RunX3對(duì)小鼠骨髓造血干細(xì)胞功能的影響
2010-09-08 08:14:18石桂英陳陟陽(yáng)胡永艷鞠振宇
中國(guó)比較醫(yī)學(xué)雜志 2010年4期
關(guān)鍵詞:小鼠
姚 超,石桂英,陳陟陽(yáng),黃 馨,胡永艷,鞠振宇
(中國(guó)醫(yī)學(xué)科學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物研究所 北京協(xié)和醫(yī)學(xué)院,衛(wèi)生部人類疾病比較醫(yī)學(xué)重點(diǎn)實(shí)驗(yàn)室,北京 100021)
研究報(bào)告
RunX3對(duì)小鼠骨髓造血干細(xì)胞功能的影響
姚 超,石桂英,陳陟陽(yáng),黃 馨,胡永艷,鞠振宇
(中國(guó)醫(yī)學(xué)科學(xué)院醫(yī)學(xué)實(shí)驗(yàn)動(dòng)物研究所 北京協(xié)和醫(yī)學(xué)院,衛(wèi)生部人類疾病比較醫(yī)學(xué)重點(diǎn)實(shí)驗(yàn)室,北京 100021)
目的 研究 RunX3基因?qū)υ煅杉?xì)胞自我更新和分化能力的影響。方法流式細(xì)胞術(shù)測(cè)定小鼠骨髓干細(xì)胞和外周血單個(gè)核細(xì)胞的比例;通過(guò)競(jìng)爭(zhēng)性骨髓移植實(shí)驗(yàn)檢測(cè) RunX3轉(zhuǎn)基因小鼠骨髓干細(xì)胞的功能。結(jié)果移植后來(lái)源于 RunX3-/-小鼠骨髓干細(xì)胞供體的外周血細(xì)胞占總外周血細(xì)胞的比例與野生對(duì)照鼠相比無(wú)明顯差異,移植后來(lái)源于 RunX3-/-小鼠骨髓干細(xì)胞供體的外周血中髓系細(xì)胞占總外周血髓系細(xì)胞的比例較野生型對(duì)照鼠高。結(jié)論RunX3基因缺失對(duì)骨髓造血干細(xì)胞的自我更新沒有影響,但其可能參與了骨髓造血干細(xì)胞的分化過(guò)程。
R-33;小鼠,基因敲除;骨髓造血干細(xì)胞
RunX即編碼具有Runt結(jié)構(gòu)域蛋白的基因,其家族成員包括RunX1、RunX2和RunX3,它們都有一個(gè)DNA結(jié)合結(jié)構(gòu)域 runt,該結(jié)構(gòu)域與果蠅的成對(duì)控制(pair rule)基因 runt序列同源。作為 RunX家族的重要成員,RunX3位于人染色體1p36.1區(qū)域,至少具有5個(gè)外顯子。在小鼠胚胎形成過(guò)程中RunX3表達(dá)于造血器官、表皮附屬物、骨骼以及感覺神經(jīng)中樞[1]。RunX3是胃上皮細(xì)胞主要生長(zhǎng)調(diào)控因子,RunX3敲除小鼠由于上皮細(xì)胞的過(guò)度增殖和凋亡抑制表現(xiàn)為胃粘膜增生。RunX3在TGF-β介導(dǎo)的腫瘤抑制信號(hào)途徑中起重要作用,該基因功能的缺失導(dǎo)致胃癌的發(fā)生和進(jìn)展[2,3]。RunX3和RunX1基因在抑制 CD4表達(dá)方面具有協(xié)同性[4]。RunX3基因敲除導(dǎo)致加速成熟,對(duì) T細(xì)胞刺激朗格漢斯細(xì)胞缺乏和樹突細(xì)胞的加強(qiáng)以及使β2整合素家族如CD11a、CD11b、CD11c的表達(dá)異常[5]。綜合前期研究結(jié)果提示RunX3可能與細(xì)胞增殖、凋亡、分化有關(guān),對(duì)免疫系統(tǒng)疾病[6]和癌癥的發(fā)生有影響。然而,目前還沒有研究報(bào)道RunX3和造血干細(xì)胞的自我更新能力以及應(yīng)激反應(yīng)之間的關(guān)系。為了研究RunX3是否直接參與骨髓造血干細(xì)胞本身的自我更新能力的基因調(diào)控,我們利用競(jìng)爭(zhēng)性骨髓造血干細(xì)胞移植的方法來(lái)研究RunX3基因在造血干細(xì)胞自我更新和分化中的功能。
1 方法
1.1 競(jìng)爭(zhēng)性骨髓移植實(shí)驗(yàn)
供體小鼠:RunX3-/-小鼠為12周齡;對(duì)照組小鼠采用 C57BL/6野生型,為 12周齡。受體小鼠: C57BL/6野生型,為8周齡。競(jìng)爭(zhēng)者小鼠:為CD45. 1亞型C57BL/6小鼠,均來(lái)自北京協(xié)和醫(yī)學(xué)院比較醫(yī)學(xué)中心,動(dòng)物生產(chǎn)許可證號(hào):SCXK(京)2009-0007,動(dòng)物使用許可證號(hào):SYXK(京)2005-0001。受體小鼠在接受骨髓移植前4 h接受致死劑量的放射線照射,劑量率0.5~1 Gy/min,總劑量達(dá)9 Gy。無(wú)菌條件下制備RunX3-/-小鼠和野生型對(duì)照小鼠的骨髓單個(gè)核細(xì)胞作為移植供體,與競(jìng)爭(zhēng)者小鼠骨髓單個(gè)核細(xì)胞1:1混合,將等量的細(xì)胞懸液(約200~300 μL,含單個(gè)核骨髓細(xì)胞約4×106個(gè))經(jīng)眼后靜脈注射到的各組受體小鼠體內(nèi)。實(shí)驗(yàn)所用小鼠飼養(yǎng)在SPF級(jí)動(dòng)物飼養(yǎng)室 ,移植后1周給予受體小鼠口服抗生素預(yù)防感染,并注意飲食飲水補(bǔ)給,以提高移植成活率。
1.2 5-氟尿嘧啶移植后再處理
通過(guò)腹腔注射給予移植后5月的受體小鼠150 mg/kg的5-氟尿嘧啶(5-FU)。使得骨髓中休眠的造血細(xì)胞重新開始造血。在給予5-FU后7 d和1月時(shí)取外周血分析供體來(lái)源的血細(xì)胞比例[7]。
1.3 PCR方法鑒定RunX3-/-小鼠的基因型
提取小鼠基因組DNA,PCR鑒定基因型。反應(yīng)條件:95℃變性5 min;95℃ 30 s,64℃ 30 s,72℃30 s,30個(gè)循環(huán);72℃延伸10 min。鑒定引物為上游5′-GAGATAGATGGGGTCAGAGG-3′,下游5′-AGG TCGGGTCTAGAGAATAGG-3′(Invitrogen),RunX3-/-產(chǎn)物長(zhǎng)度300 bp,WT產(chǎn)物長(zhǎng)度200 bp,PCR試劑購(gòu)自大連寶生物工程有限公司(圖1)。

注:Marker為DL2000 marker,H20為空白對(duì)照,WT為野生型對(duì)照,1,2,3為RunX3-/-。
1.4 流式細(xì)胞術(shù)分析
我們對(duì)競(jìng)爭(zhēng)性移植的供體小鼠的脾臟,胸腺,骨髓細(xì)胞進(jìn)行分析。并在移植后1月,3月分析受體小鼠外周血中供體來(lái)源細(xì)胞的嵌合率。將脾臟、胸腺、骨髓細(xì)胞分別制成單個(gè)細(xì)胞混懸液,懸浮于 staining medium(2.5% FBS在 PBS中),供體脾臟、胸腺、骨髓細(xì)胞采用 CD4-APC、CD8-FITC、B220-PE、CD11b-Percp-cy5.5,并冰上孵育30 min。流式細(xì)胞儀采用FASCAria(Becton Dickinson)。骨髓細(xì)胞同時(shí)還采用 LSK分析,lineage marker antibodies-biotin(biotinylated anti-Ter-119抗體、biotinylated anti-Gr-1抗體、biotinylated anti-CD11b抗體、biotinylated anti-B220抗體、biotinylated anti-IL-7R 抗體、biotinylated anti-CD4抗體、biotinylated anti-CD8抗體)、SA-Percpcy5.5、Sca-1-PE及 ckit-APC。
移植后外周血嵌合率分析:通過(guò)眼后靜脈叢取外周血50~100 μL,采用0.1 mol/L EDTA20 μL抗凝,Redbloodcell(RBC)lysis buffer(0.83% NH4Cl)裂解紅細(xì)胞。采用 CD45.1-PE、CD45.2-Percp-cy5.5、B220-FITC、CD11b-APC分別標(biāo)記供受體來(lái)源的B淋巴細(xì)胞和髓系細(xì)胞,并計(jì)算供體來(lái)源的外周血細(xì)胞比例。
1.5 統(tǒng)計(jì)學(xué)分析
數(shù)據(jù)結(jié)果均用x±s表示,組間資料比較采用Student t檢驗(yàn)。
2 結(jié)果
2.1 RunX3-/-小鼠的造血系統(tǒng)分析
為了解RunX3基因的缺失對(duì)年輕小鼠(12周)造血系統(tǒng)所造成的影響,我們分析了 RunX3-/-小鼠和野生型對(duì)照小鼠的骨髓造血的情況(圖2A,2B)。結(jié)果顯示RunX3基因缺失不但對(duì)小鼠的骨髓B淋巴細(xì)胞和髓系細(xì)胞的發(fā)育沒有影響(圖2C)。這些數(shù)據(jù)表明 RunX3基因?qū)δ贻p小鼠的骨髓造血系統(tǒng)向B淋巴細(xì)胞和髓系細(xì)胞分化的能力沒有影響。進(jìn)而,我們分析了RunX3基因?qū)δ贻p小鼠骨髓造血干細(xì)胞數(shù)量的影響,結(jié)果顯示RunX3敲除小鼠骨髓細(xì)胞總數(shù)沒有差異,同時(shí)骨髓細(xì)胞中 lineage陰性,Sca-1和c-kit雙陽(yáng)性的細(xì)胞也沒有差異。說(shuō)明年輕小鼠在正常生理狀態(tài)時(shí),RunX3基因?qū)υ煅杉?xì)胞穩(wěn)態(tài)維持及其分化沒有影響。
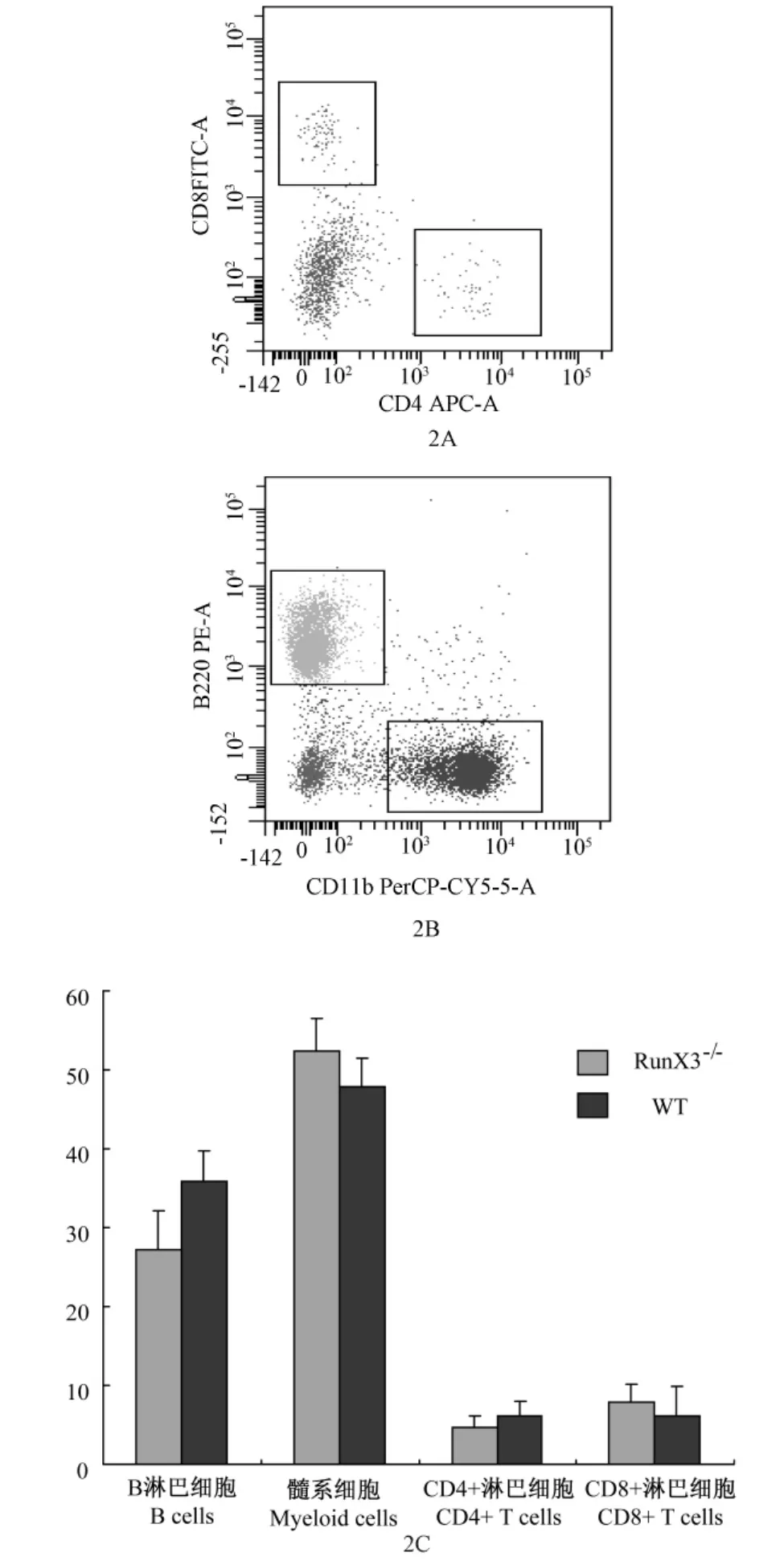
圖2 RunX3-/-小鼠骨髓中白細(xì)胞各亞群細(xì)胞比例分析Note:2C:the histogram showing the percentage of leukocyte subsets in the bone marrow of RunX3-/-mice.WT:wild-type mice;RunX3-/-:RunX3-/-mice.Fig.2 The percentage of leukocyte subsets in the bone marrow of RunX3-/--mice
2.2 競(jìng)爭(zhēng)性骨髓移植及5-FU處理
為進(jìn)一步研究 RunX3基因?qū)υ煅杉?xì)胞的造血重建能力和長(zhǎng)期自我更新能力是否有影響,我們進(jìn)行了競(jìng)爭(zhēng)性骨髓移植及5-FU處理實(shí)驗(yàn)。
2.2.1 競(jìng)爭(zhēng)性骨髓移植是評(píng)價(jià)造血干細(xì)胞自我更新能力的標(biāo)準(zhǔn)試驗(yàn):放射性照射后的受體中,供體來(lái)源的骨髓造血干細(xì)胞在造血重建的過(guò)程中,對(duì)外周血細(xì)胞的貢獻(xiàn)在移植后1月時(shí)主要來(lái)自于短期骨髓干細(xì)胞的造血及分化功能,在3月后則來(lái)自于長(zhǎng)期干細(xì)胞的功能表現(xiàn)。我們利用FACS分析受體小鼠外周血來(lái)源(圖3A、B、E、F)。統(tǒng)計(jì)結(jié)果顯示:移植1月后來(lái)源于 RunX3-/-小鼠骨髓干細(xì)胞供體的外周血細(xì)胞占總外周血細(xì)胞的比例與野生對(duì)照鼠相比無(wú)明顯差異(圖4A)。進(jìn)一步分析外周血細(xì)胞各個(gè)亞群中供體來(lái)源的血細(xì)胞的比例顯示,移植一月時(shí)來(lái)源于 RunX3-/-小鼠骨髓干細(xì)胞供體的外周血中髓系細(xì)胞占總外周血髓系細(xì)胞細(xì)胞的比例較野生對(duì)照鼠高(圖4B)。骨髓移植后3個(gè)月的流式分析結(jié)果與移植后 1個(gè)月時(shí)的分析結(jié)果趨勢(shì)一致(圖4C)。說(shuō)明RunX3基因缺失對(duì)骨髓短期和長(zhǎng)期造血干細(xì)胞的自我更新能力沒有影響,但是可能對(duì)骨髓造血干細(xì)胞向髓系血細(xì)胞的分化能力有促進(jìn)作用。
2.2.2 為了進(jìn)一步了解在應(yīng)激狀態(tài)和損傷修復(fù)情況下,RunX3基因缺失對(duì)骨髓短期和長(zhǎng)期造血干細(xì)胞的自我更新能力是否影響,我們對(duì)移植后3個(gè)月的小鼠進(jìn)行 5-FU處理。5-FU是 S期抑制劑,在DNA復(fù)制過(guò)程中可阻斷胸苷的合成[8]。使用5-FU處理后,可將增殖活躍的細(xì)胞殺死,使得處于休眠期的細(xì)胞進(jìn)入細(xì)胞周期。利用5-FU處理可以進(jìn)一步明確供體骨髓造血干細(xì)胞在損傷修復(fù)情況下的增殖及分化功能。在5-FU處理1周后外周血流式分析結(jié)果顯示:來(lái)源于 RunX3-/-小鼠骨髓干細(xì)胞供體的外周血細(xì)胞占總外周血細(xì)胞的比例與野生對(duì)照鼠相比無(wú)明顯差異;而來(lái)源于 RunX3-/-小鼠供體的外周血髓系細(xì)胞占總外周血髓系細(xì)胞的比例較野生對(duì)照鼠高(圖4D)。在5-FU處理1月后,來(lái)源于RunX3-/-小鼠的外周血細(xì)胞和髓系細(xì)胞分別占總外周血細(xì)胞和總髓系細(xì)胞的比例與野生對(duì)照鼠相比無(wú)明顯差異(圖 4E)。這些數(shù)據(jù)提示RunX3基因缺失不影響骨髓造血干細(xì)胞在應(yīng)激狀態(tài)下的損傷修復(fù)能力,但是可能影響骨髓造血干細(xì)胞的分化能力。
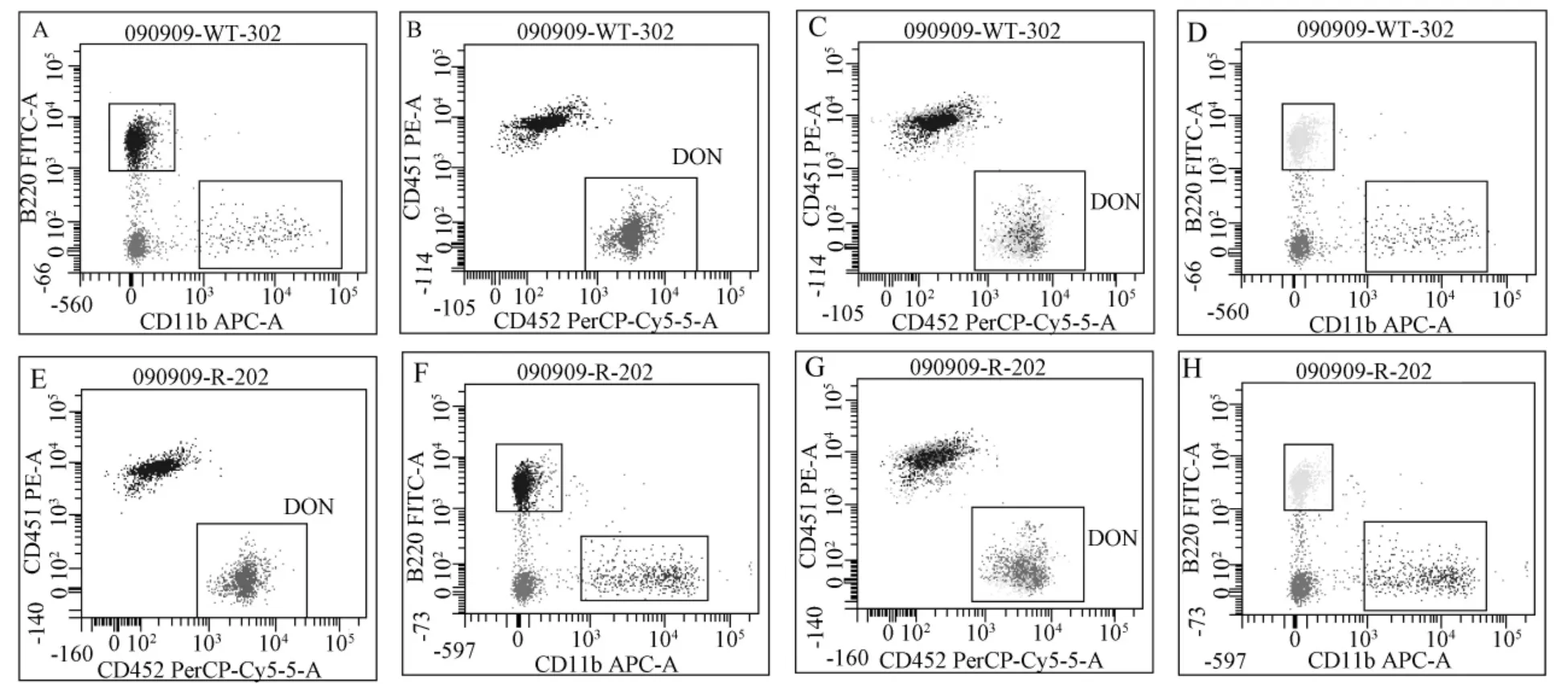
圖3 受體小鼠外周血中供體來(lái)源的細(xì)胞在骨髓移植后所占比例流式分析圖Note:A,B,C,D:FACS plots of the donor wild type mice.E,F(xiàn),G,H:FACS plots of the donor RunX3-/-mice.A,E:FACS plots of the proportion of leukocyte subsets in peripheral blood cells of recipient mice.B,F(xiàn):The proportion of leukocyte subsets in peripheral blood cells of recipient mice.C,G:Histogram showing the percentage of donor-derived peripheral blood cells in total peripheral cells after competitive transplantation.D,H:Histogram showing the percentage of donor-derived cells in subsets of peripheral cells.WT:wild-type mice;RunX3-/-:RunX3-/-mice.DON: donor-derived cells.Fig. 3 FACS plots show the percentage of donor-derived peripheral blood cells in recipient mice after competitive transplantation
2.2.3 為了進(jìn)一步明確RunX3基因缺失對(duì)骨髓造血干細(xì)胞分化能力的影響,我們分析了 RunX3-/-小鼠供體來(lái)源的外周血細(xì)胞中白細(xì)胞各亞群所占的比例(圖3C、D、G、H)。結(jié)果顯示:在移植后1個(gè)月和3個(gè)月時(shí),供體來(lái)源的B淋巴細(xì)胞和髓系細(xì)胞占總供體外周血細(xì)胞的比例在 RunX3-/-組和野生對(duì)照組之間差異無(wú)顯著性(圖5A、B)。在5-FU處理一周后外周血中髓系細(xì)胞比例 RunX3敲除組較野生組升高(圖5C),在5-FU處理一月后外周血中髓系細(xì)胞比例兩組無(wú)明顯差別,但B細(xì)胞比例下降(圖5D)。這組數(shù)據(jù)提示在應(yīng)激的早期RunX3基因的缺失可能導(dǎo)致了骨髓造血干細(xì)胞向髓系細(xì)胞的分化增多,而隨著時(shí)間的推移,骨髓造血干細(xì)胞向B淋巴細(xì)胞的分化也逐漸減少。
3 討論
已知RunX3基因?qū)τ诟杏X神經(jīng)中樞,免疫系統(tǒng),多種癌癥的發(fā)生均有重要作用,尤其是對(duì)于 T淋巴細(xì)胞的發(fā)育有重要調(diào)節(jié)作用[9]。然而 RunX3基因?qū)τ谠煅杉?xì)胞的影響還不清楚。我們對(duì)RunX3-/-小鼠骨髓干細(xì)胞的穩(wěn)態(tài)維持和造血重建能力進(jìn)行了研究。競(jìng)爭(zhēng)性骨髓移植實(shí)驗(yàn)是判斷干細(xì)胞自我更新能力的標(biāo)準(zhǔn)的實(shí)驗(yàn)。應(yīng)用致死劑量的放射線照射清除受體小鼠自體的骨髓干細(xì)胞后進(jìn)行移植供體骨髓,同時(shí)每只受體移植入等量的競(jìng)爭(zhēng)者骨髓細(xì)胞,受體小鼠外周血細(xì)胞主要來(lái)源于供體骨髓干細(xì)胞和競(jìng)爭(zhēng)性的骨髓干細(xì)胞的共同貢獻(xiàn)。由于競(jìng)爭(zhēng)性骨髓細(xì)胞作為內(nèi)參,同時(shí)每只受體小鼠所接受的受試供體骨髓細(xì)胞數(shù)量也是相等的,因此供體來(lái)源的外周血細(xì)胞所占的比例直接反映供體骨髓干細(xì)胞的自我更新能力。
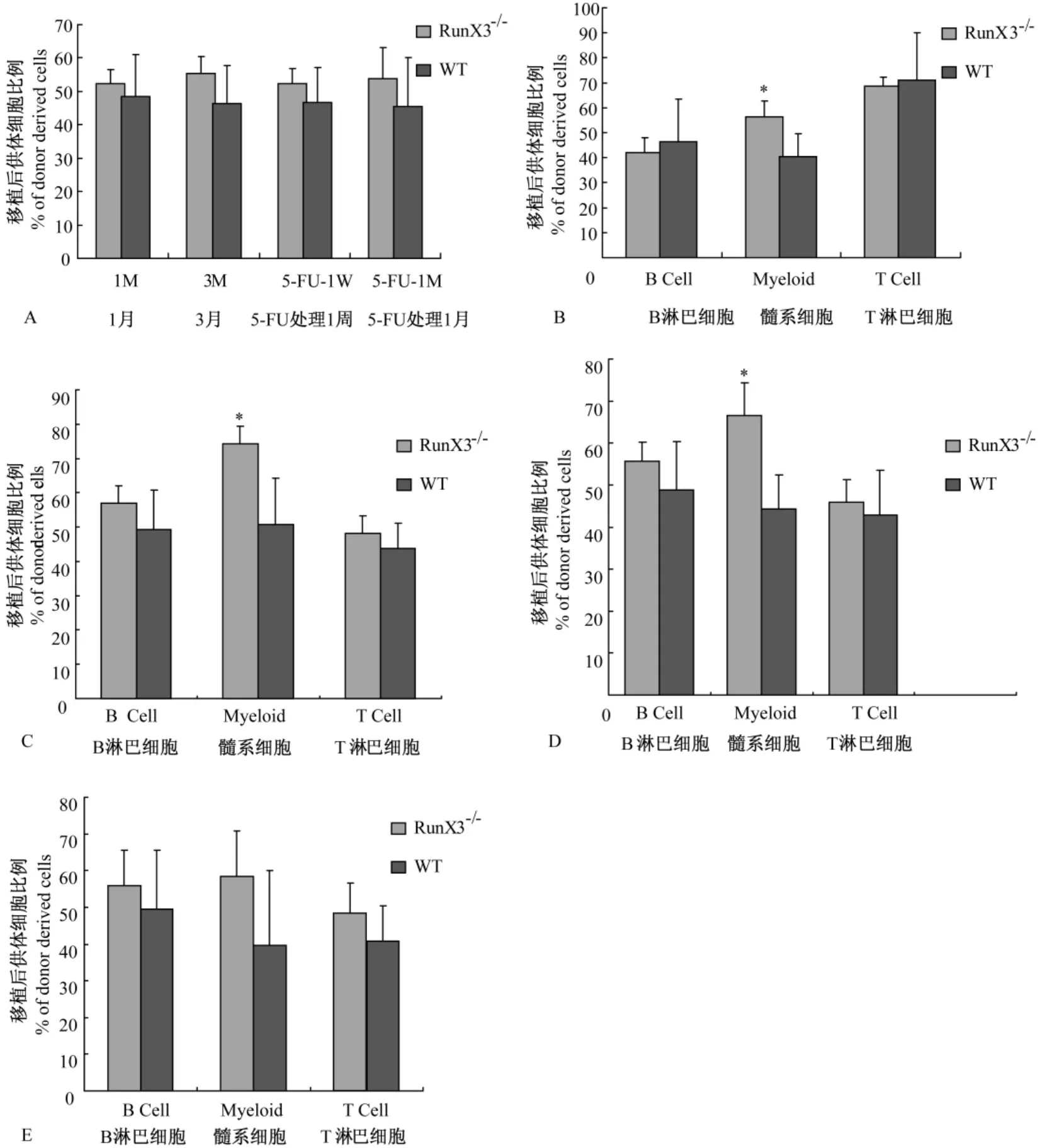
圖4 骨髓移植后受體小鼠外周血中供體來(lái)源的細(xì)胞所占比例Note:(A)Histogram showing the percentage of donor-derived peripheral blood cells in the total peripheral cells after competitive transplantation.(B) Histogram showing the percentage of donor-derived cells in the subsets of peripheral cells at one month after competitive transplantation.(C)Histogram showing the percentage of donor-derived cells in the subsets of peripheral cells at three months after competitive transplantation.(D)Histogram showing the percentage of donor-derived cells in the subsets of peripheral cells at one week after 5-FU treatment.(E)Histogram showing the percentage of donorderived cells in the subsets of peripheral cells at one month after 5-FU treatment.* P<0.05;WT:wild-type mice;RunX3-/-:RunX3-/-mice.Fig.4 The percentage of donor-derived peripheral blood cells in recipient mice after competitive transplantation
以往研究表明RunX3基因缺失的小鼠 CD11b陽(yáng)性的髓系細(xì)胞在過(guò)敏性氣道炎癥狀態(tài)下顯著增加[5]。我們研究發(fā)現(xiàn),RunX3基因?qū)δ贻p小鼠的骨髓造血系統(tǒng)向B淋巴細(xì)胞和髓系細(xì)胞分化的能力沒有影響,而且年輕小鼠在正常生理狀態(tài)時(shí),RunX3基因?qū)υ煅杉?xì)胞穩(wěn)態(tài)維持及其分化沒有影響。在移植后的應(yīng)激狀態(tài)下 RunX3-/-小鼠的骨髓造血干細(xì)胞在受體小鼠體內(nèi)的造血重建能力方面沒有顯著差異,但我們發(fā)現(xiàn)在應(yīng)激狀態(tài)下RunX3敲除小鼠的骨髓造血干細(xì)胞在受體小鼠體內(nèi)向髓系細(xì)胞分化能力增強(qiáng),而向 B淋巴細(xì)胞的分化能力減弱,提示RunX3基因可能參與了髓系細(xì)胞的分化過(guò)程,并在此過(guò)程中可能起抑制作用。
在正常生理狀態(tài)時(shí),RunX3基因?qū)π∈笤煅杉?xì)胞穩(wěn)態(tài)維持及其分化沒有影響。在移植后的應(yīng)激狀態(tài)下 RunX3-/-小鼠的骨髓造血干細(xì)胞在受體小鼠體內(nèi)的造血重建能力方面沒有顯著差異,但是在應(yīng)激狀態(tài)下向髓系細(xì)胞分化能力增強(qiáng),而向 B淋巴細(xì)胞的分化能力減弱,提示RunX3基因可能參與了骨髓造血干細(xì)胞的分化過(guò)程。有關(guān) RunX3基因在衰老過(guò)程中以及其他病理生理狀態(tài)下對(duì)造血干細(xì)胞的功能的影響還需要進(jìn)一步研究。
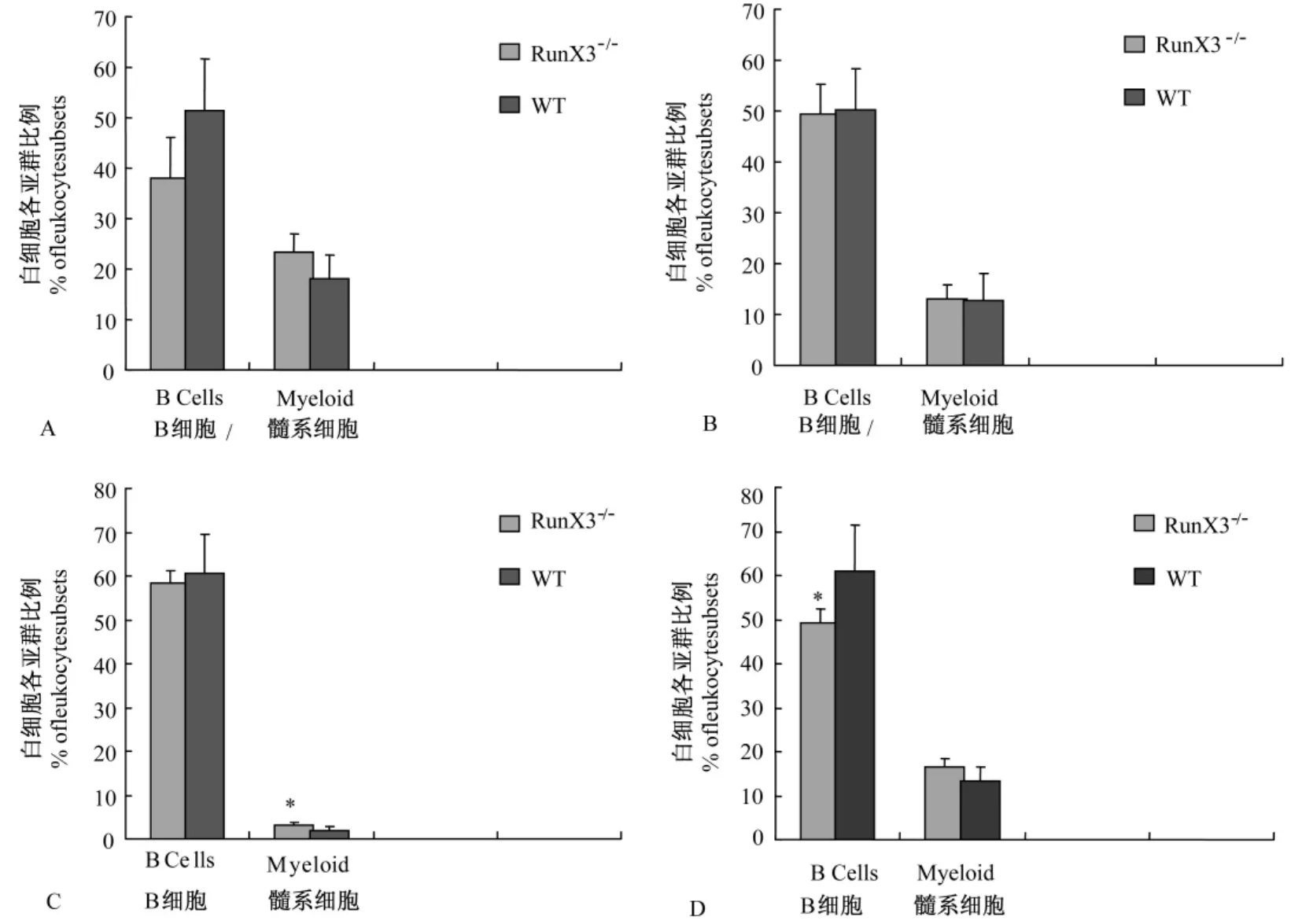
圖5 骨髓移植后供體來(lái)源的外周血細(xì)胞中白細(xì)胞各亞群所占的比例注:圖A顯示移植后1個(gè)月時(shí),供體來(lái)源的B淋巴細(xì)胞和髓系細(xì)胞占總供體外周血細(xì)胞的比例;圖B顯示移植后3個(gè)月時(shí),供體來(lái)源的B淋巴細(xì)胞和髓系細(xì)胞占總供體外周血細(xì)胞的比例;圖C顯示5-FU處理一周時(shí)外周血中供體來(lái)源的B淋巴細(xì)胞和髓系細(xì)胞占的比例;圖D顯示5-FU處理一月時(shí)外周血中供體來(lái)源的B淋巴細(xì)胞和髓系細(xì)胞占的比例。*:P<0.05;WT:野生型小鼠;RunX3-/-:RunX3-/-小鼠.Note:(A)Histogram showing the percentage of B lymphocytes or myeloid cells in the donor-derived blood cells at one month after competitive transplantation.(B)Histogram showing the percentage of B lymphocytes or myeloid cells in the donor-derived blood cells at three months after competitive transplantation.(C)Histogram showing the percentage of donor-derived B lymphocytes or myeloid cells blood cells in the donor-derived blood cells at one week after 5-FU treatment.(D)Histogram showing the percentage of donor-derived B lymphocytes or myeloid cells blood cells in the donorderived blood cells at one month after 5-FU treatment.* P<0.05;WT:wild-type mice;RunX3-/-:RunX3-/-mice.Fig.5 The percentage of leukocyte subsets repopulation in the donor-derived peripheral blood cells
[1] Inoue K,Shiga T,Ito Y.Runx transcription factors in neuronal development[J].Neural Dev,2008,3:20.
[2] Li QL,Ito K,Sakakura C,et al.Causal relationship between the loss of RUNX3 expression and gastric cancer[J].Cell,2002,109:113-124.
[3] Puig-Kroger A,Corbi A.RUNX3:a new player in myeloid gene expression and immune response[J].J Cell Biochem,2006,98:744-756.
[4] Woolf E,Xiao C,F(xiàn)ainaru O,et al.Runx3 and Runx1 are required for CD8 T cell development during thymopoiesis[J]. PNAS,2003,100:7731-7736.
[5] Fainaru O,Woolf E,Lotem J,et al.Runx3 regulates mouse TGF-beta-mediated dendritic cell function and its absence results in airway inflammation[J].EMBO J,2004,23:969-979.
[6] Alarcon-Riquelme ME.Role of RUNX in autoimmune diseases linking rheumatoid arthritis,psoriasis and lupus[J].Arth Res Ther,2004,6:169-173.
[7] Kevin D.Bunting.Hematopoietic Stem Cell Protocols[M]. Totowa,NJ:Humana Press.2008:87-88.
[8] Hoskins J,Butler JS.RNA-based 5-fluorouracil toxicity requires the pseudouridylation activity of Cbf5p[J].Genetics,2008,179:323-330.
[9] Wang L,Bosselut R.CD4-CD8 lineage differentiation:Thpoking into the nucleus[J].J Immunol,2009,183:2903-2910.
The Role of RunX3 Gene in Mouse Hematopoietic Stem Cells
YAO Chao,SHI Gui-ying,CHENG Zhi-yang,HUANG Xin,HU Yong-yan,JU Zhen-yu
(Key laboratory of Human Disease Comparative Medicine,Ministry of Health,Institute of Laboratory Animal Science,Chinese Academy of Medical Sciences and Peking Union Medical College,Beijing 100021,China)
ObjectiveTo study the role of RunX3 gene in the self-renewal and differentiation of hematopoietic stem cells(HSC)。MethodsThe number and function of hematopoietic stem cells in RunX3 knockout mice were determined by competitive bone marrow transplantation and flow cytometry。ResultsThe number of bone marrow stem cells in RunX3 knockout mouse was the same as those of wild type control mice.Competitive transplantation revealed no difference of HSC self-renewal in RunX3 knockout mouse,but a putative differentiation bias toward myeloid lineage。ConclusionRunX3 gene is not essential in the HSC self-renewal capacity,but appears to promote the HSC differentiation to myeloid lineage.
RunX3;knockout mice;Hematopoietic stem cells
R34915
A
1671-7856(2010)04-0028-06
2010-01-10
國(guó)家自然基金面上項(xiàng)目(30771189);十一五新藥專項(xiàng)支持(2009ZX09501-026)。
姚超(1978-),研究方向:干細(xì)胞、衰老與再生醫(yī)學(xué)。
鞠振宇,E-mail:zhenyuju@hotmail.com。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(bào)(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
