肌肉生長(zhǎng)抑制素對(duì)脂肪細(xì)胞生長(zhǎng)發(fā)育和脂代謝的調(diào)控
2010-09-19 01:29:56姬云濤屈長(zhǎng)青
姬云濤,屈長(zhǎng)青,2
(1.阜陽(yáng)師范學(xué)院生命科學(xué)學(xué)院,阜陽(yáng) 236041;2.抗衰老中草藥安徽省工程技術(shù)研究中心,阜陽(yáng) 236041)
肌肉生長(zhǎng)抑制素對(duì)脂肪細(xì)胞生長(zhǎng)發(fā)育和脂代謝的調(diào)控
姬云濤1,屈長(zhǎng)青1,2
(1.阜陽(yáng)師范學(xué)院生命科學(xué)學(xué)院,阜陽(yáng) 236041;2.抗衰老中草藥安徽省工程技術(shù)研究中心,阜陽(yáng) 236041)
肌肉生長(zhǎng)抑制素(MSTN)對(duì)骨骼肌生長(zhǎng)的抑制作用已得到證實(shí),但其調(diào)控脂肪組織的作用還不是十分清楚。本文綜述了MSTN調(diào)控脂肪細(xì)胞生長(zhǎng)發(fā)育和脂代謝的相關(guān)機(jī)制和可能研究趨勢(shì)。
肌肉生長(zhǎng)抑制素;脂肪細(xì)胞發(fā)育;脂代謝;作用機(jī)制
肌肉生長(zhǎng)抑制素(myostatin,MSTN)為 TGF-β超家族成員,又稱為 GDF-8(growth/differentiation factor 8)。它是骨骼肌細(xì)胞分泌的高度保守的一種功能性糖蛋白[1],雖說(shuō)源于肌肉,但其靶器官并非只限于肌肉,在正常生理?xiàng)l件下,MSTN不但抑制骨骼肌細(xì)胞的生長(zhǎng)發(fā)育,同時(shí)可能還參與脂肪的生長(zhǎng)、分化和代謝調(diào)控。目前雖然對(duì)MSTN調(diào)控脂肪細(xì)胞生長(zhǎng)發(fā)育方面進(jìn)行了一些研究,但仍未得出十分明確的結(jié)果,有的研究表明它的突變或缺失會(huì)導(dǎo)致脂肪細(xì)胞的減少[2,3],它在骨骼肌中特異性過(guò)表達(dá),體內(nèi)的脂肪儲(chǔ)積會(huì)增加[4];但也有實(shí)驗(yàn)發(fā)現(xiàn)它的過(guò)表達(dá)會(huì)使脂肪幾乎全部喪失[5]。而研究脂肪細(xì)胞的增殖與分化以及脂代謝機(jī)理仍是當(dāng)今的一個(gè)熱點(diǎn),因此,MSTN對(duì)脂肪組織的確切作用還是值得深入研究的課題。本文就MSTN調(diào)控脂肪細(xì)胞生長(zhǎng)發(fā)育和脂代謝的研究作一綜述。
1 脂肪細(xì)胞的生長(zhǎng)發(fā)育特點(diǎn)
脂肪細(xì)胞的生長(zhǎng)分化增殖貫穿于整個(gè)生命過(guò)程,但是不同生長(zhǎng)發(fā)育階段其特點(diǎn)也不同。脂肪細(xì)胞發(fā)生的階段大致為:①中胚層發(fā)育而來(lái)脂肪組織源性干細(xì)胞(adipose-derived stem cells,ADSCs);②前體脂肪細(xì)胞發(fā)生克隆性增生;③細(xì)胞內(nèi)甘油三酯大量積累,形成典型的成熟脂肪細(xì)胞。白色脂肪組織(WAT)是體內(nèi)貯能的主要場(chǎng)所,主要由脂肪細(xì)胞組成,對(duì)維持體內(nèi)能量平衡發(fā)揮重要作用。脂肪細(xì)胞的生長(zhǎng)包括其體積的增大和新脂肪細(xì)胞的生成。新生動(dòng)物脂肪組織的增長(zhǎng)主要是新脂肪細(xì)胞的生長(zhǎng),即通過(guò)脂肪細(xì)胞分化,成年動(dòng)物以脂肪細(xì)胞的體積變化為主,但也保留有脂肪細(xì)胞的分化能力。研究認(rèn)為脂肪細(xì)胞的數(shù)量在出生前和生長(zhǎng)發(fā)育的早期就已被確定,而合成、蓄脂能力則受出生后各種因素的影響,在肥育期僅僅通過(guò)脂肪細(xì)胞體積的增大和重量增加來(lái)完成脂肪的沉積。
2 MSTN基因和蛋白
MSTN的合成受到多種結(jié)合于MSTN基因的啟動(dòng)子近端區(qū)域的細(xì)胞因子的轉(zhuǎn)錄調(diào)控。Moustakas等發(fā)現(xiàn),MSTN與其他 TGF-β超家族成員一樣,先合成前體蛋白質(zhì)(52×103),該前體蛋白由信號(hào)肽、編碼前肽(26×103)的 N-末端區(qū)和編碼成熟肽(12.5×103)的 C-末端區(qū)三部分組成[6]。前體蛋白質(zhì)經(jīng)兩次蛋白酶酶解活化,首先由信號(hào)肽引導(dǎo)將MSTN基因表達(dá)產(chǎn)物(大小約26×103的成熟糖蛋白)分泌到胞外,游離于血液中并處于潛伏狀態(tài),分泌后形成的前肽在RSRR區(qū)(蛋白酶水解位點(diǎn)Arg-Ser-Arg-Arg)被金屬蛋白酶(metalloproteinase)中的骨形態(tài)發(fā)生蛋白1(bone morphogenic protein-1/tolloid family)切除掉N端大約266個(gè)氨基酸,成熟區(qū)剩余的109個(gè)氨基酸通過(guò)其中的9個(gè)保守的半胱氨酸形成二硫鍵,生成二聚體后活化的MSTN與細(xì)胞膜上的Ⅰ型和Ⅱ型受體(激活素受體蛋白ⅠB,ActRⅠB和激活素受體蛋白ⅡB,ActRⅡB)相結(jié)合進(jìn)一步發(fā)揮作用(如圖1所示)。
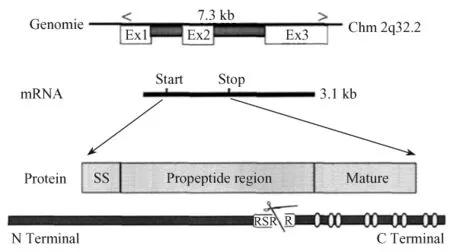
圖1 MSTN基因的和蛋白質(zhì)的結(jié)構(gòu)簡(jiǎn)圖Fig.1 A diagram of the structure of MSTN gene and protein
3 MSTN對(duì)脂肪組織的調(diào)控
目前對(duì)MSTN調(diào)控脂肪細(xì)胞生長(zhǎng)發(fā)育的作用機(jī)制的研究還很有限。叉頭轉(zhuǎn)錄因子(forkhead box class O,F(xiàn)oxO)是胰島素樣生長(zhǎng)因子(IGF)/磷脂酰肌醇3激酶(PI3K)/FoxO信號(hào)轉(zhuǎn)導(dǎo)通路中的關(guān)鍵分子。Nakae等[7]發(fā)現(xiàn),在胰島素受體缺失鼠,組成性的具活性的FoxO1阻礙前脂細(xì)胞的分化。而體外實(shí)驗(yàn)證明,MSTN通過(guò)抑制 IGF-1/PI3K/Akt通路,可以提高FoxO的活性[8]。還有實(shí)驗(yàn)表明FoxO1可以結(jié)合于MSTN啟動(dòng)子區(qū)域,并增強(qiáng)啟動(dòng)子活性,而誘導(dǎo)MSTN表達(dá)量增加[9]。C/EBPα是第一個(gè)被證明在脂肪細(xì)胞分化過(guò)程中起重要作用的轉(zhuǎn)錄因子。而PPARγ是PPARs家族中最具有脂肪專(zhuān)一性的亞型,是體內(nèi)脂肪形成的必需轉(zhuǎn)錄因子。Kim等[10]在用重組的 MSTN處理3T3-Ll脂肪細(xì)胞系時(shí)發(fā)現(xiàn),MSTN可能通過(guò) C/EBP-α和 PPAR-γ的信號(hào)傳導(dǎo)途徑抑制前體脂肪細(xì)胞的分化。Rebbapragada等[11]研究MSTN調(diào)控脂肪發(fā)生的信號(hào)通路時(shí),得到MSTN能夠抑制BMP-7誘導(dǎo)的脂肪細(xì)胞分化這一結(jié)果。其抑制過(guò)程與對(duì)肌細(xì)胞的作用方式一樣,在脂肪細(xì)胞中 MSTN同樣結(jié)合到絲氨酸/蘇氨酸受體ActRⅡB上,而后與 I型膜受體 ActRIB或 TbetaRI(TGF-β的I型受體)結(jié)合,并誘導(dǎo) Smad2/Smad3磷酸化并激活 TGF-β類(lèi)似的信號(hào)傳導(dǎo)通路,這樣MSTN通過(guò)阻止BMP-7結(jié)合到受體上而抑制BMP-7誘導(dǎo)脂肪細(xì)胞分化的活性。但值得注意的是,在實(shí)驗(yàn)中他們還發(fā)現(xiàn),MSTN能夠誘導(dǎo)C3H10T1/2間充質(zhì)干細(xì)胞系低水平表達(dá)脂肪細(xì)胞分化前期標(biāo)志基因C/EBP-α和 PPAR-γ,這是否能說(shuō)明 MSTN有促進(jìn)C3H10T1/2早期的成脂分化趨勢(shì)?因?yàn)樵谟^察3T3-L1前體脂肪細(xì)胞系時(shí),既沒(méi)有 C/EBP-α和PPAR-γ的低水平表達(dá),也沒(méi)有脂肪細(xì)胞分化后期標(biāo)志基因(GPDH和ADSF/resistin)表達(dá)。從他們的實(shí)驗(yàn)可以看出,雖然MSTN對(duì)3T3-L1和C3H10T(l/2)的成脂分化最終都表現(xiàn)出抑制,但在成脂分化的不同階段似乎存在不同。
有關(guān)MSTN對(duì)脂代謝調(diào)節(jié)的研究也不是很明確。Zhiliang等[12]在對(duì)雞 MSTN基因的5′調(diào)控區(qū)和3′調(diào)控區(qū)單核苷酸多態(tài)性(SNPs)的分析中也發(fā)現(xiàn),MSTN基因5′調(diào)控區(qū)的一個(gè)SNP位點(diǎn)與雞的脂肪的沉積有顯著相關(guān)。Lee等[13]發(fā)現(xiàn),MSTN基因敲除小鼠體形較野生型小鼠增大約30%,全身的所有骨骼肌都增大了2~3倍,脂肪重量沒(méi)有太大的變化。但進(jìn)一步實(shí)驗(yàn)[14]又發(fā)現(xiàn):MSTN基因敲除小鼠和肥胖小鼠雜交的雙突變小鼠,其脂肪減少,而且糖代謝紊亂也相應(yīng)得到改善。雜交后代小鼠在10個(gè)月之前體重超過(guò)肥胖小鼠,此后則與之體重相同甚至還會(huì)低于對(duì)照組。分析10個(gè)月之前體重超過(guò)肥胖小鼠可能是由于肌肉增大的原因,而10個(gè)月之后對(duì)照鼠體重增加應(yīng)該是脂肪含量增多所致。看來(lái)正如Lee等所提出,正常小鼠都會(huì)在年老時(shí)儲(chǔ)積過(guò)多的脂肪,而 MSTN-/-小鼠則根本不出現(xiàn)這種生理現(xiàn)象。是否與胚胎期MSTN缺失導(dǎo)致定向分化生成的前體脂肪細(xì)胞數(shù)少有關(guān)?是值得進(jìn)一步探討的。他們推測(cè)MSTN抑制脂肪沉積的機(jī)制一種可能是MSTN直接作用于脂肪組織來(lái)調(diào)節(jié)其代謝過(guò)程,另一種可能是骨骼肌中MSTN信號(hào)缺乏間接影響脂肪組織中的 MSTN突變,從而影響脂肪代謝。在Zimmers等[5]關(guān)于 MSTN與惡病質(zhì)關(guān)系的研究中,通過(guò)在無(wú)胸腺裸鼠的大腿肌肉植入穩(wěn)定表達(dá)小鼠MSTN的CHO細(xì)胞的方法,觀察全身給予過(guò)量的MSTN后的代謝變化。實(shí)驗(yàn)發(fā)現(xiàn),過(guò)量施與 MSTN的無(wú)胸腺裸鼠呈現(xiàn)一種明顯的惡病質(zhì)狀態(tài),除了全身骨骼肌重量減少之外,全身的白色脂肪也幾乎全部喪失。推測(cè)脂肪的下降可能與 Feldman等[15]的轉(zhuǎn)基因小鼠代謝率增高相似,因?yàn)?Feldman等的MSTN轉(zhuǎn)基因小鼠的脂肪細(xì)胞表現(xiàn)出對(duì)胰島素敏感度和葡萄糖氧化自發(fā)性增加,這些效應(yīng)能提高對(duì)飼料引起的肥胖的抗性,當(dāng)轉(zhuǎn)基因小鼠與野生型小鼠同時(shí)喂給高脂肪食物時(shí),轉(zhuǎn)基因小鼠抵御肥胖的能力明顯比野生型小鼠強(qiáng)。因此,可以考慮應(yīng)用MSTN治療一些代謝疾病,如由于飲食而引起的糖尿病,從而為解決日益增加的肥胖癥和治療代謝性疾病提供一種新的思路[16,17]。分析以上的研究結(jié)果可以看出:MSTN過(guò)表達(dá),導(dǎo)致肌萎縮和脂肪分解加強(qiáng);而MSTN缺失導(dǎo)致肌肉強(qiáng)大和脂肪減少,兩種情況下肌肉的變化是一致的,但脂肪減少途徑是不同的,前者是促進(jìn)脂肪分解(是針對(duì)機(jī)體已經(jīng)分化的成熟脂肪細(xì)胞),脂肪細(xì)胞數(shù)目沒(méi)變只是體積變小?后者是抑制脂肪細(xì)胞生長(zhǎng)發(fā)育(主要是不能促進(jìn)脂肪細(xì)胞早期的定向分化)有待確證。除了MSTN轉(zhuǎn)基因小鼠,F(xiàn)eldman等[15]同時(shí)也研究了MSTN代替地塞米松誘導(dǎo)多能性前體細(xì)胞(pluripotent progenitor cells)成脂分化,實(shí)驗(yàn)表明通過(guò)MSTN誘導(dǎo)的脂肪細(xì)胞在體外細(xì)胞培養(yǎng)與轉(zhuǎn)基因鼠體內(nèi)表現(xiàn)一樣,都很小并表達(dá)出未成熟的脂肪細(xì)胞的標(biāo)志特征。這是不是也暗示,在早期間充質(zhì)干細(xì)胞發(fā)育成前體脂肪細(xì)胞過(guò)程中,MSTN發(fā)揮促進(jìn)作用,而一旦前體脂肪細(xì)胞形成,MSTN則會(huì)抑制其向成熟脂肪細(xì)胞分化?而同樣是過(guò)表達(dá)MSTN,Reisz-Porszasz等[4]通過(guò)連接組織特異性啟動(dòng)子的方法,在雄性小鼠骨骼肌肉中特異性的過(guò)表達(dá)MSTN,同樣觀察到小鼠肌塊減少,但脂肪儲(chǔ)積卻增加。這與Zimmers等[5]的研究結(jié)果相悖,倒與下述 MSTN基因敲除小鼠的脂肪儲(chǔ)積減少一致。Lin等[3]通過(guò)對(duì)MSTN基因敲除小鼠(KO)與野生型小鼠(WT)研究比較發(fā)現(xiàn),KO鼠增加肌肉生成和減少脂肪生成,KO鼠脂肪組織中 C/EBP-α和 PPAR-γ以及l(fā)eptin水平也遠(yuǎn)遠(yuǎn)低于WT鼠。另外還有MSTN基因突變男孩[2]肌肉中 MSTN蛋白含量很低,出生時(shí)肌肉異常發(fā)達(dá),脂肪含量卻只有一半。這又一次表明MSTN功能的缺失影響脂肪的生成。
對(duì)MSTN調(diào)控脂肪細(xì)胞增殖與分化的研究,目前常用的細(xì)胞系有以C3H10T(1/2)為代表的多能干細(xì)胞系以及以3T3-L1為代表的前體脂肪細(xì)胞系[18,19]。Artaza 等[20]利用重組 MSTN 和抗 MSTN抗體,對(duì)C3H10T(l/2)間充質(zhì)干細(xì)胞系成脂成肌分化中的作用研究表明,MSTN抑制 C3H10T(l/2)的成肌分化,促進(jìn)C3H10T(l/2)向脂肪分化。根據(jù)該實(shí)驗(yàn)中MSTN作用的時(shí)間分析,其抑肌促脂作用是在干細(xì)胞系前期分化階段,那么在后期成肌細(xì)胞和前體脂肪細(xì)胞一旦形成,繼續(xù)成熟分化時(shí),MSTN的這種抑肌促脂作用是否一致呢?從前面Rebbapragada等[11]的研究推測(cè),MSTN 可能誘導(dǎo)C3H10T(l/2)干細(xì)胞系前期向前體脂肪細(xì)胞分化,但后期卻抑制其向成熟脂肪細(xì)胞分化。Kim等[21]的研究結(jié)果就表明,MSTN抑制3T3-L1前體脂肪細(xì)胞系向成熟脂肪細(xì)胞分化。這一結(jié)果同樣在牛前體脂肪細(xì)胞分化中得到證實(shí)[22]。盡管對(duì)于脂肪細(xì)胞的終末分化過(guò)程已有了深入的認(rèn)識(shí),如:發(fā)現(xiàn)了前體脂肪細(xì)胞的克隆擴(kuò)增(mitotic clonal expansion)、過(guò)氧化物酶體增殖物激活受體 γ(peroxisome proliferator-activated receptor-γ,PPAR-γ) 的關(guān)鍵作用等,但由于受到實(shí)驗(yàn)材料的限制,所以大部分成果僅限于前體脂肪細(xì)胞的增殖及其分化為成熟脂肪細(xì)胞這一階段,對(duì)脂肪細(xì)胞分化早期決定時(shí)的分子事件極少涉及,而一般文獻(xiàn)上所提到的脂肪細(xì)胞分化通常是指前脂肪細(xì)胞分化為成熟脂肪細(xì)胞的過(guò)程。可以看出,對(duì)于MSTN在C3H10T(l/2)干細(xì)胞系成脂分化的整個(gè)過(guò)程中的作用如何還沒(méi)有完整的答案。
目前,對(duì)MSTN如何控制脂肪生長(zhǎng)發(fā)育及代謝的機(jī)理知之甚少,而且粗略看來(lái)還有矛盾之處。例如同是MSTN基因敲除,大多表現(xiàn)為體內(nèi)脂肪減少,但也有報(bào)道脂肪無(wú)變化;同是MSTN過(guò)表達(dá),有的脂肪明顯減少,有的則表現(xiàn)為脂肪增多。可能MSTN對(duì)脂肪組織的作用主要是通過(guò)影響脂肪發(fā)生和脂代謝這兩方面來(lái)實(shí)現(xiàn)的,而且MSTN對(duì)脂肪發(fā)生的作用可能因靶細(xì)胞類(lèi)型、不同的發(fā)育生長(zhǎng)階段、外界微環(huán)境等的影響而呈現(xiàn)完全不同的效應(yīng),并且MSTN對(duì)機(jī)體脂肪調(diào)節(jié)可能存在著不同調(diào)控通路。根據(jù)已發(fā)表的研究結(jié)果推測(cè),MSTN可能作為細(xì)胞因子分泌到血液中,通過(guò)血液循環(huán)到達(dá)脂肪細(xì)胞,而后直接作用于脂肪細(xì)胞從而發(fā)揮影響脂肪代謝的作用。那么在胚胎發(fā)育早期,沒(méi)有MSTN的作用會(huì)不會(huì)影響間充質(zhì)干細(xì)胞定向分化出脂肪源性干細(xì)胞或前體脂肪細(xì)胞的數(shù)量?敲除MSTN基因或MSTN基因突變的個(gè)體脂肪沉積減少,是在出生后表現(xiàn)出來(lái)的,這與出生前分化出的脂肪源性干細(xì)胞或前體脂肪細(xì)胞的數(shù)量減少也可能有關(guān)?本實(shí)驗(yàn)室目前研究的結(jié)果也初步顯示,MSTN調(diào)控間充質(zhì)干細(xì)胞成脂分化過(guò)程中有階段性變化(數(shù)據(jù)整理中)。
通過(guò)對(duì)以上各項(xiàng)研究結(jié)果的分析,是否可以推測(cè):MSTN在胚胎期促進(jìn)脂肪源性干細(xì)胞和前體脂肪細(xì)胞的生成;而后又抑制前體脂肪細(xì)胞向成熟脂肪細(xì)胞分化;對(duì)脂肪組織則表現(xiàn)為促進(jìn)脂分解代謝。當(dāng)然這些推測(cè)中間還有很多問(wèn)題尚待進(jìn)一步探索,還有待于進(jìn)一步通過(guò)系統(tǒng)的全面的研究得以檢驗(yàn)和確證。畢竟在脂肪細(xì)胞分化過(guò)程中,存在著復(fù)雜調(diào)控機(jī)制,除了要考慮多種調(diào)節(jié)因素的相互作用外,還要考慮體內(nèi)外環(huán)境的差異。隨著對(duì)脂肪細(xì)胞分化調(diào)控研究的深入,進(jìn)一步研究MSTN對(duì)脂肪細(xì)胞發(fā)育和脂代謝的調(diào)控,對(duì)控制動(dòng)物脂肪、提高動(dòng)物胴體瘦肉率和肉品質(zhì)非常重要,而且對(duì)人類(lèi)肥胖病、與血糖血脂有關(guān)的代謝病的研究也將起著很大的推動(dòng)作用。
[1] Amirouche A,Durieux AC,Banzet S,et al.Down-regulation of Akt/mammalian targetofrapamycin signaling pathway in response to myostatin overexpression in skeletal muscle[J].Endocrinology,2009,150(1):286-294.
[2] Schuelke M,Wagner KR.Stolz LE,et al.Myostatin mutation associated with gross muscle hypertrophy in a child[J].New Engl J Med,2004,350(26):2682-2688.
[3] Lin J,Arnold HB,Della-Fera MA,et al.Myostatin knockout in mice increases myogenesis and decreases adipogenesis[J].Biochem Biophys Res Commun,2002,291:701-706.
[4] Reisz-Porszasz S,Bhasin S,Artaza JN,et al.Lower skeletal muscle massin male transgenic mice with muscle-speciflc overexpression of myostatin[J].Am J Physiol Endocrinol Metab,2003,285:E876-888.
[5] Zimmers TA,Davies MV,Koniaris LG,et al.Induction of cachexia in mice by systemically administered myostatin[J].Scienee,2002,296:1486-1488.
[6] Grobet L,Martin LJ,Poncelet D,et al.A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle[J].Nat Genet,1997,17(1):71-74.
[7] Nakae J,Cao Y,Oki M,et al.Forkhead transcription factor FoxO1 in adipose tissue regulates energy storage and expenditure[J].Diabetes,2008,57(3):563-576.
[8] Stitt TN,Drujan D,Clarke BA,et al.The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors[J].Mol Cell,2004,14:395-403.
[9] Foulstone E,Prince S,Zaccheo O,et al.Insulin-like growth factor ligands,receptors,and binding proteins in cancer[J].J Pathol,2005,205(2):145-153.
[10] Kim HS,Liang L,Dean RG,et al.Inhibition of preadipocyte differentiation by myostatin treatment in 3T3-L1 cultures[J].Biochem Biophys Res Commun,2001,281:902-906.
[11] Rebbapragada A,Benchabane H,Wrana JL,et al.Myostatin signals through a transforming growth factor beta-like signaling pathway to block adipogenesis[J].Mol Cell Biol,2003,23:7230-7242.
[12] Zhiliang G,Dahai Z,Ning L,et al.The single nucleotide polymorphisms of the chicken myostatin gene are associated with skeletal musele and adipose growth[J].Sci China C Life Sci,2004,47:25-30.
[13] McPherron,AC,Lawler,AM,Lee,SJ.Regulation of skeletal muscle in mice by a new TGF-β superfamily member[J].Nature,1997,387(6628):83-90.
[14] Alexandra C,McPherron AC,Lee SJ.Suppression of body fat accumulation in myostatin-deficient mice[J].J Clin Invest,2002,109:595-601.
[15] Feldman BJ, StreeperRS, Farese RV, etal. Myostatin modulates adipogenesis to generate adipocytes with favorable metabolic effects[J].Proc Natl Acad Sci U S A,2006,103(42):15675-15680.
[16] 董世芬,洪纓,樊江波,等.實(shí)驗(yàn)性2型糖尿病心肌病大鼠模型的建立與評(píng)價(jià).中國(guó)實(shí)驗(yàn)動(dòng)物學(xué)報(bào),2009,17(4):245-251.
[17] 崔麗娟,都健,王娟,等.高脂喂養(yǎng)大鼠肝臟的NF-κBp65表達(dá)與胰島素抵抗的相關(guān)性.中國(guó)實(shí)驗(yàn)動(dòng)物學(xué)報(bào),2008,17(6):417-420.
[18] Rosen ED,Walkey CJ,Puigserver P,et al.Transcriptional regulation of adipogenesis[J].Genes Dev,2000,14(11):1293-1307.
[19] Novakofski J.Adipogenesis:usefulness of in vitro and in vivo experimental models[J].J Anim Sci,2004,82(3):905- 915.
[20] Artaza JN,Bhasin S,Magee TR,et al.Myostatin inhibits myogenesis and promotes adipogenesis in C3H10T(l/2)mesenchymal multipotent cells[J].Endocrinology 2005,146:3547-3557.
[21] Kim HS,Liang L,Dean RG,et al.Inhibition of preadipocyte differentiation by myostatin treatment in 3T3-L1 cultures[J].Biochem Biophys Res Commun,2001,281:902-906.
[22] Hirai S,Matsumoto H,Hino N.Myostatin inhibits differentiation of bovine preadipocyte[J].Domest Anim Endocrinol,2007,32:1-14.
Research Progress on MSTN Regulation Mechanism of Adipocyte Development and Lipid Metabolism
JI Yun-tao1QU Chang-qing1,2
(1.School of Life Science,F(xiàn)uyang Normal College.Fuyang 236041,China;2.Engineering Technology Research Center of Anti-Aging Chinese Herbal Medicine,Anhui Province,F(xiàn)uyang 236041,China)
Myostatin(MSTN),a negative regulator of skeletal muscle mass,is postulated to be involved in the pathogenesis of muscle fibrosis.But its role in the regulation of adipose tissue development is not fully understood yet.In this paper,the related mechanisms of MSTN in regulation of adipocyte growth and differentiation and lipid metabolism are reviewed and the recent progress in the research in this field is discussed.
Myostatin,MSTN;adipocyte,development;Lipid metabolism;Mechanism
Q952,R332
A
1005-4847(2010)05-0437-04
10.3969/j.issn.1005-4847.2010.05.18
2010-03-02
安徽省高校自然科學(xué)研究資助項(xiàng)目(No.KJ2008B228)。
姬云濤,女,博士,副教授,主要研究方向?yàn)閯?dòng)物繁育與生物技術(shù)。
屈長(zhǎng)青,男,博士,教授,主要研究方向細(xì)胞衰老與信號(hào)轉(zhuǎn)導(dǎo)。E-mail:qucq518@163.com.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級(jí))(2025年8期)2025-08-18 00:00:00
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國(guó)語(yǔ)大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級(jí))(2017年10期)2017-10-10 01:00:05
汽車(chē)工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02
云南中醫(yī)學(xué)院學(xué)報(bào)(2014年3期)2014-07-31 18:57:34
