沉水植物苦草對上覆水各形態磷濃度的影響
2010-08-03 08:32:00陳秋敏王國祥葛緒廣王立志
水資源保護 2010年4期
陳秋敏,王國祥,葛緒廣,王立志
(1.南京師范大學地理科學學院,江蘇南京 210046;2.江蘇省環境演變與生態建設重點實驗室,江蘇南京 210046)
磷是影響湖泊富營養化的關鍵性限制因素之一[1-2],同時也是植物生長發育的必需元素。水體中各種磷形態之間的轉化,尤其是其他形態磷化合物向正磷酸鹽轉化速率是決定湖泊營養狀態和上覆水生產力的重要因素[3-4]。
沉水植物是湖泊生態系統的一個重要組成部分和主要的初級生產者之一,對湖泊生態系統中物質和能量的循環起著重要作用,沉水植物占據了湖泊中水和沉積物的主要界面,是水體兩大營養庫間的有機結合,對湖泊生產力及湖泊生態系統過程具有重要的影響[5],沉水植物在生長發育、衰亡及季相交替過程中,通過光合、呼吸、殘體分解等作用,可以對上覆水的DO、pH、氧化還原電位(Eh)以及水動力等產生復雜的影響,并通過這些影響因子的變化進而影響到磷素的遷移轉化[6-9]。黑藻等根系退化較為嚴重、頂冠蓬散的沉水植物主要是從上覆水中吸收營養物質,而苦草等根系較為發達的沉水植物則可同時從上覆水和沉積物中吸收營養物質。目前,沉水植物對于上覆水磷濃度影響主要是從水質凈化的角度進行了一些研究工作,王圣瑞等[10]采用蓬冠狀、根系退化的沉水植物黑藻,從沉積物-水-沉水植物系統的角度研究了沉水植物對上覆水中各形態磷濃度的影響。而對根系發達即深根系的沉水植物苦草研究報道較少。筆者在室內模擬條件下,研究根系發達的苦草在生長過程中對上覆水中各形態磷的變化的影響,以期為湖泊內源磷的控制或治理提供依據。
1 材料與方法
1.1 試驗設計
試驗在南京師范大學水環境生態修復中試平臺中進行,試驗日期為2008年8月中旬至11月下旬。苦草取自南京市固城湖,在室內馴化培養15 d。沉積物取自南京市玄武湖,試驗沉積物營養鹽質量比w(TN)為 3.832g/kg,w(TP)為 1.078g/kg,w(TOC)為39.580g/kg。試驗上覆水均是通過自來水充入。將沉積物分別過100目篩去除其中的植物殘體、石塊等雜質后,平鋪入試驗桶(底直徑×口直徑×高=45cm×55cm×80cm),厚度為10cm。然后選取每株有4~5葉、葉長40 cm、生長狀況基本一致、長勢良好的馴化后苦草,進行移栽,每桶30株苦草(鮮質量75g)。植物組和對照組各設3個平行。
1.2 監測指標與分析方法
試驗開始后即監測各項理化指標。DO、pH值、溫度等物理指標采用YSI水質在線監測儀監測;間隙水樣收集利用專門的滲濾裝置,原位滲濾后通過一次性采樣針管吸取。
TP濃度的測定:采用過硫酸鉀氧化消煮;DTP濃度的測定:將水樣預先過0.45μ m孔徑的醋酸纖維濾膜,而后用過硫酸鉀氧化消煮;SRP濃度的測定:將水樣預先過0.45μ m孔徑的醋酸纖維濾膜。經過以上預處理后TP、DTP、SRP均采用荷蘭SKALAR流動水質分析儀測定。PP濃度為TP濃度與DTP濃度之差,DOP濃度為DTP濃度與SRP濃度之差。
隨機測定本試驗已經馴化培養的苦草30株,構建植株體鮮質量與葉長、葉寬、葉片數的函數如下:
W=(0.023033x1+0.202922x2-0.25757)N
式中:W為植株鮮質量,g;x1為葉長,cm;x2為葉寬,cm;N為葉片數。
2 試驗結果
2.1 苦草的生長狀況
在本研究系統中,苦草均能正常生長。如圖1所示,種植的苦草經過約1周的適應,開始逐漸生長,且生長速度逐漸增加,并于試驗進行到約80d時,生物量達到峰值,此后,隨溫度的降低,苦草趨向于衰亡,生物量開始下降。
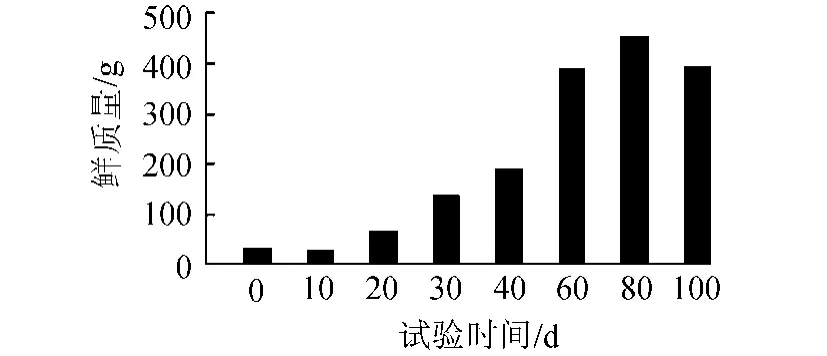
圖1 試驗期間苦草生物量(鮮質量)變化
2.2 上覆水中TP的變化
如圖2所示,苦草組與對照組中的TP均呈現先上升、后下降的趨勢。苦草組中的TP明顯低于對照組。上覆水中的 ρ(TP)在培養期間的變化范圍為0.01~0.19mg/L,平均值為0.094mg/L,其中苦草組上覆水的TP磷濃度為對照組TP濃度的62.5%。上覆水中TP濃度的變化取決于磷的收入和支出。試驗開始時,可能由于以自來水為水源使得底泥間隙水與上覆水磷存在較大的濃度差,以及植物對沉積物的擾動,上覆水中的TP表現為上升趨勢,峰值出現在20d左右;而后可能隨著間隙水與上覆水中磷的濃度差逐漸減小,且苦草開始迅速生長(圖1),對磷的需求量增大,上覆水磷的收入開始小于支出,表現為上覆水中TP下降。
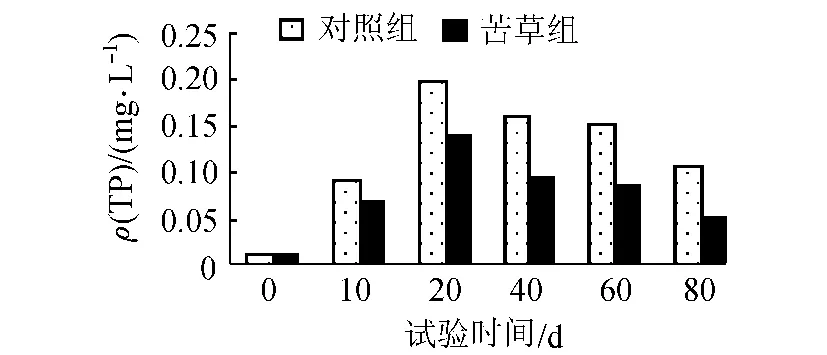
圖2 上覆水中 ρ(TP)的變化
2.3 上覆水中不同賦存形態磷的變化
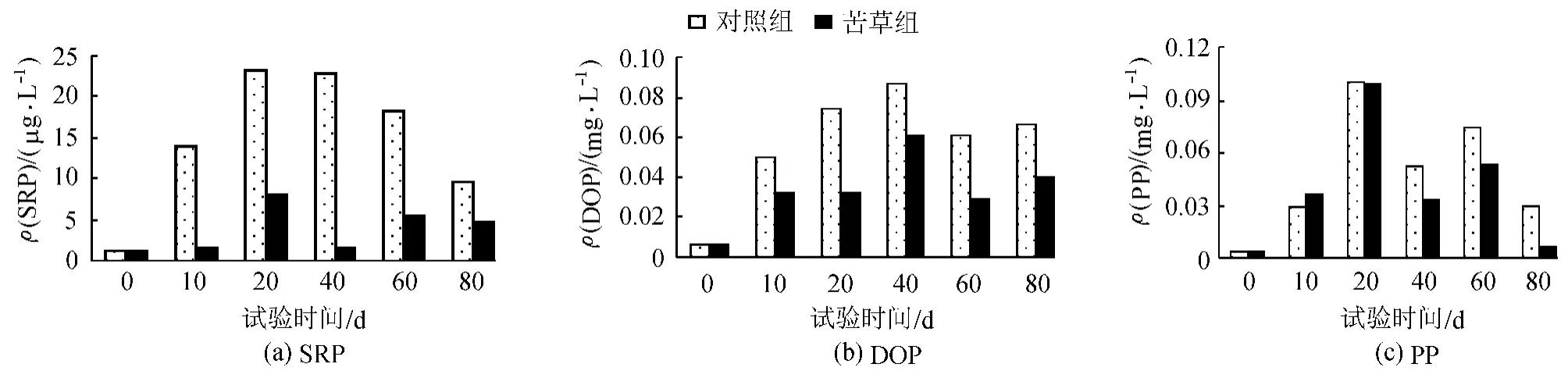
圖3 上覆水中不同P形態的變化
如圖3(a)所示,整個試驗期間,苦草組與對照組中的SRP都呈明顯的先上升后下降的趨勢,其峰值出現在20 d左右。苦草組中SRP的濃度值僅為對照組的25.4%;不同處理組的 ρ(SRP)變化范圍在1~23μ g/L,其中苦草組的 ρ(SRP)保持在 8μ g/L以下。不同處理組中的DOP基本上呈先上升后下降的變化趨勢,中間有小波動,其峰值均出現在40d左右,苦草組中DOP的濃度值僅為對照組的58.0%,ρ(DOP)的變化范圍在0.006~0.087mg/L。PP在對照組和苦草組也呈現了先上升后下降的趨勢,其峰值都出現在20d,苦草組中PP的濃度值為對照組的80.5%,ρ(PP)的變化范圍在 0.0035~0.1mg/L。在苦草種植初期,由于苦草還沒有完全適應環境,植株長勢較差,沉積物磷釋放起了最主要的作用。沉積物早期過程主要以釋放為主,上覆水中的各形態磷濃度上升到一定值后,然后下降。然而由于沉水植物苦草對不同形態磷影響的差異,導致各形態磷波動又有所不同。
由圖4可知,在本試驗條件下的沉積物上覆水系統中,TP中以有機磷為主,其所占比例的變化范圍在76.42%~98.40%,其中PP在整個試驗階段平均約占42.24%,DOP平均約占48%,SRP平均約占9.76%。在不同的采樣時間差異比較大。對于DOP和PP變化,苦草組大于對照組,而SRP的變化則是對照組大于苦草組。
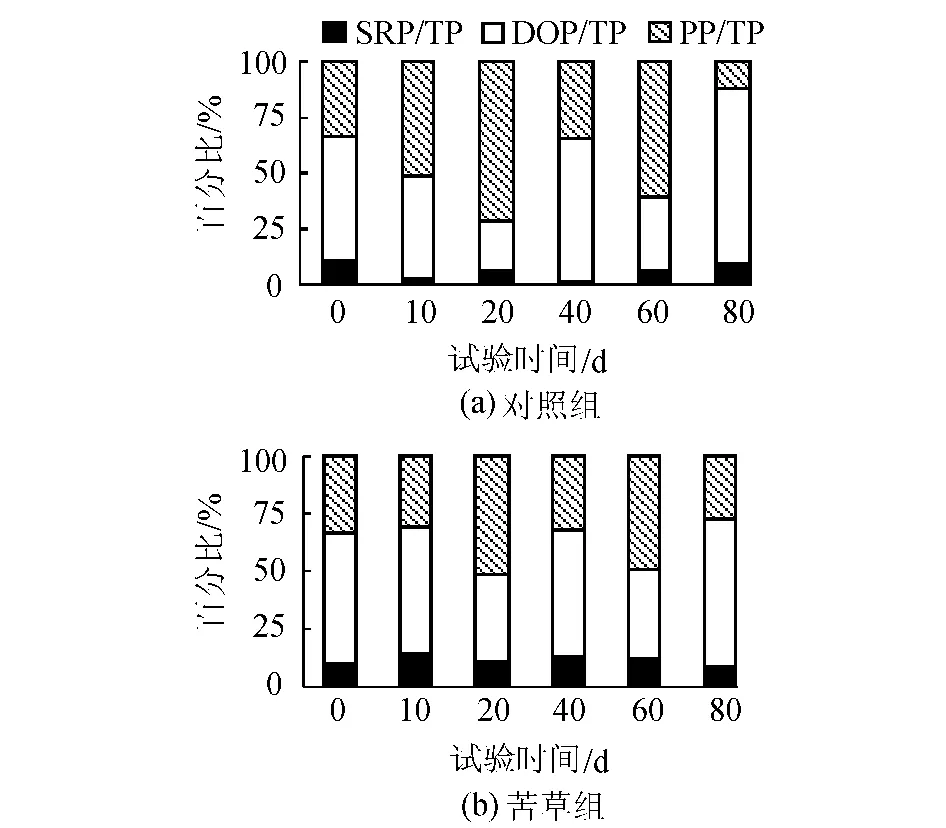
圖4 各形態P在不同培養時間不同處理下占TP的比例
3 討 論
3.1 苦草生長對沉積物磷釋放速率的影響
沉水植物的生長狀況以及生長特性會影響沉積物的釋放與吸收,同時也直接影響到上覆水中各形態磷的變化。為此,本研究采用相對釋放速率來定量描述各形態磷在其上升和下降階段的不同釋放速率。苦草處理中,將苦草和底質看作為一個統一的整體,來計算其向上覆水體釋放總磷的相對速率;對照組中,計算底質(沉積物)向上覆水體釋放總磷的相對速率[11]。計算公式如下:
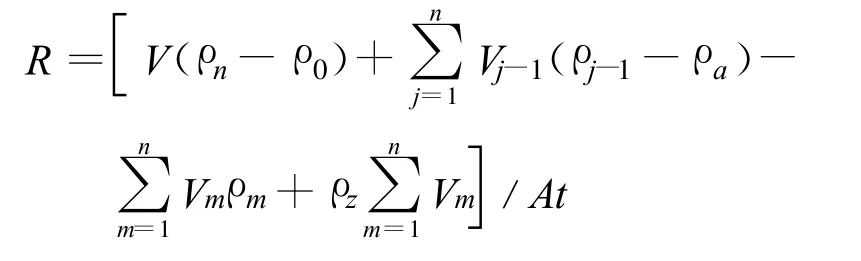
式中:R為磷釋放速率,mg/(m2?d);V為試驗桶中上覆水體積,L;ρn為第n次采樣時磷的質量濃度,mg/L;ρ0為磷初始值,mg/L;ρj-1為第j-1次采樣時磷的質量濃度,mg/L;Vj-1為第j-1次采樣體積,L;ρa為補償采樣量而添加自來水中磷的質量濃度,mg/L;Vm為第m次添加自來水體積,L;ρm為補償蒸發量而添加的第m次自來水中磷的質量濃度,mg/L;ρz為蒸餾水中磷的質量濃度,mg/L;A為水-沉積物界面接觸面積,m2;t為釋放時間,d。
如表1所示,在植物種植初期,由于植物還沒有完全適應環境,植物長勢較差,沉積物磷釋放起到最主要的作用,加上前期種植植物和加入上覆水對沉積物的擾動造成了沉積物磷的快速上升。對照組在早期過程主要以釋放為主且釋放相對速率高于苦草組,其中PP釋放速度最高。上覆水的磷濃度上升到一定值后,然后下降。在下降階段,不同處理組中各個形態磷的相對釋放速率均呈現負釋放,其中苦草組由于苦草生理代謝旺盛,對不同形態磷的吸收、吸附作用下導致其磷的負釋放速率(吸收速率)高于對照組。對照組中TP的負釋放主要由于顆粒態磷(PP)的沉降作用,SRP、DOP的負釋放可能由于其上覆水中藻類的吸收所致。整個試驗過程中苦草組上覆水的各形態磷濃度值均低于無沉水植物的對照組,這與通常認為沉水植物對水質具有凈化作用的結果一樣[12-13]。本試驗結果與王圣瑞等[10]對黑藻影響上覆水磷形態研究結果有所差異,可能原因是黑藻與苦草不同的生長特性與吸收釋放強度所致,同時也可能與所選取的沉積物不同有關。
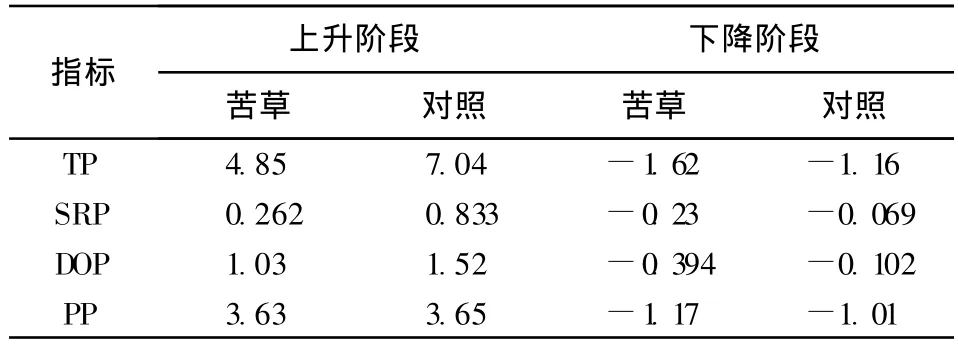
表1 各形態磷的相對釋放速率 mg/(m2?d)
一般來說,湖泊中的總磷絕大部分為有機磷(>90%),其中70%以上是懸浮態顆粒有機磷,剩下部分是溶解態及膠體態有機磷。而作為能被生物直接利用的無機態的溶解性正磷酸鹽,其比例較小,不足湖水總磷的5%[14]。而本試驗中PP(懸浮態顆粒磷)所占比例比自然水體要小,DOP為48%、SRP為9.76%,所占比例比自然水體高,這與本試驗在室內進行、水體環境受外界干擾較小有關。
3.2 環境因子對上覆水磷濃度的影響
水體中的DO、pH值、溫度等環境因子是影響沉積物磷釋放的重要因素[15]。本試驗中,苦草通過影響試驗水體中的DO、pH值,進而間接影響到底泥磷的釋放速率,最后可能導致上覆水各形態磷的差異。如圖5(a)、(b),苦草組中的DO由于植物的光合作用釋放氧氣而明顯高于對照組;同樣的,由于植物吸收了CO2,苦草組的pH值小于對照組。水體中較高的溶解氧能提高水-沉積物界面的氧化還原電位(Eh),Eh的升高,導致Fe3+與磷酸鹽結合成不溶的磷酸鐵,可溶性磷也被氫氧化鐵吸附而逐漸沉降,于是降低了底泥磷的釋放速率[16]。pH值的升高,在弱堿條件下,沉積物輕微吸收磷,吸收量和吸收速率隨pH值升高而升高[17],如圖2、3所示,苦草組中上覆水中的TP、SRP、DOP的濃度均小于對照組,這與DO、pH值的影響不無關系。試驗期間的環境溫度如圖5(c),前期溫度一直持續在27~28℃左右,后期降至14~15℃,而較高溫度會導致沉積物磷的釋放,較低溫下,水體中的磷向沉積物沉降[18]如圖3,這與上覆水中各形態磷,尤其是與PP磷前期升高、后期下降的變化趨勢是一致的。
3.3 間隙水對上覆水各形態磷濃度的影響
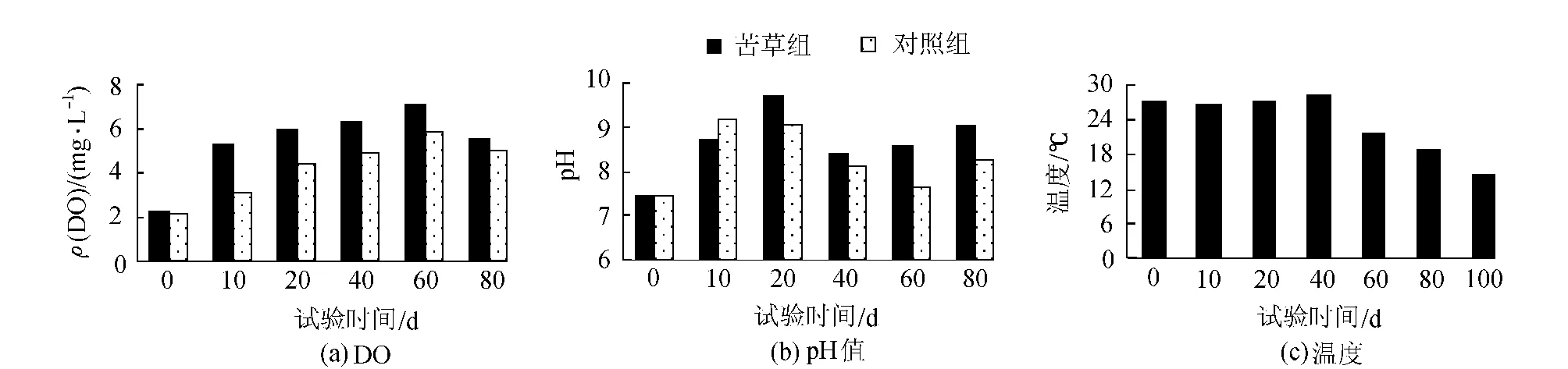
圖5 上覆水中DO、pH值及溫度的變化
如圖5所示,間隙水中各形態磷濃度隨培養時間的變化較為明顯。比較圖3與圖6結果可見,總體上來講,整個試驗期間各處理間隙水中各形態磷濃度均高于上覆水,這表明了在整個試驗期間各形態磷在上覆水與間隙水間存在較為明顯的濃度梯度。苦草組間隙水中各形態磷濃度均低于對照處理組,這是由于苦草主要通過根部吸收間隙水中的可溶性磷[19],間隙水中DTP與SRP濃度與上覆水中DTP、SRP濃度的變化呈顯著正相關(R2=0.75)。間隙水磷濃度的變化直接影響上覆水磷的濃度。在試驗初期,苦草組間隙水中的DTP和SRP濃度為增加趨勢并在40d左右達到峰值,而后為下降趨勢。這可能與當時的環境溫度較高(如圖5(c)所示)、沉積物磷釋放起了最主要的作用有關[20]。此后,TDP、SRP濃度降低,其原因可能為環境溫度的降低,減弱了微生物解磷活性[21],導致沉積物中釋放磷的減少進而使間隙水中磷濃度表現為下降趨勢。
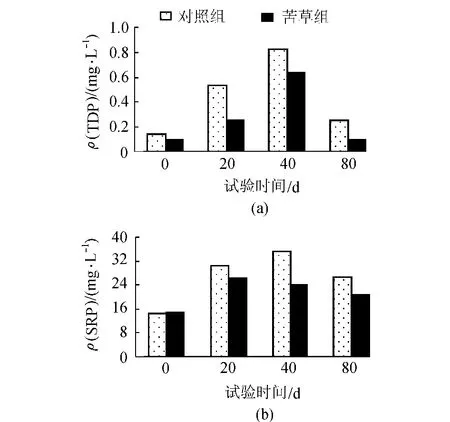
圖6 間隙水中不同磷形態的變化
4 結 語
本試驗條件下,沉水植物苦草對上覆水各形態磷影響明顯。在苦草生長過程中因其對磷營養的需求和對環境因子DO、pH值的影響減緩了沉積物磷的釋放速率,使得上覆水中TP、SRP 、DOP、PP均呈現不同程度的下降趨勢,其濃度值小于無沉水植物的對照組。上覆水各形態磷以DOP為主,這與本試驗在室內進行,受外界干擾效小有關。間隙水對上覆水各相態磷濃度的影響主要與環境溫度的變化以及苦草根部直接吸收間隙水中的磷有關。
[1] ISEMANN K.Share of agriculture in nitrogen and phosphorous emissions into surface waters of Western Europe against the background of their eutrophication[J].Fertilizer Research,1990,26:253-269.
[2] DAVID L C.The role of phosphorus in the eutrophication of receiving water:a review[J].Journal of Environment Quality,1998,27:261-266.
[3] 吳重華,王曉蓉,孫昊.羊角月牙藻的生長與湖泊水中磷形態關系的建立[J].環境化學,1997,16(4):341-346.
[4] 周易勇,付永清.水體磷酸酶:來源、特征及生態學意義[J].湖泊科學,1999,11(3):274-282.
[5] 劉健康.高級水生生物學[M].北京:科學出版社,1999:224-240.
[6] PETTICREW E L,KALFF J.Water flow and clay retention in submergedmacrophyte beds[J].Can J Fish Aquat Sci,1992,49:2483-2489.
[7] STEPHEN D,MOSS B,GEOFF P.Do rooted macrophytes increase sediment phosphorus release?[J].Hydrobiologia,1997,342/343:27-34.
[8] HORPPILA J,NURMINEN L.Effects of different macrophyte growth forms on sediment and P resuspension in a shallow lake[J].Hydrobiologia,2005,545(1):167-175.
[9] 胡俊,豐民義,吳永紅,等.沉水植物對沉積物中磷賦存形態影響的初步研究[J].環境化學,2006,25(1):28-31.
[10] 王圣瑞,金相燦,趙海超,等.沉水植物黑藻對上覆水中各形態磷濃度的影響[J].地球化學,2006,35(2):179-186.
[11] 范成新,張路,楊龍元,等.湖泊沉積物氮磷內源負荷模型[J].海洋與湖沼,2002,33(4):371-377.
[12] 吳振斌,邱東茹,賀峰,等.水生植物對富營養化水體水質凈化作用研究[J].武漢植物學研究,2001,19(4):299-303.
[13] 高鏡清,熊治延,張維昊,等.常見沉水植物對東湖重度富營養化水體磷的去除效果[J].長江流域資源與環境,2007,16(6):796-800.
[14] WETZEL R G.Limnology,2nd ed.[M].Philadelphia:Saunders Collage Publishing,1983.
[15] 黎穎治,夏北成.影響湖泊沉積物-水界面磷交換的重要環境因子分析[J].土壤通報,2007,38(1):162-166.
[16] 吳曉磊.人工濕地廢水處理機制[J].環境科學,1995,16(3):83-86.
[17] 耿金菊,王強,牛曉君,等.環境因子對湖泊沉積物中吸附態磷化氫生成和釋放的影響[J].環境科學學報,2005,25(5):681-685.
[18] JENSEN H S,ANDERSEN F O.Importance of temperature,nitrate and pH for phosphorus release from aerobic sediment of four shallow,eutrophic lake[J].Limnol Oceanogr,1992,37(3):577-589.
[19] WILHELM G,DORIS S.Influence of aquatic macrophytes on phosphorus cycling in lakes[J].Hydrobiologia,1988,170:245-266.
[20] S?NDERGAARD M,JENSEN J P,JEPPESEN E.Role of sediment and internal loading of phosphorus in shallow lakes[J].Hydrobiologia,2003,506-509,135-145.
[21] KIM Lee-Hyung,Choi E,STENSTROM M K.Sediment characteristics,phosphorus types and phosphorus release rate between river and lake sediments[J].Chemosphere,2003,50(1):53-61.
