瀕危植物白桂木的遺傳多樣性研究
2010-07-24 01:30:34范繁榮
浙江農(nóng)林大學(xué)學(xué)報(bào) 2010年2期
范繁榮
(福建三明林業(yè)學(xué)校, 福建 三明365001)
白桂木Artocarpus hypargyreus 是桑科Moraceae 波羅蜜屬Artocarpus 常綠喬木, 是一種經(jīng)濟(jì)價(jià)值很高的樹種。 種子含有豐富的白桂木凝集素, 正引起醫(yī)藥學(xué)界的廣泛注意。 白桂木樹干高大, 樹形優(yōu)美, 枝葉繁茂, 遮蔭面寬, 也是園林綠化的優(yōu)良樹種。 現(xiàn)主要零星分布于福建南部和西部、 江西南部、 湖南東南部和西南部、 廣東北部、 海南、 廣西、 云南東南部。 長期以來, 由于人們對(duì)白桂木天然林的破壞性經(jīng)營, 加上白桂木自身生物學(xué)特性及自然環(huán)境條件變化等方面的原因, 致使白桂木野生資源接近枯竭, 處于瀕危境地, 已被列為國家三級(jí)瀕危保護(hù)樹種[1]。 目前, 關(guān)于白桂木的研究很少, 僅局限于白桂木藥用化學(xué)成分的研究[2-6]和白桂木形態(tài)特征、 生態(tài)特性及地理分布方面的報(bào)道[7], 對(duì)白桂木瀕危的內(nèi)在機(jī)制及保育技術(shù)研究還是空白。 遺傳多樣性是生物種內(nèi)表現(xiàn)在從DNA 分子到形態(tài)特征各個(gè)層次上可遺傳所有變異的總稱, 是瀕危植物保護(hù)生物學(xué)研究的核心之一[8]。 對(duì)瀕危物種遺傳多樣性和群體遺傳結(jié)構(gòu)的研究是揭示其適應(yīng)潛力的基礎(chǔ), 也為進(jìn)一步探討瀕危物種的瀕危機(jī)制和制定相應(yīng)的保護(hù)措施提供科學(xué)依據(jù)[9]。 隨機(jī)擴(kuò)增多態(tài)性DNA(RAPD, random amplified polymorphic DNA)分析不受器官和組織種類、 發(fā)育階段和生境條件等因素的影響, 需要的DNA 模板量少, 質(zhì)量要求不高,多態(tài)性豐富, 具有簡(jiǎn)便、 快速、 易行、 價(jià)廉等優(yōu)點(diǎn), 為從分子水平研究生物遺傳多樣性中應(yīng)用最廣和最具代表性的方法之一[10]。 RAPD 分子標(biāo)記用于屬以下等級(jí)特別是種內(nèi)各居群或品種的研究具有很強(qiáng)的優(yōu)越性[11]。 本研究利用RAPD 分子標(biāo)記技術(shù)對(duì)福建不同白桂木天然種群的遺傳多樣性進(jìn)行分析, 為從遺傳多樣性角度揭示白桂木的瀕危機(jī)制和更好地保護(hù)白桂木種群提供科學(xué)依據(jù)。
1 材料與方法
1.1 材料采集
實(shí)驗(yàn)材料分別采自福建白桂木保存較好的三明市梅列區(qū)陳大、 三元區(qū)莘口, 泉州市永春縣牛姆林, 清流縣沙蕪, 永安市貢川的白桂木天然林。 由于各個(gè)種群的白桂木個(gè)體數(shù)量較少, 因此, 只能在各個(gè)采樣區(qū)隨機(jī)選取生長良好的3 株白桂木(株間間隔必須5 m 以上), 采集白桂木健康小枝, 并將它們迅速放入密封塑料袋中, 裝入冰壺帶回實(shí)驗(yàn)室, 放在- 80 ℃超低溫冰箱保存?zhèn)溆谩?/p>
1.2 DNA 提取
采用改進(jìn)的十六烷基三甲基溴化銨(CTAB)法, 參照王關(guān)林等[12]的方法提取DNA。 稱取葉片0.3 g左右, 加液氮快速充分研磨至粉末狀, 加入0.1 g 聚乙烯吡咯烷酮(PVP), 將粉末移至1.5 mL 離心管中; 加入600 μL·管-1預(yù)熱至65 ℃的2 × CTAB 提取緩沖液, 同時(shí)加入7 μL 2-琉基乙醇, 充分混勻,65 ℃水浴30 min, 其間顛倒幾次; 取出離心管, 冷至室溫, 加等體積的氯仿/異戊醇(24 ∶1), 輕緩顛倒混勻, 靜置10 min, 室溫下1.2 萬r·min-1離心10 min; 取上清液至另一離心管, 加2/3 體積異丙醇, 輕緩顛倒混勻, 室溫放置15 min, 于4 ℃條件下1.2 萬r·min-1離心10 min; 用體積分?jǐn)?shù)為70%乙醇洗滌DNA 2~3 次, 風(fēng)干, 將DNA 溶于500 μL TE(tris-EDTA, 三羥甲基氨基甲烷-乙二胺四乙酸)溶液, 加入質(zhì)量濃度為50 g·L-1的RNase A, 并于37 ℃保溫1 h; 用等體積氯仿/異戊醇(24 ∶1)抽提1 ~3 次; 上清液中加入終濃度為0.2 ~ 0.4 mol·L-1的氯化鈉, 2 倍體積的無水乙醇, 放置1 h 左右, 1.2萬r·min-1離心10 min; 除去上清液; 用體積分?jǐn)?shù)為70%乙醇洗滌沉淀2 ~ 3 次, 風(fēng)干后溶于50 μL TE 中備用。
1.3 聚合酶鏈?zhǔn)椒磻?yīng)(PCR)擴(kuò)增與產(chǎn)物的檢測(cè)
擴(kuò)增體系采用25.0 μL 體系: 25 ng 模板DNA, 0.40 μmol·L-1引物, 0.15 mmol·L-1三磷酸堿基脫氧核苷酸(dNTP), 16.67 nkat TaqDNA 聚合酶, 1.50 mmol·L-1氯化鎂, 2.5 μL 10 ×聚合酶鏈?zhǔn)椒磻?yīng)(PCR)緩沖液, 加入滅菌的超純水至25.0 μL。
PCR 擴(kuò)增條件為: 94 ℃預(yù)變性5 min, 94 ℃變性1 min, 36 ℃1 min, 72 ℃延伸2 min, 共40 個(gè)循環(huán), 72 ℃延伸10 min。 擴(kuò)增完畢, 取經(jīng)15.0 g·L-1瓊脂糖凝膠電泳檢查。
1.4 數(shù)據(jù)統(tǒng)計(jì)與分析
RAPD 是顯性標(biāo)記, 同一引物擴(kuò)增產(chǎn)物中電泳遷移率一致的條帶被認(rèn)為具有同源性。 根據(jù)反應(yīng)產(chǎn)物在凝膠對(duì)應(yīng)位置上條帶的有無, 有帶記為1, 無帶記為0, 得到RAPD 分析的原始數(shù)據(jù)。 使用POPGEN 32 軟件計(jì)算多態(tài)位點(diǎn)百分率、 Shannon 指數(shù)、 Nei 指數(shù)和基因流(Nm); 運(yùn)用Nei 指數(shù)法[13]計(jì)算遺傳距離 (D)和遺傳一致度 (F), 聚類分析運(yùn)用DPS 軟件采用非加權(quán)配對(duì)算術(shù)平均法 (UPGMA, unweighted pai-group method using an arithmetic average)分析。
2 結(jié)果與分析
2.1 引物篩選
隨機(jī)引物購自上海生工生物工程有限公司。 用5 個(gè)DNA 樣品(每個(gè)種群選1 個(gè)樣品)對(duì)隨機(jī)引物進(jìn)行篩選, 從60 個(gè)引物中篩選出S237, S248,S249, S262, S263, S271 等6 個(gè)擴(kuò)增條帶清晰、 重復(fù)性好的引物(表1), 對(duì)所有個(gè)體進(jìn)行擴(kuò)增。

表1 RAPD 分析的6 個(gè)隨機(jī)引物序列Table 1 Sequences of 6 random primers used in RAPD analysis
2.2 白桂木種群遺傳多樣性分析
2.2.1 多態(tài)位點(diǎn)百分率 利用6 個(gè)隨機(jī)引物對(duì)5 個(gè)白桂木種群15 個(gè)個(gè)體DNA 樣品進(jìn)行RAPD 分析,共擴(kuò)增出37 個(gè)條帶, 其中多態(tài)條帶30 個(gè), 總的多態(tài)位點(diǎn)百分率為81.08%, 平均可獲得多態(tài)位點(diǎn)5個(gè)·引物-1。 引物S263, S269 和S271 對(duì)白桂木不同個(gè)體擴(kuò)增的帶型見圖1 ~ 3。 各居群間多態(tài)位點(diǎn)百分率存在著較大差異, 其中永安種群最高, 為24.32%, 梅列種群最低, 為8.11%, 平均多態(tài)位點(diǎn)百分率為15.67%, 其大小排序?yàn)橛腊卜N群>三元種群>永春種群>清流種群>梅列種群。

圖1 引物S263 擴(kuò)增的RAPD 帶型Figure 1 RAPD amplification products from primer S263

圖2 引物S269 擴(kuò)增的RAPD 帶型Figure 2 RAPD amplification products from primer S269
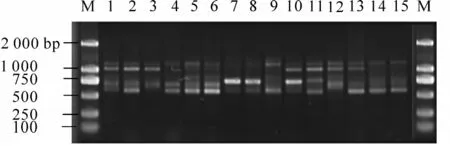
圖3 引物S271 擴(kuò)增的RAPD 帶型Figure 3 RAPD amplification products from primer S271
2.2.2 Shannon 多樣性和Nei 基因多樣性指數(shù) Shannon 指數(shù)是生態(tài)學(xué)中用于度量物種多樣性最常用的指標(biāo)[14]。 從表2 的估算結(jié)果可看出, 白桂木總遺傳多樣性指數(shù)為0.231 9, 各種群平均多樣性指數(shù)為0.219 3, 其中最高的是永安種群, 為0.261 7, 其大小排序?yàn)榍辶鞣N群>三元種群>梅列種群>永春種群, 最小的永春種群僅為0.181 9。 從表2 中可見, 由Nei 指數(shù)估算的白桂木總遺傳多樣性為0.166 0, 各種群平均

表2 不同白桂木種群的遺傳多樣性Table 2 Genetic diversity of Artocarpus hypargyreus
遺傳多樣性為0.152 6, 其中最高的是永安種群, 為0.182 0, 其他大小排序?yàn)榍辶鞣N群>三元種群>梅列種群>永春種群, 最小的永春種群僅為0.120 2。 由該指數(shù)估算的遺傳多樣性較Shannon 指數(shù)估算的低, 但其大小排序與Shannon 多樣性指數(shù)基本一致。 與瀕危植物銀杉Cathaya argyrophylla 和珙桐Davidia involucrata 相比, 白桂木的遺傳多樣性高于銀杉[11], 低于珙桐[15]。
2.3 白桂木種群間的遺傳分化
由Nei 指數(shù)計(jì)算的白桂木種群間的遺傳分化系數(shù)結(jié)果見表3。 白桂木種群間的分化系數(shù)GST為0.737 8, 白桂木的遺傳變異有73.78%存在于種群之間, 只有26.22%的遺傳變異存在于種群內(nèi), 這與蘇何玲等[16]對(duì)瀕危植物冷杉Abies ziyuanensis 的研究結(jié)果相似。 可見白桂木種群間的分化程度較高, 種群內(nèi)的分化程度較低。

表3 Nei 指數(shù)估算的白桂木種群間遺傳分化Table 3 Genetic differentiations among 5 populations estimated by Nei’ index
基因流Nm的計(jì)算結(jié)果為0.125 1, 表明白桂木種群間的基因流動(dòng)較小, 這與由Nei 指數(shù)計(jì)算的種群間的遺傳分化系數(shù)所得的大部分遺傳變異存在于種群間的結(jié)論相一致。 Wright 認(rèn)為種群間基因流大于1, 則能發(fā)揮其均質(zhì)化作用; 反之若小于1, 則說明基因流成為遺傳分化的主要原因[17]。 白桂木種群在自然狀態(tài)下分布零散, 各種群呈“島嶼” 狀, 種群間的空間距離遠(yuǎn), 花粉的傳播受限, 同時(shí)其生殖生態(tài)學(xué)研究發(fā)現(xiàn)白桂木個(gè)體數(shù)量少、 雄雌花序的分布不利于授粉、 開花時(shí)期雨水多等原因造成結(jié)實(shí)量低, 加上動(dòng)物的取食, 自然狀態(tài)下可供更新的有效種子數(shù)量少。 同時(shí)種子含水量高, 不易儲(chǔ)存, 種子有后熟現(xiàn)象, 在自然條件下發(fā)芽困難, 種子成苗率低, 自然更新能力差。 有效種子流的形成受到極大的限制, 使得白桂木種群間的基因流極低, 只有0.125 1, 遠(yuǎn)小于1.000, 極低的基因流造成種群間很高的遺傳分化。
2.4 白桂木種群的遺傳距離與遺傳相似度
遺傳距離和遺傳相似度是衡量植物變異水平的重要指標(biāo)。 白桂木種群間的遺傳距離和遺傳相似度計(jì)算結(jié)果見表4。 從表4 中可看出, 梅列種群與三元種群的相似度最大, 為0.747 7, 它們之間的地理距離最近, 永春種群與清流種群的的相似度最小, 為0.528 7, 它們之間的地理距離也是最大的。 遺傳距離則是永春種群與清流種群最大,為0.637 3, 梅列種群與三元種群的遺傳距離最小, 為0.290 7, 5 個(gè)不同種群的平均相似度和遺傳距離分別為0.6665 和0.411 8。

表4 不同白桂木種群的遺傳相似度與遺傳距離Table 4 Genetic distance and genetic identity of populations of Artocarpus hypargyreus
2.5 聚類分析
種群間的遺傳距離一般隨著地理距離的增大而增大[18]。 根據(jù)RAPD 基因表型矩陣, 通過UPGMA法進(jìn)行不同白桂木種群的聚類分析, 得到白桂木個(gè)體間的樹狀圖(圖4), 進(jìn)一步反映了白桂木種群間的親緣關(guān)系。 從聚類結(jié)果可看出, 聚類結(jié)果與該種群所處的地理緯度位置比較一致, 地理位置相對(duì)最近的梅列種群和三元種群先聚在一起, 然后是清流種群和永安種群, 最后是地理位置最遠(yuǎn)的永春種群。
3 討論
通常認(rèn)為, 瀕危物種因種群縮小而導(dǎo)致近親繁殖和等位基因的隨機(jī)固定, 從而降低了其種群生存能力, 遺傳多樣性水平較低, 如華木蓮Sinomanglietia glauca 和銀杉等[9,19], 而黃久香等[20]和惲銳等[21]對(duì)觀光木Tsoongiodendron odorum 和遼東櫟Quercus liaotungensis 的研究發(fā)現(xiàn)有些瀕危物種也保持持著較高的遺傳多樣性水平。 白桂木遺傳多樣性水平的分析結(jié)果表明: 白桂木種群具有較高的遺傳多樣性, 總的多態(tài)位點(diǎn)頻率為81.08%, 與觀光木(84.62%)和遼東櫟(79.00%)相當(dāng), 遠(yuǎn)高于銀杉(32.00%)和華木蓮(53.22%)。 平均Shannon指數(shù)為0.231 9, Nei 指數(shù)為0.166 0, 白桂木的遺傳分化系數(shù) (GST)為0.737 8, 基因流 (Nm)為0.125 1。 說明白桂木種群間的基因流受限嚴(yán)重,其遺傳變異有73.78%存在于種群間, 只有26.22%發(fā)生于種群內(nèi), 可見白桂木種群間的分化程度較高, 種群內(nèi)的分化程度較低, 這與瀕危植物冷杉相似。

圖4 不同白桂木種群的UPGMA 聚類圖Figure 4 UPGMA dendrogram for populations of Artocarpus hypargyreus
通過UPGMA 聚類分析表明: 不同白桂木種群的聚類結(jié)果與其地理緯度位置基本一致。 地理位置最近的先聚一起, 其次相對(duì)較近的聚在一起, 最后是距離最遠(yuǎn)的。 可見不同白桂木種群間的基因交流少, 小種群分化嚴(yán)重, 種群內(nèi)雜合度低,適應(yīng)性弱, 是其種群衰退的重要原因。 同時(shí), 白桂木的物種具有較高的遺傳多樣性, 因此在就地保護(hù)基礎(chǔ)上, 更要注意遷地保護(hù)。 由于其種群間基因流很小, 遺傳分化強(qiáng)烈, 應(yīng)注意多種群采種, 進(jìn)行人工繁殖, 尤其應(yīng)在遺傳多樣性比較高的種群中采種, 采用扦插和組織培養(yǎng)等快繁途徑來擴(kuò)大和更新現(xiàn)有種群。 同時(shí)通過打破白桂木種子休眠, 創(chuàng)造條件提高白桂木種子發(fā)芽率, 在較大的空間范圍進(jìn)行播種, 增加該種群的幼苗數(shù)量, 創(chuàng)造基因交流和重組的條件, 進(jìn)一步保護(hù)白桂木的遺傳多樣性。
[1] 傅立國. 中國植物紅皮書[M]. 北京: 北京科學(xué)技術(shù)出版社, 1992: 98.
[2] 甘茂羅, 歐陽勝, 謝平, 等. 白桂木的化學(xué)成分研究[J]. 中草藥, 2005, 36 (7): 988-989.GAN Maoluo, OUYANG Sheng, XIE Ping, et al. Study on chemical composition of Artocarpus hypargyreus [J].Chin Tradit Herbal Drugs, 2005, 36 (7): 988-989.
[3] 陳黎明, 謝平, 肖慶青, 等. 白桂木化學(xué)成分研究[J]. 中草藥, 2007, 38 (6): 815-818.CHEN liming, XIE Ping, XIAO Qingqing, et al.Study on chemical composition of Artocarpus hypargyreus [J]. Chin Tradit Herbal Drugs, 2007, 38 (6): 815-818.
[4] 周德義, 楊爾濱, 鄧勇, 等. 白桂木凝集素的純化與性質(zhì)的研究[J]. 生物化學(xué)與生物物理學(xué)報(bào), 1995, 17 (1): 61- 66.ZHOU Deyi, YANG Erbin, DENG Yong, et al. Purification and characterization of lectin from the seeds of Artocarpus hypargyreus Hance [J]. Acta Biochem Biophys Sin, 1995, 17 (1): 61-66.
[5] 吳耀生, 張紅, 周素芳, 等. 酶標(biāo)白桂木凝集素糖蛋白結(jié)合特性的分析[J]. 中國生物化學(xué)與分子生物學(xué)報(bào), 2000,16 (2): 210-214.WU Yaosheng, ZHANG Hong, ZHOU Sufang, et al. Characterization of Artocarpus hypargyreus Hance lectin interacting with glycoproteins [J]. Chin J Biochem Mol Biol, 2000, 16 (2): 210-214.
[6] 鄧勇, 周德義, 關(guān)祺芳, 等.紅桂木、 白桂木、 木菠蘿種子凝集素生物學(xué)性質(zhì)的比較分析[J].廣西醫(yī)科大學(xué)學(xué)報(bào), 1995,12 (2): 183-185.DENG Yong, ZHOU Deyi, GUAN Qifang, et al. Biology characteristic analysis of seed agglutinin between Artocarpus lignanensis Merr, Artocarpus hypargyreus Hance and Artocarpus heterophyllus Lam. [J]. J Guangxi Med Univ, 1995, 12(2): 183-185.
[7] 陳開團(tuán), 張宗華, 蔣延生.福建省25 種珍貴樹種的分布現(xiàn)狀與保護(hù)對(duì)策的初步研究[J]. 福建林業(yè)科技, 2003, 30 (2):70-73.CHEN Kaituan, ZHANG Zhonghua, JIANG Yansheng. Preliminary on the distribution status and protective countermeasures of rare tree species in Fujian Province [J]. J Fujian For Sci Technol, 2003, 30 (2): 70-73.
[8] 祖元?jiǎng)偅?張文輝, 閻秀峰, 等. 瀕危植物裂葉沙參保護(hù)生物學(xué)[M]. 北京: 科學(xué)出版社, 1999: 182-223.
[9] 葛頌, 王海群, 張大明. 八面山銀杉林的遺傳多樣性和群體分化[J]. 植物學(xué)報(bào), 1997, 39 (3): 266-271.GE Song, WANG Haiqun, ZHANG Daming. Genetic diversity and population differentiation of Cathaya argyrophylla Chun et Kuang in Bamian Mountain [J]. Chin Bull Bot, 1997, 39 (3): 266-271.
[10] 蔣志剛, 馬克平, 韓興國. 保護(hù)生物學(xué)[M]. 杭州: 浙江科學(xué)技術(shù)出版社, 1997.
[11] 汪小全, 鄒喻蘋, 張大明, 等. RAPD 應(yīng)用于遺傳多樣性和系統(tǒng)學(xué)研究中的問題[J]. 植物學(xué)報(bào), 1996, 38 (12):954-962 WANG Xiaoquan, ZOU Yuping, ZHANG Daming, et al. Problems in the use of RAPD to the study of genetic diversity and systematics [J]. Chin Bull Bot, 1996, 38 (12): 954-962.
[12] 王關(guān)林, 方宏筠. 植物基因工程[M]. 北京: 科學(xué)出版社, 2002: 744.
[13] NEI M, LI W H. Mathematical model for studying genetic variation in terms of restriction endonucleases [J]. Proc Natl Acad Sci USA, 1979, 76: 5269-5273.
[14] 羅美娟, 葉功富, 盧昌義. 短枝木麻黃群體的遺傳分化和遺傳結(jié)構(gòu)[J]. 福建林學(xué)院學(xué)報(bào), 2007, 27 (4): 343-348.LUO Meijuan, YE Gongfu, LU Cangyi. Genetic differentiation and structure of Casuarina equisetifolia populations [J]. J Fujian Coll For, 2007, 27 (4): 343-348.
[15] 宋叢文, 包滿珠. 天然珙桐群體的RAPD 標(biāo)記遺傳多樣性研究[J]. 林業(yè)科學(xué), 2004, 40 (4): 75-79.SONG Congwen, BAO Manzhu. Study on genetic diversity of RAPD mark for natural Davidia involucrata population [J].Sci Silv Sin, 2004, 40 (4): 75-79.
[16] 蘇何玲, 唐紹清. 瀕危植物資源冷杉遺傳多樣性研究[J]. 廣西植物, 2004, 24 (5): 414-417.SU Heling, TANG Shaoqing. Genetic diversity of the endangered plant Abies ziyuanensis in two populations [J]. Guihaia, 2004, 24 (5): 414-417.
[17] WRIGHT S. Evolution in Mendelian population [J]. Genetics, 1931, 16: 97-159.
[18] NEBAUER S G, CASTILLO-AGUDO L, SEGURA J, et al. RAPD variation within and among natural populations of outcrossing willow-leaved foxglove (Digitalis obscurea L.)[J]. Thror Appl Genet, 1999, 98: 985-994.
[19] 林新春, 俞志雄, 裘利洪, 等. 瀕危植物華木蓮的遺傳多樣性研究[J]. 江西農(nóng)業(yè)大學(xué)學(xué)報(bào): 自然科學(xué)版, 2003, 25(6): 805-810.LIN Xinchun, YU Zhixiong, QIU Lihong, et al. Studies on genetic diversity of endangered Sinomanglietia glauca(Magnoliaceae)[J]. Acta Agric Univ Jiangxi, 2003, 25 (6): 805-810.
[20] 黃久香, 莊雪影. 華南三地觀光木遺傳多樣性的RAPD 分析[J]. 華南農(nóng)業(yè)大學(xué)學(xué)報(bào), 2002, 23 (2): 54-57.HUANG Jiuxiang, ZHUANG Xueying. Comparison of genetic diversity of Tsoongiodendron odorum in southern China by RAPD markers [J]. J South China Agric Univ, 2002, 23 (2): 54-57.
[21] 惲銳, 鐘敏, 王洪新, 等. 北京東靈山遼東櫟種群DNA 多樣性的研究[J]. 植物學(xué)報(bào), 1998, 40 (2): 169-175.YUN Rui, ZHONG Ming, WANG Hongxin, et al. Study on DNA diversity of Quercus liaotungensis Koidz population[J]. Chin Bull Bot, 1998, 40 (2): 169-175.
