寄生性吸蟲谷胱甘肽過氧化物酶生物信息學(xué)分析*
2010-06-07 06:03:02蔡國(guó)斌蔣明森
中國(guó)人獸共患病學(xué)報(bào) 2010年4期
關(guān)鍵詞:分析
章 瑩,蔡國(guó)斌,何 立,蔣明森
谷胱甘肽過氧化物酶(glutathione peroxidase,GPx)是生物體內(nèi)廣泛存在的一種重要的過氧化物分解酶,以谷胱甘肽作為還原劑催化還原H2O2、有機(jī)氫過氧化物和脂氫過氧化物等活性氧分子(ROS)〔1〕,從而保護(hù)細(xì)胞膜的結(jié)構(gòu)及功能不受過氧化物的干擾及損害,對(duì)維持細(xì)胞膜結(jié)構(gòu)的完整性具有重要的意義。迄今發(fā)現(xiàn)GPx是一個(gè)至少包含6種不同同工酶(isoenzyme)的大型酶家族,包括經(jīng)典/胞漿型谷胱甘肽過氧化物酶(GPx1/c-GPx)、胃腸型谷胱甘肽過氧化物酶(GPx2/GPx-GI)、血漿型谷胱甘肽過氧化物酶(GPx3/p-GPx)、磷脂氫谷胱甘肽過氧化物酶(GPx4/PHGPx)、附睪特異性谷胱甘肽過氧化物酶(GPx5/e-GPx)和氣味代謝谷胱甘肽過氧化物酶(GPx6/o-GPx)。其中PHGPx被認(rèn)為是一種獨(dú)特的抗氧化酶,能直接保護(hù)生物膜免受過氧化損傷〔2〕。Cookson等(1992)和 Zelck 等(2004)認(rèn)為GPx在感染性寄生蟲中是組成酶防御系統(tǒng)的主要前線,以確保它們暴露在宿主代謝產(chǎn)生的有害ROS環(huán)境中得以生存〔3-4〕。
本文旨在通過網(wǎng)絡(luò)資源,搜尋到包括幾種重要醫(yī)學(xué)吸蟲在內(nèi)的不同生物GPx的cDNA、基因組DNA序列,然后通過生物信息學(xué)和比較基因組學(xué)方法,分析吸蟲GPx基因及其所編碼蛋白的分子特性、探討吸蟲GPx與其他生物體GPx的相關(guān)性,以及吸蟲GPx在生物系統(tǒng)發(fā)育中的進(jìn)化地位等。
1 材料與方法
1.1 醫(yī)學(xué)吸蟲GPx的cDNA(包括EST)和基因組DNA序列檢索 以華支睪吸蟲(Clonorchis sinensis)PHGPx1 cDNA序列(EF056481)作為查詢?cè)~(query)用于BLASTx,獲得從GenBank數(shù)據(jù)庫中已注冊(cè)的吸蟲GPx序列。然后,為了獲得更多常見寄生性吸蟲GPx的核酸序列,一些常用的寄生蟲學(xué)專業(yè)的基因組或EST數(shù)據(jù)庫用來作進(jìn)一步的檢索:如Sanger institute配備的數(shù)據(jù)庫網(wǎng)站http://www.sanger.ac.uk/Projects/Helminths可檢索曼氏血吸蟲(Schistosoma mansoni)、埃及血吸蟲(S.haematobium)、肝片形吸蟲(Fasciola hepatica)等的序列;上海生命科學(xué)和生物技術(shù)信息中心配備的日本血吸蟲數(shù)據(jù)庫(Shanghai Center for Life Science and Biotechnology Information,http://lifecenter.sgst.cn/sj.do)可檢索日本血吸蟲(S.japonicum)的序列等。
1.2 6大GPx家族DNA和氨基酸序列的檢索以華支睪吸蟲PHGPx1氨基酸序列(ABK58679)作為query進(jìn)行BLASTP,獲得從GenBank數(shù)據(jù)庫中的許多相近GPx序列。此外,人 c-GPx/GPx1(CAA68491)和p-GPx/GPx3(AAP50261)以及線蟲Brugia pahangi GPx3(CAA48882)也用來進(jìn)行BLASTP以獲得其他家族的GPx氨基酸和DNA序列。考慮到相似值和分類學(xué)上的分布,共107個(gè)成員(分別處在不同GPx家族和不同等級(jí)生物體)的氨基酸或cDNA序列用來作生物信息學(xué)分析。
1.3 一般生物信息學(xué)分析 cDNA序列所編碼的開放閱讀框(ORF)和同源性分析用美國(guó)生物技術(shù)信息中心(National Center for Biotechnology Information,NCBI:http://www.ncbi.nlm.nih.gov)網(wǎng)站下的ORF Finder和基本局部比對(duì)搜索工具(basic local alignment searvh tool,BLAST)進(jìn)行分析。使用SECISearch程序(版本2.19,http://genome.unl.edu/SECISearch.html)推測(cè) cDNA 3'-端非翻譯區(qū)(3'-UT R)是否含有硒半胱氨酸插入序列(selenocysteine insertion sequence,SECIS)。利用SignalP程序(http://www.cbs.dtu.dk/services/SignalP)推測(cè)所編碼的蛋白質(zhì)是否含有疏水信號(hào)肽,和用ScanSite pI/Mw程序(http://scansite.mit.edu/calc_mw_pi.html)計(jì)算理論分子量(Mr)和等電點(diǎn)值(pI)。
1.4 氨基酸序列比對(duì)(Amino acid sequence alignment)和系統(tǒng)進(jìn)化樹(Phylogenetic tree)分析 根據(jù)相似值和分類學(xué)上的分布,各家族成員的氨基酸先用ClustalX工具進(jìn)行對(duì)比,然后用GeneDoc程序優(yōu)化。首先對(duì)所有檢索到的醫(yī)學(xué)吸蟲GPx序列進(jìn)行比對(duì),并計(jì)算所檢測(cè)序列的相似值。然后對(duì)所有6大GPx家族成員進(jìn)行比對(duì),并用PHYLIP工具包(版本,ver 3.6b)中的 PROTPARS程序、或NEIGHBOR程序通過最大簡(jiǎn)約法則(maximum parsimony algorithm,MP)、或鄰近歸并聚類法則(又名鄰接法則,neighbor-joining algorithm,NJ)進(jìn)行系統(tǒng)學(xué)分析。系統(tǒng)樹用 TreeView顯示,每個(gè)分支上的數(shù)值用100個(gè)隨機(jī)樣品作為輸入序列、通過SEQBOOT程序算出有統(tǒng)計(jì)意義的數(shù)值。
1.5 比較基因組學(xué)分析 從基因組和/或EST數(shù)據(jù)庫,利用網(wǎng)絡(luò)資源檢索獲得各分類學(xué)上不同位置GPx代表成員的cDNA和基因組DNA,通過比較它們之間的堿基序列,得到各成員的基因結(jié)構(gòu):包括外顯子(exon)和內(nèi)含子(intron)的數(shù)量、外顯子和內(nèi)含子的序列長(zhǎng)度以及內(nèi)含子的剪接插入位置(intron phase)。然后進(jìn)行比較基因組學(xué)分析,探討醫(yī)學(xué)吸蟲GPx基因家族在分類學(xué)上的進(jìn)化關(guān)系。
2 結(jié) 果
2.1 醫(yī)學(xué)吸蟲GPx序列分析、及與其他GPx家族氨基酸比對(duì)分析 通過檢索,共獲得吸蟲 12個(gè)GPx序列,包括華支睪吸蟲(Cs)4個(gè)、衛(wèi)氏并殖吸蟲(P.westermani,Pw)2個(gè)、曼氏血吸蟲(Sm)2個(gè)、日本血吸蟲(Sj)2個(gè)、埃及血吸蟲(Sh)1個(gè)和肝片形吸蟲(Fh)1個(gè)。各序列分別設(shè)定為:CsGPx1(EF056481)、CsGPx2(EF056482)、CsGPx3(EF056483)、CsGPx4(EF056484)、PwGPx1(DQ454159)、PwGPx2(DQ454160)、SmGPx1(L37762)、SmGPx2(AY729668)、SjGPx1(CV689936)、SjGPx2(AY223160)、ShGPx1(Shaem32f07.q1k)和FhGPx1(Fhep45e11.q1k)。各基因編碼的氨基酸序列所預(yù)測(cè)的理論分子量均為20 kDa左右,部分基因N-末端含有疏水信號(hào)肽序列(圖1中5'-端底劃線處)。各氨基酸序列之間具很高的相似度,尤其在活性區(qū)高度保守。GeneDoc程序檢測(cè)的這些吸蟲GPx基因的氨基酸序列同一性值從35%到87%,如表1。
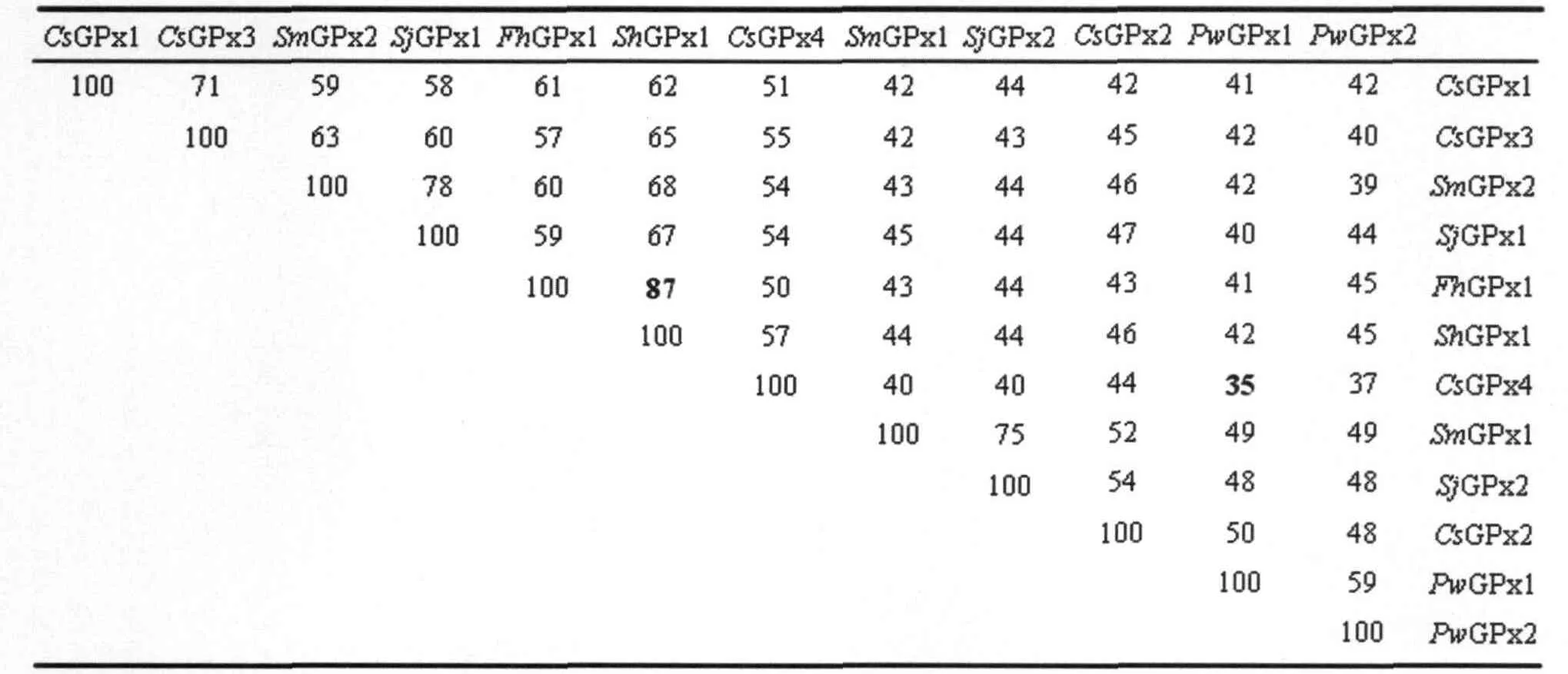
表1 寄生性吸蟲GPx基因編碼的氨基酸之間的同源性值(%)Tab.1 Identities of the GPXs from different parasitic trematodeds
吸蟲GPx與哺乳動(dòng)物宿主人(Homo sapiens,Hs)和鼠(Musmusculus,Mm)6大GPx家族成員氨基酸序列比對(duì)結(jié)果如圖1所示。從圖中可見:各家族之間具有較高的同源性,尤其在3個(gè)特征區(qū)域(即酶活性區(qū),圖中方框A、B和C)和3個(gè)保守活性殘基(圖中箭頭所示,硒半胱氨酸(U或X)/半胱氨酸(C)、谷氨酰胺(Q)、色氨酸(W)/酪氨酸(Y))具有高度的保守性,非常相似。此外,比對(duì)圖也顯示,有2個(gè)區(qū)域僅存在于GPx1(c-GPx),GPx2(GPx-GI),GPx3(p-GPx),GPx5(e-GPx)和GPx6(o-GPx)家族,而在GPx4(PHGPx)家族的成員中缺失(圖2中用虛線盒I和II表示)。吸蟲GPx氨基酸一級(jí)結(jié)構(gòu)中缺失這2個(gè)插入序列,表明吸蟲GPx均屬于PHGPx家族成員。
2.2 醫(yī)學(xué)吸蟲GPx SECIS、及與其他GPx家族SECIS分析 在分析常見醫(yī)學(xué)吸蟲(包括華支睪吸蟲、衛(wèi)氏并殖吸蟲、日本血吸蟲、曼氏血吸蟲、埃及血吸蟲和肝片形吸蟲等)12個(gè)GPx核酸序列是否存在硒半胱氨酸(Sec)和SECIS時(shí)發(fā)現(xiàn),除PwGPx1外,其他11個(gè)基因在其開放閱讀框內(nèi)均含有一非尋常“TGA”密碼子,編碼第21個(gè)稀有氨基酸Sec(見圖1,U),同時(shí)在這些基因的cDNA 3'-UTR均含有相對(duì)應(yīng)的SECIS區(qū)(如圖2B,Trematode GPx)。在其他GPx4家族成員中,Sec和相關(guān)SECIS序列僅發(fā)現(xiàn)在哺乳動(dòng)物宿主的mRNA序列中存在,而在被檢測(cè)的節(jié)肢動(dòng)物中,僅發(fā)現(xiàn)一種牛蜱(Boophilus)GPx4(ABA25916)含有Sec和SECIS。其他節(jié)肢動(dòng)物和所有植物中PHGPx均無Sec和SECIS結(jié)構(gòu)。而GPx1,GPx2,GPx3家族僅高等哺乳動(dòng)物有此特征。所有這些GPx基因的SECIS均含有幾個(gè)保守的結(jié)構(gòu)域包括:由一個(gè)內(nèi)環(huán)隔開的2個(gè)螺旋,一SECIS核心結(jié)構(gòu),一四集體位于第二個(gè)螺旋的基部和一個(gè)頂環(huán)等(圖2A)。
2.3 GPx家族系統(tǒng)發(fā)育樹分析 當(dāng)用CsGPx1的氨基酸序列進(jìn)行BALSTP同源性檢索時(shí),幾百個(gè)GPx蛋白能從GenBankTM數(shù)據(jù)庫中獲得,它們與CsGPx1有不同程度的相似性(序列同源性identities值在44%~67%之間,E值小于4e-26)。但是,由于其中絕大部分蛋白質(zhì)的氨基酸序列是PHGPx-樣蛋白,因而用人的GPx1(CAA68491)和GPx3(AAP50261)及線蟲CAA48882 GPx3-like再進(jìn)行一次BLASTP搜索,即獲得更多其他GPx的成員信息。另外配備在Sanger和 TIGR的各種醫(yī)學(xué)蠕蟲的EST數(shù)據(jù)庫也用GPx1和GPx3及GPx4進(jìn)行篩選,最后有選擇性地挑選一批代表各家族成員共107個(gè)進(jìn)行系統(tǒng)發(fā)育樹分析。
系統(tǒng)發(fā)育樹所選擇的GPx成員包括從真菌(Fungi)、節(jié)肢動(dòng)物(Arachnida)、醫(yī)學(xué)吸蟲(Trematoda)、營(yíng)自生生活線蟲(Soil nematoda)、寄生性線蟲(Parasitic nematoda)、無脊椎動(dòng)物(Invertebra-ta)、脊椎動(dòng)物門(Vertebrata)和植物(Plant)等各分類層次代表。結(jié)果顯示:根據(jù)在生物學(xué)分類上不同的地位,各類生物的GPx均一般聚集在一起。迄今Fungi的GPx屬哪個(gè)家族不是很清楚,GenBank數(shù)據(jù)庫中有的認(rèn)為是PHGPx家族,有的沒有具體分類。包括人和鼠等高等脊椎動(dòng)物的GPx被分為至少6大家族,而且發(fā)現(xiàn)每個(gè)基因在其基因組內(nèi)均以單拷貝形式存在。在較低等的動(dòng)物中,GPx蛋白存在明顯的偏向分布:除蛛形綱(Arachnida)的Ixodes scapulans(AAY66814)外,其他節(jié)肢動(dòng)物、吸蟲以及植物等的GPxs蛋白與脊椎動(dòng)物(包括哺乳動(dòng)物和人)的PHGPx分布在同一分枝,具有同源性。線蟲的GPx基因具有較復(fù)雜的分布:Parasitic nematoda的GPx蛋白顯示與脊椎動(dòng)物的GPx1/GPx3 lineage具有更相近的拓?fù)潢P(guān)系,而Soil nematoda中Caenorhabditiselegans有兩種情況,有些GPx分布在GPx3 lineage,有些分布在GPx4 lineage。此外,系統(tǒng)發(fā)育樹亦顯示:吸蟲和昆蟲的PHGPxs均單獨(dú)分成了兩個(gè)不同的亞類。如在吸蟲中,CsGPx2、SmGPx1、PwGPx1和PwGPx2被歸類在吸蟲I亞類 ,而 CsGPx1、CsGPx3、CsGPx4 與 SmGPx2 被歸類在吸蟲II亞類。

圖1 寄生性吸蟲GPx與哺乳動(dòng)物宿主(人和鼠)6大GPx家族成員的氨基酸多重比對(duì)分析Fig.1 Multiple sequence alignment of parastic trematodes GPxs with members of six GPx families from Mammalian host(Homo sapiens and Musmusculus)
2.4 醫(yī)學(xué)吸蟲GPx基因結(jié)構(gòu)及與其他GPx基因結(jié)構(gòu)分析 通過網(wǎng)絡(luò)數(shù)據(jù)庫和信息學(xué)分析,共獲得8個(gè)吸蟲GPx基因的完整基因組結(jié)構(gòu),見圖3。除CsGPx4是由5個(gè)外顯子和4個(gè)內(nèi)含子組成外,其他幾種吸蟲GPx基因均被5個(gè)內(nèi)含子剪接為6個(gè)外顯子,且所有這些吸蟲GPx基因結(jié)構(gòu)高度保守。
各吸蟲GPx與不同等級(jí)生物(包括高等脊椎動(dòng)物、線蟲、昆蟲和植物)PHGPx的基因組結(jié)構(gòu)比較結(jié)果如圖3所示:根據(jù)它們的內(nèi)含子的數(shù)量和位置,各不同等級(jí)生物代表物種GPx4基因分享相對(duì)比較保守的基因組結(jié)構(gòu);外顯子—內(nèi)含子的結(jié)構(gòu)在吸蟲和哺乳動(dòng)物宿主之間存在高度相似性。在吸蟲GPx基因的 5個(gè)內(nèi)含子中,有 4個(gè)跟哺乳動(dòng)物GPx4基因在位置和ORF剪接插入位置均相同(圖中橫線上數(shù)字所示);從營(yíng)自由生活的線蟲C.elegans和C.briggsae分離的GPx4基因的第1個(gè)內(nèi)含子跟吸蟲和哺乳動(dòng)物的第2個(gè)內(nèi)含子類似,具有一定程度的同源性;從昆蟲分離的PHGPx基因與吸蟲和脊椎動(dòng)物的基因沒有顯示任何結(jié)構(gòu)上的關(guān)系,反而與從植物分離的GPx基因存在某種程度的同源性。
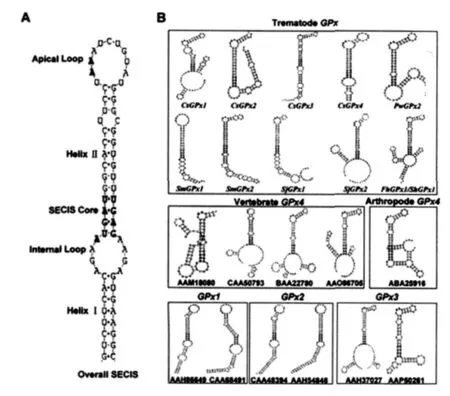
圖2 寄生性吸蟲GPx與其他GPx家族相關(guān)成員mRNA中3'-UTR的SECIS分析Fig.2 Analysisof SECIS motif fromdifferent parastic trematodes and other related members of mRNA3'-UTR regions
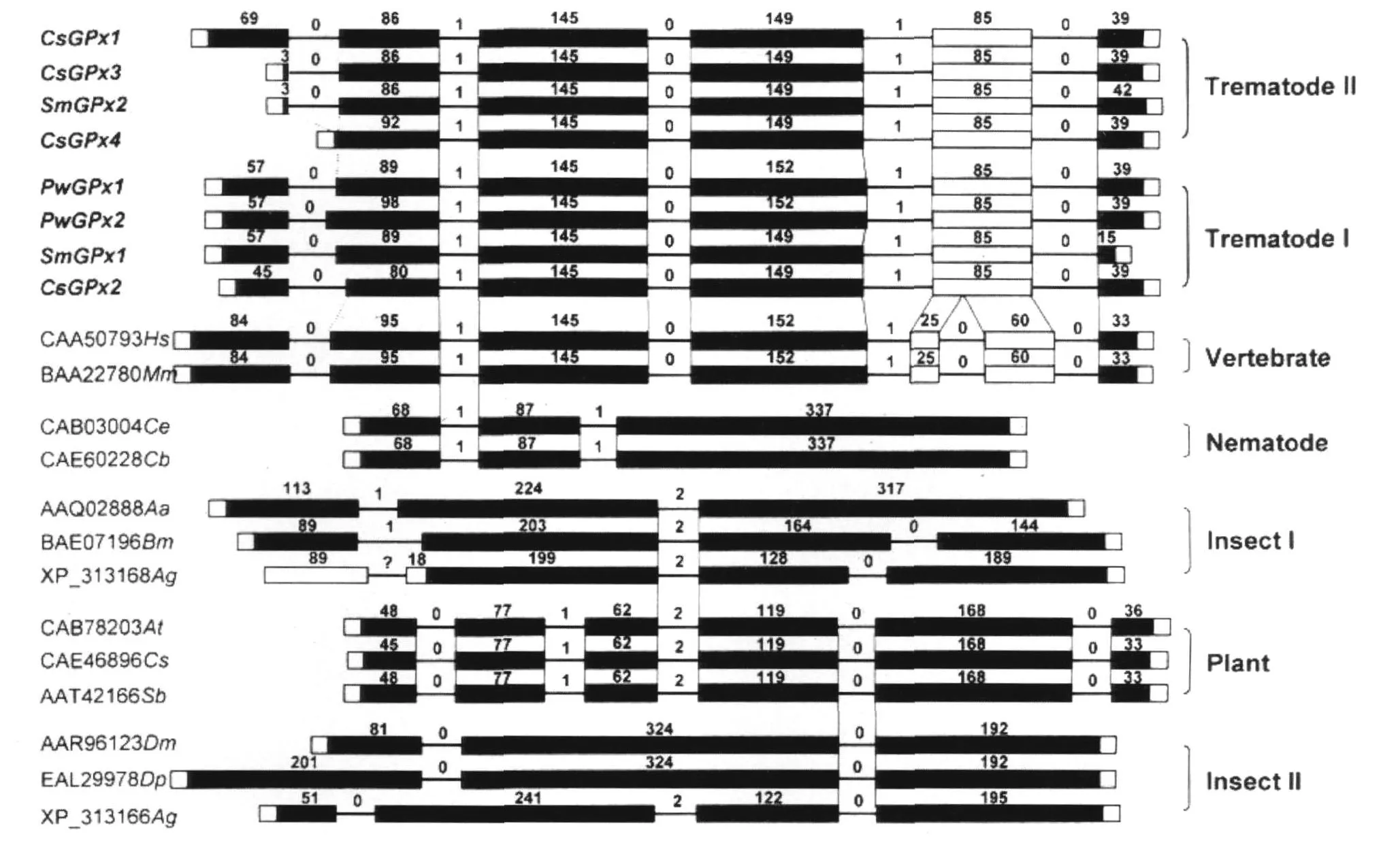
圖3 寄生性吸蟲 GPx和其他相關(guān)PHGPx“外顯子-內(nèi)含子”基因組結(jié)構(gòu)比較分析Fig.3 Exon-intron structures analysis between the parastic trematode GPx genes and other related PHGPx genes from various organisms
3 討 論
在所有生物體中(包括細(xì)菌、植物和動(dòng)物),GPx被認(rèn)為是在抗-ROS防御機(jī)制中重要的關(guān)鍵酶〔2〕。PHGPx由于其不僅能降解H2O2,而且還能直接降解整合在細(xì)胞膜上的磷脂化合物,因而被認(rèn)為是一種獨(dú)特的抗氧化酶〔5〕。本研究通過網(wǎng)絡(luò)資源,搜尋到多種重要醫(yī)學(xué)吸蟲GPx序列。通過氨基酸比對(duì)、系統(tǒng)發(fā)育樹和Sec/SECIS的分析,證明醫(yī)學(xué)吸蟲GPx分子均為PHGPx家族成員。一般認(rèn)為PHGPx在其3個(gè)催化區(qū)域中,有一個(gè)位點(diǎn)含有Sec。Sec插入蛋白質(zhì)的合成過程受其mRNA的框內(nèi)UGA密碼子和它的下游鏈的莖-環(huán)(stem-loop)結(jié)構(gòu)所控制,這種特殊的莖-環(huán)結(jié)構(gòu)稱為硒半胱氨酸插入元件(SECIS)。在生物學(xué)上,把含有Sec的蛋白稱為硒蛋 白 (selenoprotein)〔6〕。分 析 發(fā) 現(xiàn),吸 蟲 除PwGPx1基因外,其它均為含硒蛋白(如圖1和圖2)。不同生物體內(nèi),硒-依賴PHGPx(sPHGPx)和硒-非依賴PHGPx(siPHGPx)存在嚴(yán)重的偏向分布特征。圖2顯示,除一節(jié)肢動(dòng)物Boophilus microplus外,僅在脊椎動(dòng)物和吸蟲的PHGPx mRNA中檢測(cè)到含有Sec和SECIS序列。以往研究報(bào)道認(rèn)為,sPHGPx比siPHGPx酶的活性要更高,其原因是由于在生理?xiàng)l件下sPHGPx(pKa=5.2)比siPHGPx(pKa≥8.0)更容易使還原狀態(tài)轉(zhuǎn)變?yōu)檠趸癄顟B(tài)〔7〕。因此,siPHGPx酶可能存在另外的機(jī)制以一種新的方式發(fā)揮其在細(xì)胞內(nèi)抗氧化的功能〔8〕。或者,也有可能siPHGPx作為抗氧化酶在特殊的細(xì)胞器中發(fā)揮作用,這些細(xì)胞器內(nèi)可能受半胱氨酸的巰基基團(tuán)離子能的作用,使得其有特殊的pH微環(huán)境。
在分析基因組結(jié)構(gòu)時(shí),內(nèi)含子的獲得或丟失、以及內(nèi)含子的數(shù)量和長(zhǎng)度大小均可反應(yīng)生物體之間的進(jìn)化關(guān)系〔9〕。在吸蟲GPx基因中,內(nèi)含子1、4和 5存在明顯的長(zhǎng)度多態(tài)性現(xiàn)象。CsGPx1、CsGPx3、CsGPx4和SmGPx2的第5內(nèi)含子(長(zhǎng)度分別為6,768、2,960、4,851和 1,438bp)明顯地大于哺乳動(dòng)物基因(如人CAA50793為 80bp,鼠 BAA22780為86bp);而 PwGPx1、PwGPx2、CsGPx2 和 SmGPx1的第1內(nèi)含子(長(zhǎng)度分別為 1,772、2,497、2,082和1,290bp)和第4內(nèi)含子(長(zhǎng)度分別為3,018、5,889、2,054和1,609bp)均明顯地大于哺乳動(dòng)物基因(如人CAA50793為1,058和432bp,鼠BAA22780為798和803bp),因而吸蟲GPx基因分成了2亞類。總的來說,內(nèi)含子的長(zhǎng)度和數(shù)量還是展現(xiàn)出一定的規(guī)律,即與生物體基因組的復(fù)雜性相關(guān)〔10〕,并且這些延長(zhǎng)的內(nèi)含子是與轉(zhuǎn)座子插入有關(guān)〔11〕。在寄生性吸蟲基因組中,如存在最大長(zhǎng)度多態(tài)性現(xiàn)象的CsGPx1和CsGPx2基因的第5內(nèi)含子就整合了一非長(zhǎng)末端重復(fù)序列反轉(zhuǎn)座子(non-LTR retrotransposons),其與曼氏血吸蟲的SR2非長(zhǎng)末端重復(fù)序列反轉(zhuǎn)座子同源。PwGPx1和PwGPx2基因的第4個(gè)內(nèi)含子也分別整合了一與曼氏血吸蟲的penelope-like LT R retrotransposon,Perere-10同源的反轉(zhuǎn)座子和一與曼氏血吸蟲non-LTR retrotransposon,Perere-6同源的非長(zhǎng)末端重復(fù)序列反轉(zhuǎn)座子。
比較基因組學(xué)分析表明,吸蟲PHGPx基因結(jié)構(gòu)與哺乳動(dòng)物PHGPx具有更高的同源性(圖3),表明寄生蟲與宿主之間PHGPx基因可能是從共同的祖先基因逐漸進(jìn)化而來,此基因含有6個(gè)外顯子和5個(gè)內(nèi)含子。在進(jìn)化過程中,哺乳動(dòng)物PHGPx基因獲得一內(nèi)含子,而吸蟲 PHGPx基因通過復(fù)制(duplication),然后形成分支(divergence)。第一分支(Trematode I)保留了祖先基因結(jié)構(gòu),而第5個(gè)內(nèi)含子的長(zhǎng)度得到延伸(如CsGPx2,PwGPx1,PwGPx2和SmGPx1)。第二分支(T rematode II)內(nèi)含子1和內(nèi)含子4的長(zhǎng)度則變短(如CsGPx1,CsGPx3,CsGPx4和SmGPx2)。另外,在生物進(jìn)化過程中,一些PHGPx基因在祖先基因的第1和第2外顯子之間丟失了1個(gè)內(nèi)含子(如CsGPx4),而有些PHGPx基因保持了祖先基因的結(jié)構(gòu)或者丟失了信號(hào)肽序列(如CsGPx3)。更復(fù)雜的分析包括昆蟲、線蟲和植物等在內(nèi)的所有不同分類等級(jí)生物體的PHGPx基因的進(jìn)化方式需更進(jìn)一步的分析。
〔1〕A rthur,JR.The glutathione peroxidases〔J〕.Cell Mol Life Sci,2000,57(13-14):1825-1835.
〔2〕Imai H,Nakagawa Y.Biological significance of phospholipid hydroperoxide glutathione peroxidase(PHGPx,GPx4)in mammalian cells〔J〕.Free Radic Biol Med,2003,34(2):145-169.
〔3〕Cookson E,Blaxter M L,Selkirk ME.Identification of the major soluble cuticular glycoprotein of lymphatic filarial nematode parasites(gp29)as a secretory homolog of glutathione peroxidase〔J〕.Proc Natl Acad Sci USA,1992,89(13):5837-5841.
〔4〕Zelck UE,Von Janowsky B.Antioxidant enzymes in intramolluscan Schistosoma mansoni and ROS-induced changes in ex pression 〔J〕.Parasitology,2004,128(Pt 5):493-501.
〔5〕U rsini F,Bindoli A.T he role of selenium peroxidases in the protection against oxidative damage of membranes〔J〕.Chem Phys Lipids,1987,44(2-4):255-276.
〔6〕Kryukov GV,Castellano S,Novoselov SV,et al.Characterization of mammalian selenoproteomes〔J〕.Science,2003,300(5624):1439-1443.
〔7〕Stadtman TC.Selenocy steine 〔J〕.Annu Rev Biochem,1996,65:83-100.
〔8〕Utomo A,Jiang X,Furuta S,et al.Identification of a novel putative non-selenocy steine containing phospholipid hydroperoxide glutathione peroxidase(NPGPx)essential for alleviating oxidative stress generated from polyunsaturated fatty acids in breast cancer cells〔J〕.J Biol Chem,2004,279(42):43522-43529.
〔9〕Jeffares DC,M ourier T,and Penny D.The biology of intron gain and loss〔J〕.Trends Genet,2006,22(1):16-22.
〔10〕Lynch M,Conery JS.The origins of genome complexity 〔J〕.Science,2003,302(5649):1401-1404.
〔11〕Drew AC,Minchella DJ,King LT,et al.SR2 elements,nonlong terminal repeat rctrotransposons of the RTE-1 lineage from the human flood fluke schistosoma mansoni〔J〕.M ol Biol Evol,1999,16(9):1256-1269.
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
