含鎘廢水人工濕地處理系統中基質微生物數量和酶活性研究
2010-08-03 08:32:02張超蘭韋必帽李忠義
水資源保護 2010年4期
關鍵詞:植物
張超蘭,韋必帽,劉 敏,李 磊,李忠義,高 暢
(廣西大學農學院,廣西南寧 530005)
人工濕地凈化廢水的原理主要是利用濕地中基質、植物和微生物之間的相互作用,通過一系列物理的、化學的以及生物的途徑凈化污水[1-2]。其中基質微生物和酶在污水凈化過程中起了非常重要的作用。各種酶在基質中的積累是基質微生物、動物區系和植物根系生命活動的結果,它們參與了基質中腐殖質的合成與分解,有機化合物、高等植物和微生物殘體的水解及其轉化成為可利用的形態、以及氧化還原反應等[3]。重金屬鎘對植物的生理具有脅迫作用[4],對微生物和酶活性的影響已有相關報道。同樣,人工濕地植物、基質、微生物和酶在重金屬污染水體的生態修復中扮演重要的角色。筆者通過對復合垂直流人工濕地處理無機鎘廢水的基質微生物數量和酶活性的研究,分析其與去除重金屬鎘的關系,探討人工濕地去除污水中重金屬的機理。
1 材料和方法
1.1 復合垂直流人工濕地系統
試驗在廣西大學農業資源與環境科學系教學實習基地上建立下行-上行流為主要單元的垂直流人工濕地系統[5],該系統由下行流、上行流2個單元串聯而成,各單元大小為120cm×83cm×70 cm,自下而上分別填充礫石(?16~32mm)、豆石(?8~16 mm)、粗沙(?4~6mm)和細沙(?0~4mm)4種基質。植物配置分別設:①美人蕉(下行池)+紅蛋(Echinodorus osiris)(上行池)(簡寫為MH);②彩葉草(下行池)+風車草(上行池)(簡寫為CF);③無植物系統(CK)。復合垂直流人工濕地系統結構如圖1。
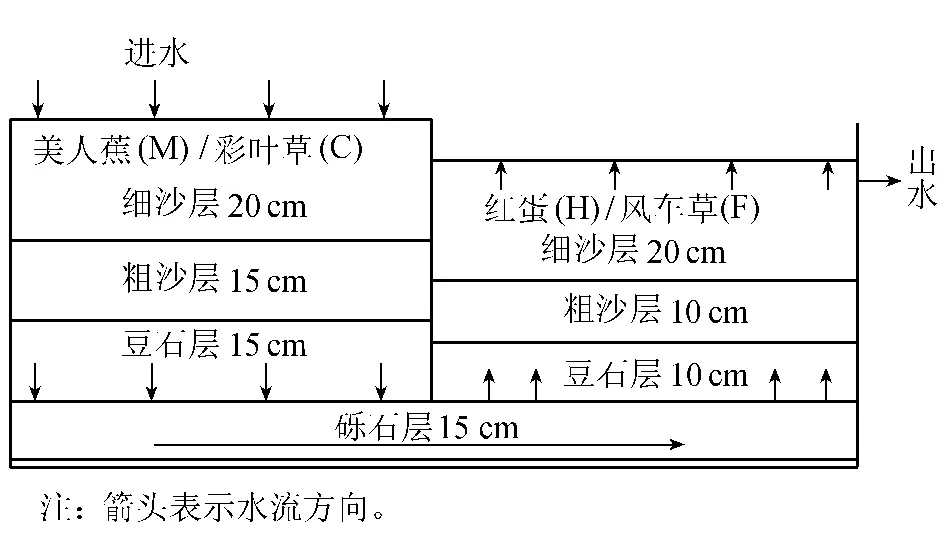
圖1 復合垂直流人工濕地系統結構示意圖
試驗用水用自來水配制,進水的鎘質量濃度為29~ 33 mg/L,電導率為 349~ 417μ S/cm,用濃度為5mol/L的HCl溶液調pH值至6.21~6.45。進水方式為間歇式,2007年9~11月每月進水運行一次,運行時間為24h,運行3個月,水力負荷為600mm/d。
1.2 樣品的采集
采樣時,先讓出水口水流2min后再采樣,測定重金屬Cd的濃度;在運行后第8天采集基質樣,把下行池、上行池中基質從上至下分為0~5 cm、5~15cm、15~20cm 3層,分別采用梅花點法分層取樣,然后把同層次基質充分混勻,分成兩部分,一部分鮮樣過2mm篩,保存于4℃冰箱待測微生物數量。另一部分風干,磨碎,過1mm篩,待測各種酶的活性。
1.3 微生物數量的測定
基質的細菌、真菌、放線菌采用平板計數法測定,分別用牛肉膏蛋白胨瓊脂、改良馬丁氏培、淀粉銨鹽培養基測定[6]。
1.4 酶活性的測定
脲酶測定采用苯酚-次氯酸鈉比色法,蔗糖酶采用3,5二硝基水楊酸比色法[7]。
2 結果與分析
2.1 植物根系分布特點
人工濕地4種植物根系分布特點各不相同,美人蕉根量最大,其塊根盤錯在人工濕地基質的表面,且須根發達,根系深度主要在0~10cm;紅蛋主要以須根為主,在濕地中其根系分布深度較美人蕉深,主要分布在0~20cm;彩葉草的根系也都是須根,根淺且根系相對細小;風車草根系較粗且較深,至40cm的深度含量也較大,表層相對少量分布。
2.2 基質微生物數量及其空間分布
由表1可見,不同植物濕地系統的細菌數量明顯不同,其中,MH與CK之間細菌數量差異顯著(P<0.05),但是MH與CF、CF與CK之間差異都不顯著;上行池中基質細菌數量極顯著大于下行池(P<0.01),從細菌數量的垂直空間分布看,總體規律性差異不顯著(P>0.05)。
人工濕地基質中真菌數量較細菌數量少,在104~106個/g之間。真菌在系統中受植物影響比較大,其不同植物系統真菌數量大小順序也是MH、CF、CK。其中MH與CK、CF與CK之間的差異性達到極顯著水平(P=0.0002<0.01,P=0.0327<0.05);但是,MH與CF之間的差異不顯著(P=0.201>0.05)。真菌在下行池中的數量極顯著大于上行池的數量(P=0.0059<0.01);從真菌的垂直分布看,總體規律是0~5cm>5~15cm>15~20cm,在 0~5cm基質的真菌極顯著高于5~15 cm、15~20 cm(P<0.01),而5~15cm與15~20cm之間的差異性不顯著(P>0.05)。
放線菌與真菌在垂直流人工濕地基質中的規律基本一致,其數量也基本維持在104~106個/g的水平,但是各處理之間放線菌數量差異不顯著性,上行池與下行池之間放線菌數量差異也不顯著;其垂直分布總體規律是0~5cm>5~15cm>15~20cm,0~5cm與5~15cm中放線菌數量達到顯著性差異(P<0.05),0~5cm與15~20cm中放線菌數量差異則達到極顯著水平(P=0.0009<0.01)。
2.3 上層基質微生物數量與鎘去除率相關性
人工濕地基質上層(0~5cm)微生物數量與廢水中鎘去除率的相關性分析結果(表1和表2)表明,在下行池中,上層真菌和放線菌數量與去除率相關性均呈顯著正相關(r=0.778,P=0.014<0.05;r=0.789,P=0.011<0.05),而細菌數量與鎘去除率相關性不顯著(P>0.05);在上行池中,真菌數量與鎘去除率相關性顯著正相關(r=0.682,P=0.042<0.05);而細菌和放線菌數量與鎘去除率相關性都不顯著(P>0.05)。
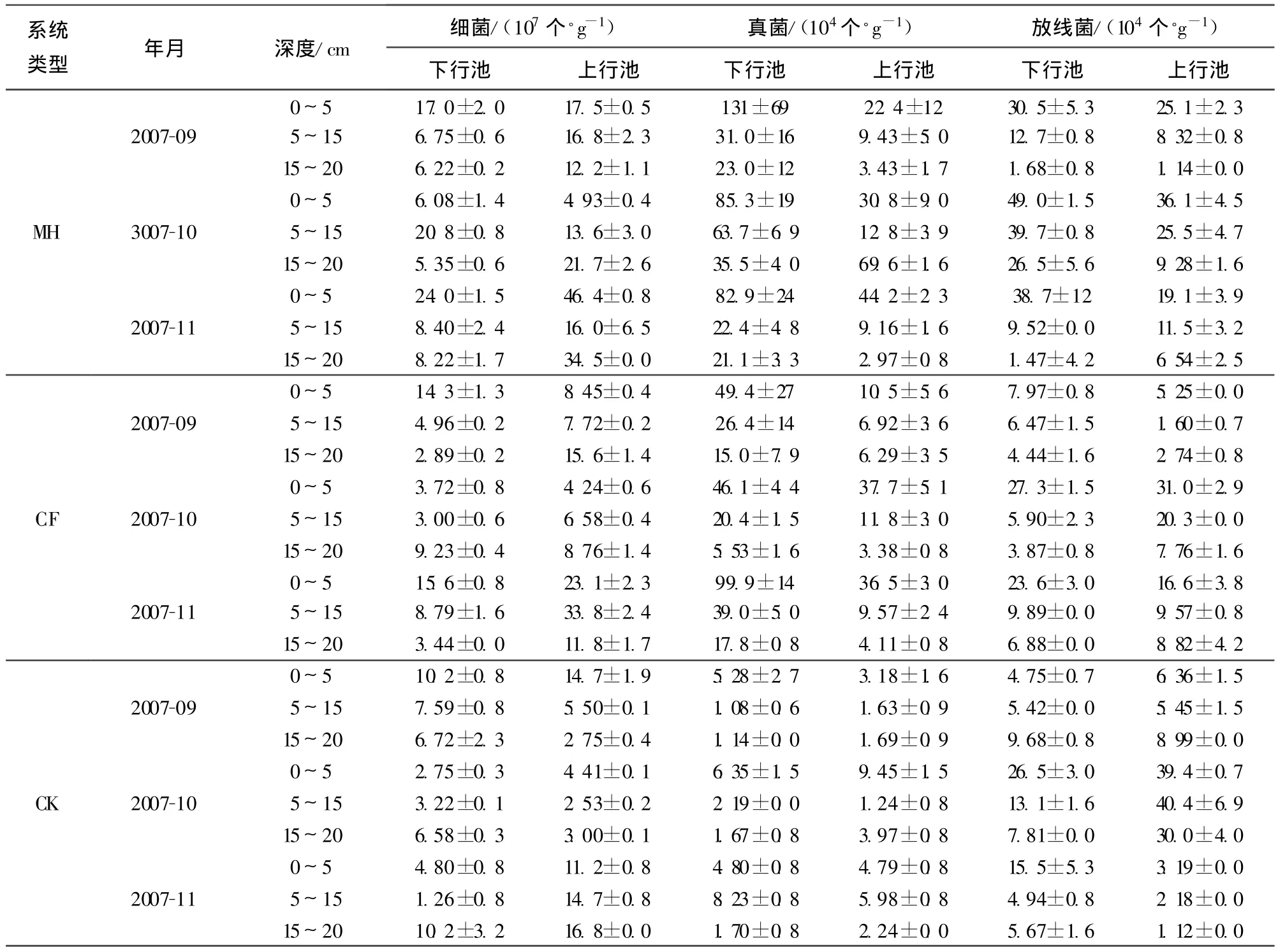
表1 基質微生物數量及其空間分布
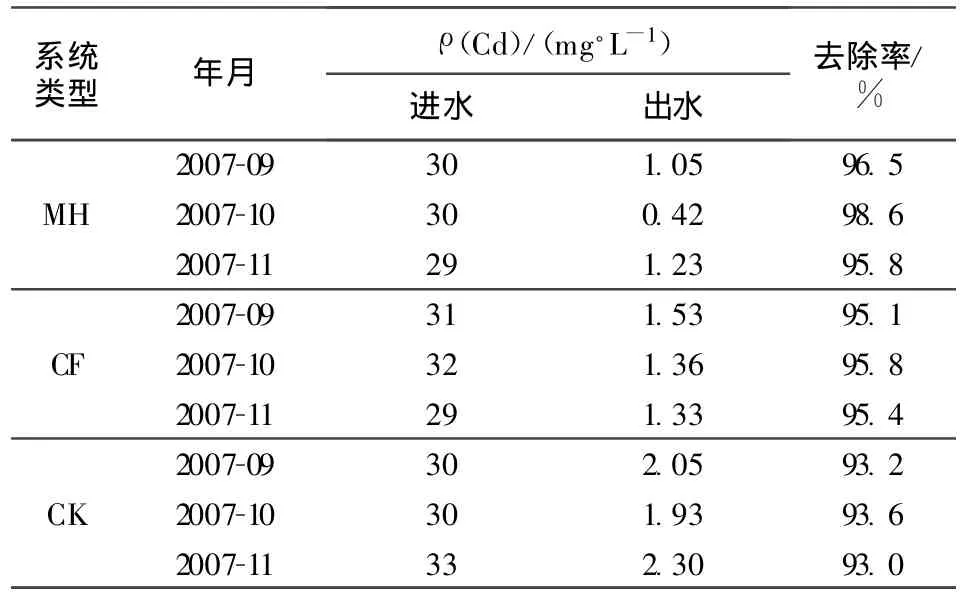
表2 廢水中鎘的去除率
2.4 基質酶活性及其空間分布
基質脲酶活性與基質微生物數量、有機物質濃度、總氮和速效氮濃度有關,在不同植物的人工濕地系統中,脲酶活性的大致規律為:MH>CK>CF,MH中脲酶活性極顯著高于CK和CF(P=0.0001<0.01),而CK與CF差異不顯著。基質中脲酶的活性隨深度的增加而下降,在下行池中,其中0~5cm中脲酶活性極顯著高于15~20cm的(P=0.0392<0.05),而其余各層之間差異性都未達到顯著水平。上行池脲酶活性的垂直分布各層間的差異性都不顯著(P>0.05)。
濕地基質蔗糖酶來自植物根系和濕地微生物,其活性受植被影響較大。由表3可以看出,不同植物系統濕地基質蔗糖酶的活性大小順序:MH>CF>CK,且MH與CF、MH與CK、CF與CK之間差異都達到極顯著水平(P<0.01);垂直空間分布上,蔗糖酶的活性隨深度增加而呈下降趨勢,但各層間的差異性都不顯著(P>0.05)。
2.5 上層基質酶活性與鎘去除率相關性
根據表2和表3,對濕地上層(0~5 cm)基質酶活性與鎘去除率做相關性分析,結果發現,無論是上行池還是下行池中,上層基質中蔗糖酶活性與鎘去除率相關性都呈極顯著正相關(上行池r=0.863,P=0.003<0.01;下行池 r=0.837,P=0.005<0.01);而脲酶活性與鎘去除率相關性均未達到顯著水平(P>0.05)。
3 討 論
a.微生物抗重金屬機制有生物吸附作用和細胞外的沉淀結晶作用[8],許多微生物能產生大量的胞外多糖,胞外多糖帶有負電荷,作為重金屬的有效吸附,其分泌的有機酸對重金屬也有絡合作用;有些微生物能參與形成沉積物,這些沉積物含有大量的重金屬,從而達到去除重金屬的作用。在人工濕地處理無機鎘廢水中,細菌在上行池的數量大于下行池,這表明細菌數量除了與植物根系有關外,還可能與處理過程中鎘廢水濃度有關,因為經過下行池基質和植物根系的吸附凈化作用,當廢水流經上行池的時候,其濃度應有所下降,這樣對細菌的毒害作用降低。有研究表明,細菌受重金屬鎘濃度影響較大,在三大菌中最敏感[9-10]。細菌、真菌和放線菌在濕地基質中垂直分布都是0~5 cm>5~15cm>15~20cm,有植物系統微生物數量顯著大于無植物系統,與Ottova等[11]、Machate等[12]和成水平等[13]的研究結果一致,即人工濕地微生物數量越大,對重金屬鎘去除效果也越好。

表3 基質酶活性空間分布
b.導致土壤酶活性降低的主要因素是重金屬含量[14],重金屬對酶活性的抑制機理可能與酶分子中的活性部位一巰基和含瞇唑的配位等結合,形成較穩定的絡合物,產生了與底物的競爭性抑制作用有關,或者可能由于重金屬通過抑制土壤微生物的生長和繁殖,減少體內酶的合成和分泌,最后導致土壤酶活性下降[15-16]。脲酶受重金屬濃度影響不大[15],這與本試驗脲酶與鎘去除率相關性不顯著結論一致。人工濕地中植物的根系發達,微生物群落豐富,是影響酶活性的重要因素。試驗中,美人蕉系統表層脲酶活性最高,即0~5cm脲酶活性最高;有植物系統極顯著高于無植物系統,這與黃娟等[17]的研究結果一致。
4 結 論
a.在復合垂直流人工濕地系統中,3種微生物中細菌數量最多(為106~108個/g數量級),真菌和放線菌相差不大(都在104~106個/g數量級)。
b.不同植物配置的復合垂直流人工濕地基質微生物數量和蔗糖酶活性大小順序為:MH>CF>CK;基質細菌數量為上行池大于下行池,而真菌、放線菌數量和酶活性則為下行池高于上行池;微生物數量和酶活性的垂直空間分布都是在0~5cm極顯著高于其他各層。
c.無論在下行池還是上行池,基質上層(0~5 cm)中,真菌數量和蔗糖酶活性與鎘去除率均呈極顯著正相關,而細菌數量和脲酶活性與鎘去除率的相關性都不顯著;放線菌數量與鎘去除率的相關性則表現為,在下行池放線菌數量與鎘去除率呈顯著正相關,而在上行池相關性則不顯著。可見,真菌數量和蔗糖酶活性是影響垂直流人工濕地去除廢水中鎘的主要因素,而真菌數量和蔗糖酶活性與濕地植物的關系密切。
[1] HAMMER D A.Constructed wetlands for wastewater treatment municipal,industrialand agricultura1.Proc.1stinternal conference on constructed wetlands for wastewater treatment[M].Chattanooga,USA,Lewis Publishing,Chelsea,1988.
[2] MARTIN C D,MOSHIRI G A.Nutrient reduction in an series constructed wetland system treating landfill leachate[J].Water Science and Technology,1994,29(4):267-272.
[3] 李智,楊在娟,岳春雷.人工濕地基質微生物和酶活性的空間分布[J].浙江林業科技,2005,25(3):1-5.
[4] 張超蘭,陳文慧,韋必帽,等.幾種濕地植物對重金屬鎘脅迫的生理生化響應[J].生態環境,2008,17(4):1458-1461.
[5] 張洪剛,馬安娜,洪劍明.垂直流人工濕地設計及凈化功能初探[J].節水灌溉,2006(6):31-34.
[6] 許光輝,鄭洪元.土壤微生物分析方法手冊[M].北京:農業出版社,1986.
[7] 姚槐應,黃昌勇.土壤微生物生態學及其實驗技術[M].北京:科學出版社,2006.
[8] 池振明.現代微生物生態學[M].北京:科學出版社,2005.
[9] BAATH E.Effects of heavy metals in soils on microbial processes and populations:a review[J].Water Air Soil Pollut,1989,47:335-379.
[10] 龍健,黃昌勇,滕應,等.礦區廢棄地土壤微生物及其生化活性[J].生態學報,2003,23(3):492-503.
[11] OTTOVD V,BALCAROVOJ,VYMAZALJ.Microbial characteristics of constructed wetlands[J].Water Science and Technology,1997,35(5):117-123.
[12] MACHATE T,NOLL H,BEHRENS H,et a1.Degradation of phenan-threne and hydraulic characteristics in a constructed wetland[J].Water Research,1997,31(3):554-560.
[13] 成水平,夏宜垮.香蒲、燈心草人工濕地的研究Ⅲ:凈化污水的機理[J].湖泊科學,1998,10(2):66-71.
[14] 滕應,黃昌勇,龍健,等.鉛鋅銀尾礦污染區土壤酶活性研究[J].中國環境科學,2002,22(6):551-555.
[15] 張玲,葉正錢,李廷強,等.鉛鋅礦區污染土壤微生物活性研究[J].水土保持學報,2006,20(3):137-140.
[16] KANDELER E,LUFTENEGGER G,SCHWARZ S.Influence of heavy metals on the functional diversity of soil microbial communities[J].Biology and Fertility of Soils,1997,23:299-306.
[17] 黃娟,王世和,鄢璐,等.潛流型人工濕地的脲酶活性分布特征[J].東南大學學報:自然科學版,2008,38(1):166-169.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
