脫脂乳發酵過程中β-乳球蛋白和α-乳白蛋白抗原性變化規律*
2010-09-12 13:20:56張穎布冠好鄭喆羅永康
食品與發酵工業 2010年6期
張穎,布冠好,2,鄭喆,羅永康
1(中國農業大學食品科學與營養工程學院,北京,100083) 2(河南工業大學糧油食品學院,河南鄭州,450052)
脫脂乳發酵過程中β-乳球蛋白和α-乳白蛋白抗原性變化規律*
張穎1,布冠好1,2,鄭喆1,羅永康1
1(中國農業大學食品科學與營養工程學院,北京,100083) 2(河南工業大學糧油食品學院,河南鄭州,450052)
選擇 8株乳酸菌對脫脂乳進行發酵,通過間接競爭 EL ISA法測定了不同菌種、發酵時間、冷藏時間下發酵乳中β-乳球蛋白(β-LG)和α-乳白蛋白(α-LA)的殘留抗原性。結果表明,經不同乳酸菌發酵 12 h后,發酵乳中β-LG和α-LA的抗原性均出現不同程度的下降,且菌株間差異顯著。隨著發酵時間的延長,β-LG和α-LA的抗原性在 3~12 h內迅速下降,下降速度為瑞士乳桿菌 >保加利亞乳桿菌 >嗜熱鏈球菌,復合菌株發酵對降低抗原性具有協同效果;繼續發酵至 48h,抗原性變化緩慢。瑞士乳桿菌發酵乳在 4℃冷藏 7 d后,與冷藏前相比,α-LA的抗原性下降 5%左右,而β-LG的抗原性無顯著變化。
抗原性,發酵,乳酸菌,β-乳球蛋白,α-乳白蛋白
牛乳是較易引起過敏的食物之一,大約有 2%~6%的嬰幼兒對乳蛋白質有不同程度的過敏。乳蛋白中主要過敏原為乳清蛋白中的β-乳球蛋白(β-LG)和α-乳白蛋白(α-LA)。牛乳蛋白過敏可引起過敏性鼻炎、哮喘、濕疹、腹瀉、胃腸出血等疾病,甚至會導致過敏性休克,嚴重影響嬰幼兒的健康成長[1]。近年來研究發現,發酵乳制品具有改善消化、降低血中膽固醇、抗菌、抗癌等多種功能,日常膳食中食用發酵乳還能預防、減輕過敏癥狀[2-3]。Majamaa[4]研究表明,牛奶過敏嬰兒食用乳酸菌水解的乳清蛋白 1個月后,嬰兒的腸道炎癥可得到有效緩解。
關于采用乳酸菌發酵來降低乳清蛋白過敏原的研究國外已有一些報道。W róblewska[5]證明,牛乳經乳酸菌發酵后,β-LG和α-LA的抗原性降低 99%以上;Kleber[6]用乳酸菌發酵脫脂乳、甜乳清后,β-LG抗原性分別降低了 90%和 70%;Maier[7]研究指出,酸乳中β-LG的消化率比生牛乳提高 30%,不同發酵方式酸乳中蛋白的抗原性降低程度存在差異。而Britt[8]用瑞士乳桿菌對脫脂乳發酵 113 h后,β-LG含量雖逐漸減低,但 IgE介導的β-LG抗原性卻沒有顯著變化。可以看出,目前關于通過乳酸菌發酵來降低乳清蛋白抗原性的研究結果差異較大,同時對發酵過程中乳清蛋白抗原性的變化規律也缺乏系統的研究。
本實驗選擇 8株乳酸菌對脫脂乳進行發酵,研究了不同菌種、發酵時間、冷藏時間對發酵乳中β-LG和α-LA抗原性的影響,并通過測定發酵乳的 pH值和游離氨基酸含量,初步探討了導致發酵過程中乳清蛋白抗原性變化的原因,從而為低致敏牛乳制品的開發提供一定的實驗依據和理論基礎。
1 材料與方法
1.1 乳酸菌菌種
乳酸菌菌種共 8株:德氏乳桿菌保加利亞亞種(Lactobacillus delbrueckiisubsp.bulgaricus)S-5、S-7、S-7*,干酪乳桿菌(Lactobacillus casei),來自中國農業大學食品學院功能乳品實驗室。瑞士乳桿菌(Lactobacillus helveticus),干酪乳桿菌 Zhang(Lactobacillus casei),嗜酸乳桿菌(Lactobacillus acidophilus),嗜熱鏈球菌(Streptococcus ther m ophilus),來自吉林農科院和內蒙古農業大學。
1.2 試劑
β-乳球蛋白(編號 L3908)、α-乳白蛋白(編號L5385)、羊抗兔 IgG-HRP(1:10000,編號 A6154)、三硝基苯磺酸(編號 P2297),均為 Sigma公司;抗體為兔抗β-LG血清,兔抗α-LA血清,自制;L-亮氨酸,上海藍季科技發展有限公司;脫脂奶粉,新西蘭進口,其中水分3.8%,蛋白質 33.4%,脂肪 0.8%,灰分 7.9%。
1.3 儀器與設備
96孔酶標板(聚苯乙烯),Costar;Thermo Multiskan MK3酶標儀,Ther mo Electron公司;YT-CJ-1ND超凈工作臺,北京亞泰科隆實驗技術開發中心;YXQLS-18SI高壓蒸汽滅菌鍋,上海博訊實業有限公司;Merttle Toledo酸度計;UV-2600紫外可見分光光度計,尤尼柯(上海)儀器有限公司;W201電熱恒溫水浴鍋,上海申生科技有限公司;LRH-250生化培養箱,上海一恒科技有限公司。
1.4 實驗方法
1.4.1 乳酸菌的活化及傳代[9]
在無菌超凈工作臺中,挑取少量菌種凍干粉,轉移到 10%脫脂乳培養基(115℃,15 min滅菌)中,傳代培養 2~3次,使各菌株活力充分恢復。菌株活化好后,劃線培養于 MRS或 M17固態培養基的平板上,置于培養箱中培養至長出合適大小的菌落。挑單菌落鏡檢觀察所得菌株是否為純菌。若菌株已經為純菌,則進行下步實驗。
1.4.2 發酵乳樣品的制備
將脫脂乳粉按照 12.5%的質量比復原為脫脂乳,經 90℃,5 min熱處理后,冷卻至 45℃,在無菌超凈工作臺中進行乳酸菌接種。按 2%(v/v)的接種量接入單株或復合菌株發酵劑,搖勻后于 37℃(桿菌)或 42℃(球菌和復合菌)生化培養箱中進行發酵。分別于 0,3,6,9,12,18,24,36,48 h取樣,攪拌均勻后在 4℃下、8000 r/min離心 10 min,取上清液分裝于小管,-20℃凍藏待測。發酵乳凝乳后放 4℃冷藏,分別于 0,0.25,0.5,1,2,3,4,5,6,7 d取樣,以下步驟同上。
1.4.3 發酵乳 pH值的測定
用Merttle Toledo酸度計測定 pH值。
1.4.4 發酵乳中游離氨基酸含量的測定
采用三硝基苯磺酸(trinitrobenzene sulfonic acid,TNBS)法測定發酵乳中游離氨基酸含量[10],以衡量發酵乳中蛋白水解程度。
1.4.5 發酵產物抗原性測定
采用間接競爭 EL ISA方法測定發酵乳中β-LG和α-LA的殘余抗原性[11]。
間接競爭 EL ISA中被測物和包被抗原競爭與抗體結合,因此包被抗原與抗體生成復合物的量與被測物中抗原量成反比。被測物的抗原性大小可用抗原殘留量表示,按下式計算:
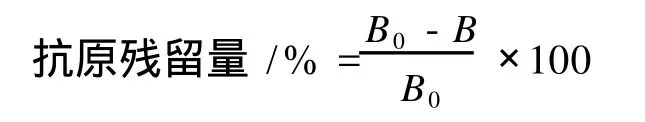
其中,B表示各被測樣品的 OD值,B0為無競爭體系的OD值。
1.4.6 數據分析
實驗數據利用 SAS9.0軟件(SAS Institute Inc.,Cary,NC,USA)進行單因素方差分析(ANOVA),用鄧肯氏多變域檢測法確定數據間的差異,P<0.05表示統計學上差異顯著。
2 結果與討論
2.1 乳酸菌菌種對發酵乳中乳清蛋白抗原性的影響
8株乳酸菌發酵脫脂乳 12 h后,發酵乳中β-LG和α-LA抗原性、pH值、游離氨基酸濃度見表 1。脫脂乳經 90℃,5 min處理后,β-LG和α-LA的抗原性有所升高,這主要是因為熱處理使蛋白質變性,較多隱藏的抗原表位暴露出來[12]。發酵 12 h后,脫脂乳中β-LG和α-LA的抗原性均出現不同程度的降低,不同菌株間差異顯著,以瑞士乳桿菌效果為最佳,β-LG和α-LA抗原性分別降低了 47.23%和 33%,而嗜熱鏈球菌效果最差,β-LG和α-LA抗原性僅分別降低了 16.93%和 16.77%。W róblewska[5]對滅菌乳(110℃,10 min)采用嗜溫和嗜熱乳酸菌或復合菌進行發酵,凝乳后乳清蛋白的抗原性比生乳降低 99%以上。Kleber[6]的研究表明,90℃,40 min熱處理的脫脂乳經不同乳酸菌發酵 24 h后,β-LG抗原性降低了 84%~98%。這或許說明發酵前熱處理的溫度對抗原性的變化有一定的影響,或者是菌株來源的不同導致上述差異[13-14]。表 1也表明,脫脂乳經 3株不同來源的保加利亞乳桿菌發酵后,β-LG和α-LA的抗原殘留量、pH值及游離氨基酸濃度有一定差別。
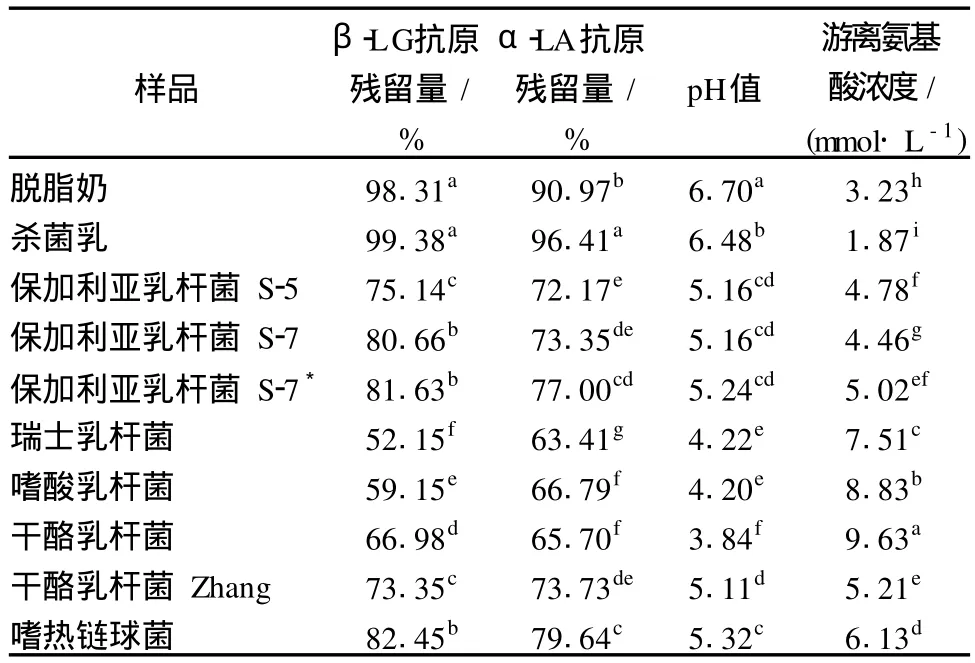
表1 脫脂乳經不同乳酸菌發酵 12 h后β-LG和α-LA抗原性、pH值、游離氨基酸濃度
另外,從表 1可知,瑞士乳桿菌、干酪乳桿菌、嗜酸乳桿菌發酵乳的 pH較低,游離氨基酸濃度較高,對應發酵乳中β-LG和α-LA的抗原性低;而嗜熱鏈球菌發酵乳的 pH較高,游離氨基酸濃度較低,其β-LG和α-LA的抗原性比較高。這或許說明發酵乳的酸度、游離氨基酸含量及蛋白抗原性 3者之間存在一定的關聯性。
2.2 發酵時間對發酵乳中乳清蛋白抗原性的影響
選取保加利亞乳桿菌、嗜熱鏈球菌、瑞士乳桿菌及桿菌與球菌以 1∶1比例復合菌株對脫脂乳進行發酵,于不同發酵時間取樣,測定發酵樣品中 pH值、游離氨基酸濃度及β-LG和α-LA的抗原性。
乳酸菌在乳中生長時將乳糖轉化為乳酸使酸度上升,pH值下降。從圖 1可知,發酵前 12 h,發酵乳的 pH值下降很快,如瑞士乳桿菌和嗜熱鏈球菌復合菌發酵 12 h后,pH由 6.55下降至 3.78;繼續發酵pH值下降變化緩慢。
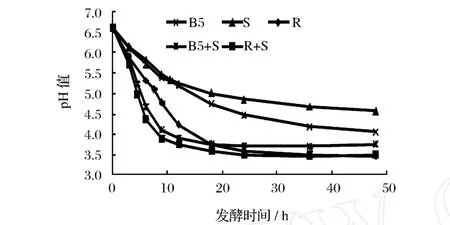
圖1 發酵時間對不同乳酸菌發酵乳中 pH的影響
發酵乳中游離氨基酸的濃度反映了在一定條件下菌體蛋白酶活力的強弱,即菌體水解乳蛋白能力的大小。從圖 2可知,發酵 0~12 h,發酵乳中游離氨基酸濃度大幅提高,特別是瑞士乳桿菌的蛋白酶水解活力較高,發酵 12h時游離氨基酸濃度達到 8.84 mmol/L,可能此階段乳酸菌數量增長快,活力高,產生了較多的蛋白酶來水解乳蛋白[13];繼續發酵,除保加利亞乳桿菌發酵乳中游離氨基酸含量仍逐漸增加外,其它菌株及復合菌變化不大。目前關于乳酸菌降低乳致敏性機理,被廣為接受的理論有 2個:一是乳酸菌可產生 I型及 II型干酪素,通過免疫調節機制防止過敏;另一種,可能是乳酸菌在發酵過程中產生了水解乳蛋白的蛋白酶,進而水解乳蛋白造成[15]。
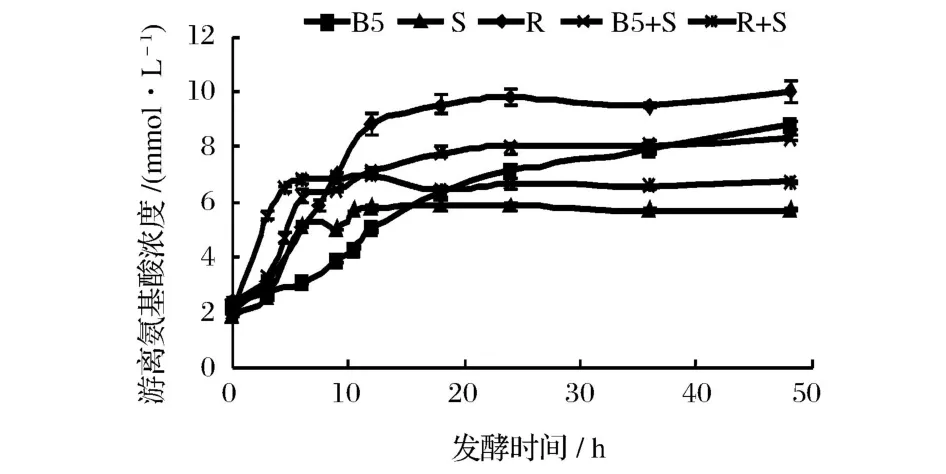
圖2 發酵時間對不同乳酸菌發酵乳中游離氨基酸濃度的影響
圖3和圖 4分別是不同乳酸菌發酵乳中β-LG和α-LA抗原性隨發酵時間的變化規律。可以看出,β-LG、α-LA抗原性變化和菌體發酵過程是密切相關的。發酵 3~12 h,脫脂乳中β-LG和α-LA的抗原性迅速下降,下降速度快慢依次為瑞士乳桿菌 >保加利亞乳桿菌 >嗜熱鏈球菌,保加利亞乳桿菌發酵脫脂乳12 h后,其β-LG抗原殘留量由 99.85%下降至71.68%,α-LA抗原殘留量由 96.35%下降至72.78%。此階段發酵乳中的游離氨基酸含量也在大幅提高,說明β-LG和α-LA抗原性的顯著下降與乳酸菌產生的蛋白酶水解其抗原表位有關。據報道,不同乳酸菌菌種產生的蛋白酶具有特異性,桿菌對乳蛋白的肽鏈內切酶活性要遠高于球菌,而嗜熱鏈球菌僅有有限的蛋白水解酶活性[16-18]。從圖 3和圖 4也可以看出,復合菌株發酵乳中乳清蛋白的抗原性下降速度更快,脫脂乳發酵6h后,瑞士乳桿菌中β-LG和α-LA抗原殘留量為 90.77%和 80.90%,嗜熱鏈球菌中β-LG和 α-LA抗原殘留量則為 99.68%和95.78%,而瑞士乳桿菌和嗜熱鏈球菌復合菌中β-LG和α-LA抗原殘留量僅為 59.64%和 66.95%,說明復合菌株發酵對降低乳清蛋白抗原性具有協同效果,這與 Kleber的研究結果相一致[6]。
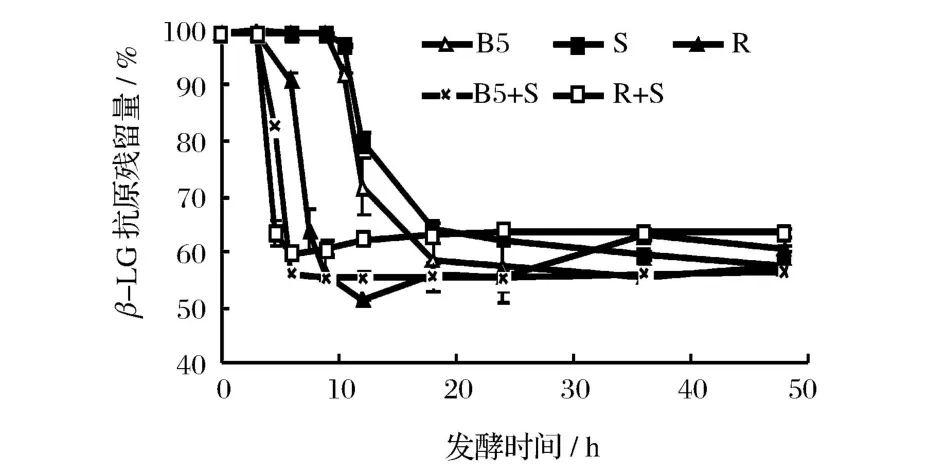
圖3 發酵時間對不同乳酸菌發酵乳中β-乳球蛋白抗原性的影響
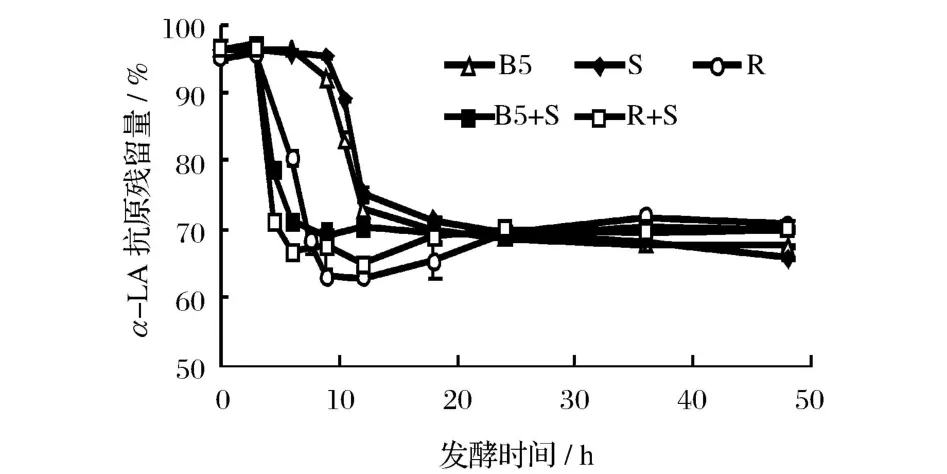
圖4 發酵時間對不同乳酸菌發酵乳中α-乳白蛋白抗原性的影響
繼續發酵至 48 h,發酵乳中乳清蛋白抗原性變化減緩,其中保加利亞乳桿菌和嗜熱鏈球菌發酵乳的抗原性繼續緩慢下降,可能是菌體產生的蛋白酶繼續酶解所致;而瑞士乳桿菌及其復合菌發酵乳抗原性則緩慢升高,則可能是因為乳清蛋白經蛋白酶水解為大肽后,菌株產生的肽酶又將大肽進一步水解,產生了更小的肽段和氨基酸,導致一些隱性表位或線性表位的暴露。這種差異與不同乳酸菌產生的蛋白酶具有特異性有一定關系[19-20]。
另外,圖 3和圖 4表明,各乳酸菌發酵對于β-LG和α-LA抗原性影響呈正相關,并且β-LG的抗原性較α-LA降低效果更好。可能是由于發酵前劇烈的熱處理使β-LG變性程度較大,空間結構充分展開,更易于蛋白酶的水解[7,17]。
2.3 冷藏時間對發酵乳中乳清蛋白抗原性的影響
脫脂乳經瑞士乳桿菌發酵凝乳后,放入 4℃冷藏,于不同冷藏時間取樣進行測定。圖 5顯示的是冷藏不同時間后瑞士乳桿菌發酵乳中β-LG和α-LA抗原性的變化情況。隨冷藏時間延長,發酵乳的α-LA的抗原性緩慢下降,與冷藏前相比,冷藏 7 d后α-LA抗原性下降 5%左右,但β-LG抗原性的變化不顯著。從圖 6可以看到,在冷藏期間,瑞士乳桿菌發酵乳的pH仍在緩慢下降,游離氨基酸濃度仍在緩慢升高,說明雖然冷藏期間乳酸菌生長和代謝變得緩慢,但乳酸菌產生的蛋白酶仍在水解蛋白質,只是可能由于溫度低、pH不適宜等因素造成蛋白水解酶活力下降,從而導致發酵乳中β-LG和α-LA抗原性變化幅度不大。
3 結論
脫脂乳經保加利亞乳桿菌、瑞士乳桿菌、干酪乳桿菌、嗜酸乳桿菌、嗜熱鏈球菌發酵 12 h后,其β-LG和α-LA的抗原性均出現不同程度的降低,不同菌株間差異顯著,以瑞士乳桿菌效果為最佳。

圖5 瑞士乳桿菌發酵乳 4℃冷藏下β-LG和α-LA抗原性隨冷藏時間的變化
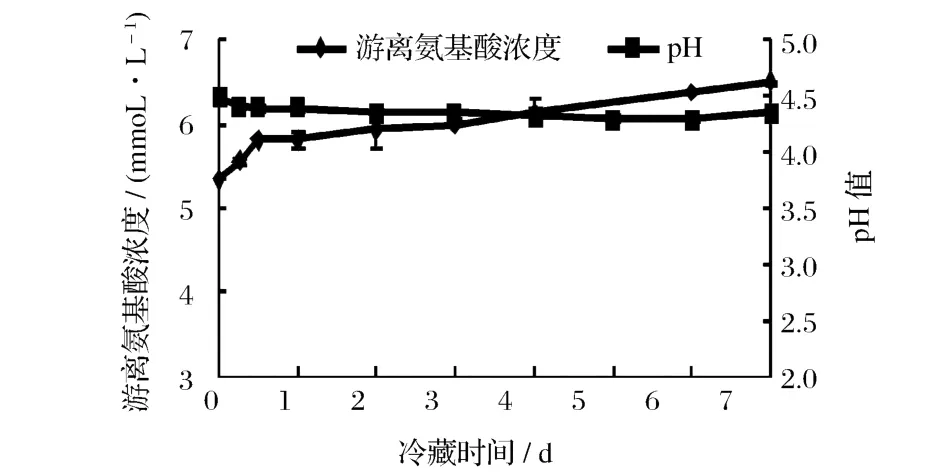
圖6 瑞士乳桿菌發酵乳 4℃冷藏下游離氨基酸濃度和 pH隨冷藏時間的變化
隨發酵時間延長,發酵乳中β-LG和α-LA的抗原性均先迅速下降后變化緩慢,抗原性下降速度依次為:瑞士乳桿菌 >保加利亞乳桿菌 >嗜熱鏈球菌,復合菌株發酵對降低β-LG和α-LA的抗原性具有協同作用;但繼續發酵至 48 h,發酵乳中蛋白抗原性變化緩慢,變化趨勢具有菌種特異性。瑞士乳桿菌發酵乳在 4℃冷藏 7d后,與冷藏前相比,α-LA的抗原性下降 5%左右,但β-LG的抗原性無顯著變化。
發酵乳中β-LG、α-LA抗原性變化和菌體發酵過程密切相關,乳酸菌產生的蛋白水解酶對降低乳清蛋白抗原性起著重要的作用。但發酵乳作為一個非常復雜的體系,其β-LG和α-LA抗原性的變化可能受多種因素的影響,發酵前熱處理溫度、發酵過程中生成的乳酸和牛乳體系中酪蛋白等對β-LG和α-LA抗原性的影響都有待進一步研究。
[1]Wal J M.Cow’s milk allergens[J].Allergy,1998,53:1013-1022.
[2]Salminen S,Ouwehand A C,Solauri E.Clinical applicationsof probiotic bacteria[J].InternationalDairyJournal,1998,8:563-72.
[3]Macfarlane G T,Cummings J H.Probiotics and prebiotics:can regulating the activities of intestinal bacteria benefit health?[J].British Medical Journal,1999,318:999-1003.
[4]Majamaa H,Isolauri E.Probiotics:A novel approach in themanagementof food allergy[J].JournalofAllergy and ClinicalImmunology,1997,99(2):179-185.
[5]Lucjan J,BarbraW.Reduction of the antigenicity ofwhey proteins by lactic acid bacteria fer mentation[J].Food and AgriculturalImmunology,1999,11:91-99.
[6]Kleber N,Weyrich U,Hinrichs J.Screening for lactic acid bacteria with potential to reduce antigenic response of β-lactoglobulin in bovine skim milk and sweet whey[J].InnovativeFood Science and Emerging Technologies,2006,7:233-238.
[7]Maier I,OkunV M,Pittne F,et al.Changes in peptic digestibility of bovineβ-lactoglobulin as a result of food processing studied by capillary electrophoresis andimmunochemicalmethods[J].Journal of ChromatographyB:Analytical Technologies in the Biomedical and Life Sciences,2006,41(8):160-167.
[8]EhnB M,Allmere T,Telemo E.Modification of IgEBinding toβ-Lactoglobulin by fer mentation and proteolysis of cow’smilk[J].Journal of Agricultural and Food Chemistry,2005,53(9):3743-3748.
[9]李平蘭,賀稚非主編.食品微生物學實驗原理與技術[M].北京:中國農業出版社,2005:238.
[10]Nissen J N.Deter mination of the degree of hydrolysis of food protein hydrolysis by trinitrobenzenesulfonic acid[J].Journal ofAgricultural and Food Chemistry,1979,27(6):1256-1263.
[11]沈小琴,鄭海,羅永康,等.酶解對乳清蛋白抗原性影響的研究[J].中國乳品工業,2006,34(6):12-15.
[12]KleberN,Hinrichs J.Antigenic response ofβ-lactoglobulin in thermally treated bovine skim milk and s weetwhey[J].Milchwissenschaft,2007,62:121-124.
[13]呂加平,駱承庠,劉鳳民.乳酸菌蛋白水解力的測定及研究[J].東北農業大學學報,1999,30(1):68-74.
[14]郭鴿,任大喜,張英華,等.發酵生產低致敏乳源蛋白基料的研究[J].食品與發酵工業,2007,33(10):81-84.
[15]CrossM L,Stevenson L M,Gill H S.Anti-allergy properties of fermented foods:an importantimmunoregulatory mechanis m of lactic acid bacteria?[J].InternationalImmunopharmacology,2001,1:891-90.
[16]Tzvetkova I,DalgalarrondoM,Danova S,et al.Hydrolysis ofmajor proteins by lactic acid bacteria from Bulgarian yogurts[J].Journal of Food Biochemistry,2007,31:680-702.
[17]Bertrand H C,Ivanova IV,DalgalarrondoM,et al.Evolution ofβ-lactoglobulin andα-lactalbumin content during yoghurt fer mentation[J].International Dairy Journal,2003,13:39-45.
[18]Rapp M.On the ability of lactic acid bacteria to degrade proteins[J].Milchwissenschaft,1969,24:208-211.
[19]Hromraksa P P,Nagano H,Boonmars T.Identification of proteolytic bacteria from Thai traditional fermented foods and their allergenic reducing potentials[J].Journal of Food Science,2008,4(73):189-195.
[20]Matar C,AmiotJ,SavoieL,et a1.The effectofmilk fermentation byLactobacillus helveticuson the release of peptides duringin vitrodigestion[J].Journal of Dairy Science,1996,79(6):971-979.
ABSTRACTIn this study,skim milk was fermented with eight lactic acid bacteria and the residual antigenicity of β-lactoglobulin(β-LG)andα-lactalbumin(α-LA)was determined bymeansof indirect competitive EL ISA at different fer mentation and storage time.The results showed that lactic acid bacteria fer mentation can reduce antigenicity of β-LG andα-LA significantly.Fermenting for 3-12h,antigenicity ofβ-LG andα-LA decreased rapidly and the descending speed was Lactobacillus helveticus>Lactobacillus bulgaricus>Streptococcus ther mophilus;Multi-bacteria fer mentation had collaborative effect.However,antigenicity changed slowly with further fer menting.Compared with pre-refrigeration,theα-LA antigenicity in Lactobacillus helveticus fer mented milk decreased by 5%after storage at 4℃ for 7d,while no significant variationswere found inβ-LG.
Key wordsantigenicity,fer mentation,lactic acid bacteria,β-lactoglobulin,α-lactalbumin
Research on the Antigen icity ofβ-LG andα-LA in the Process of Sk im Milk Fermentation
Zhang Ying1,Bu Guan-hao1,2,Zheng Zhe1,Luo Yong-kang1
1(College of Food Science and Nutritional Engineering,China AgriculturalUniversity,Beijing 100083,China)
2(College of Cereal and Food Science,Henan University of Technology,Zhengzhou 450052,China)
碩士研究生(羅永康教授為通訊作者)。
*國家自然科學基金項目(30871817,30471224)
2010-04-01
