示光合作用變化的另一指標。Pn-PAR曲線(圖 2)表明,4種結縷草的最大凈光合速率(Pnmax)從高到低依次為日本結縷草,溝葉結縷草,中華結縷草,細葉結縷草。數值為:日本結縷草 9.53 μ mol/(m2?s),溝葉結縷草7.395,中華結縷草6.30 μ mol/(m2?s),細葉結縷草 1.35 μ mol/(m2?s)。分別為日本結縷草的77.60%,66.11%和14.17%,相互間表現為差異顯著(P<0.05)。而凈光合速率的變化在光強為 0~500 μ mol/(m2?s)增加最快,之后不再增加。這可能是由于過強的光合輻射使得植株葉片的氣孔關閉,從而使其利用光能的效率下降。

圖3 不同結縷草品種葉片光合-光強(Pn-PAR)響應曲線Fig.3 The Pn-PAR response curve of 4 Zoysia species
3 結論與討論
光合作用是植物生長發育的基礎,而光照可以提供作物同化力形成所需要的能量,活化參與光合作用的關鍵酶,促進氣孔開放。植物光合作用能力的強弱與植物的遺傳性和生長環境關系密切,光合作用的某些生理參數,如 Pmax、LSP、LCP等,已成為植物速生豐產以及制定栽培措施的科學依據。
一般認為在弱光條件下,單位葉面積的葉綠體數目減少,單位重量葉綠素含量增加,從而可以提高植物對于弱光的利用率。在試驗中,日本結縷草的葉綠素含量最大,其次為中華結縷草,溝葉結縷草,而細葉結縷草葉片葉綠素含量最小。不同種結縷草葉綠素含量的差異表明,日本結縷草和中華結縷草具有更好的對遮蔭環境的適應力。
供試4種結縷草的光合作用差異顯著,LSP在日本結縷草、溝葉結縷草和中華結縷草之間沒有顯著差異(P>0.05),但這3個種與細葉結縷草之間卻有顯著差異(P<0.05),并且細葉結縷草光飽和點較低,這可能跟細葉結縷草葉片葉綠素含量少有關。日本結縷草和溝葉結縷草的LCP均為40 μ mol/(m2?s),接近于自然光照陽性植物的LCP[30 μ mol/(m2?s)][13];中華結縷草和細葉結縷草的 LCP較高,平均為80 μ mol/(m2?s),LSP 平均為 1 200 μ mol/(m2?s)。4種結縷草的 AQY與自然光照條件下植物的AQY 0.03~0.06 mol/moL一致[14]。但是中華結縷草的AQY顯著高于其他3種結縷草。
綜上所述,可初步推斷出結縷草為陽性、且耐蔭性較強的草坪草。這與在實際生產實踐中結縷草的耐蔭性表現一致,尤其是中華結縷草,耐蔭性強,對弱光的捕捉能力也更強,因此在弱光蔭蔽環境下生長較旺盛,顏色鮮艷,青綠期長,適于亞熱帶地區的栽植建坪。
[1] 王文強,李志丹,白昌軍,等.結縷草屬種質資源及其應用研究進展[J].草原與草坪,2006(2):3-6.
[2] 黃錦文,陳冬梅,鄭紅艷,等.暖季型結縷草對低溫響應的生理生態特性[J].中國草地學報,2009,31(1):64-69.
[3] 胡化廣,張振銘,黃亮亮,等.結縷草對干旱脅迫的響應[J].草原與草坪,2008(3):41-44.
[4] 趙艷,孫吉雄,王兆龍,等.不同抗旱性結縷草品種在土壤干旱及復水過程中的生理差異[J].草原與草坪,2006(6):46-48.
[5] 王加真,李建龍,王艷,等.高鹽脅迫對溝葉結縷草葉片光合色素含量的影響[J].上海交通大學學報(農業科學版),2007,25(6):582-586.
[6] 杜永吉,于磊,魯為華,等.低溫脅迫下結縷草品種的抗寒性差異[J].中國草地學報,2008(5):116-120.
[7] 圖雅,李建龍,王艷,等.微型草坪對室內微環境的生態調節作用[J].草原與草坪,2008(4):59-62,66.
[8] 付為國,李萍萍,陳歆,等.北固山濕地虉草光合日變化的研究[J].武漢植物學研究,2005,23(6):555-559.
[9] Prioul J L,Chartier P.Partitioning of transfer and carboxylation components of intercellular resistance to photosynthetic CO2fixation:a critical analysis of the method used[J].Ann Bot,1977,41:789-800.
[10] Bassman J B,Zwier J C.Gas exchange characteristics of Populus trichocarpa,Populus deltoides and Populus trichocarpa × P.deltoides clones[J].Tree Physiol,1991,8:145-149.
[11] 李淑文,李迎春,彭玉信,等.不同草坪草葉綠素含量變化及其與綠度的關系[J].草原與草坪,2008(6):54-56,74.
[12] Kitao M ,Lei T T ,Koike T ,et al.Susceptibility to photoinhibition of three deciduous broadleaf tree species with different successional traits raised under various light regimes[J].Plant Cell&Environment,2000,23:81-89.
[13] Lancher W,T rans L B.Plant physioecology[M].Beijing:Science Press,1980.
[14] 邱國雄.植物光合作用的效率[M]//.余淑文.植物生理學和分子生物學.北京:科學出版社,1992:236-243.
