雙組分調節系統對乳酸菌產細菌素過程的影響
2011-01-08 05:39:04楊郁葒王輝
中國乳品工業 2011年6期
楊郁葒,王輝
(1.黑龍江省完達山乳業股份有限公司,哈爾濱 150078;2.東北農業大學乳品科學教育部重點實驗室/食品學院,哈爾濱 150030)
雙組分調節系統對乳酸菌產細菌素過程的影響
楊郁葒1,王輝2
(1.黑龍江省完達山乳業股份有限公司,哈爾濱 150078;2.東北農業大學乳品科學教育部重點實驗室/食品學院,哈爾濱 150030)
綜述了雙組分調節系統的組成,以及雙組分調節系統對兩類細菌素合成的調節作用,來初步了解雙組分調節系統在乳酸菌產細菌素過程中的作用機制。
雙組分;細菌素;蛋白激酶;反應調節器
0 引言
雙組分系統最初是由Ninfa和Magasnik在研究大腸桿菌氮調節蛋白系(Nitrogen Regulatory Protein,NR)時發現的[1],該系統調節著大腸桿菌的基因表達。
近年來,越來越多有關應用一種稱為基于外信息素肽信號轉換途徑來調節多種細胞活動的革蘭氏陽性菌得到報道[2]。這些調節細胞活動包括:細菌感受性的生長(H?varstein and Morrison,1999)、穩定期時葡萄球菌素的分泌[3],還包括已知的產生于多種乳酸菌的細菌素。在乳酸菌所產的細菌素中,其中第二類細菌素的轉錄是通過由3種成分組成的信號轉換途徑完成的。這3種成分為:誘導類細菌素肽、HPK和RR。事實上,許多抗微生物小肽由細胞密度響應機制所調節。
1 雙組份調節系統的組成
典型的雙組份調節系統包括HPK和RR兩個組分。組氨酸蛋白激酶(HPK)感受器通常位于細胞質膜上以監測環境變化,反應調節器(RR)位于細胞質中并傳遞來自感受器的信號和調節基因的表達,可以是對一個調節子中的一個或多個基因的表達進行調節[4],以響應外界的變化。整個雙組分信號通路由信號輸入(input)、HPK自身磷酸化、RR磷酸化及輸出(output)等環節構成。
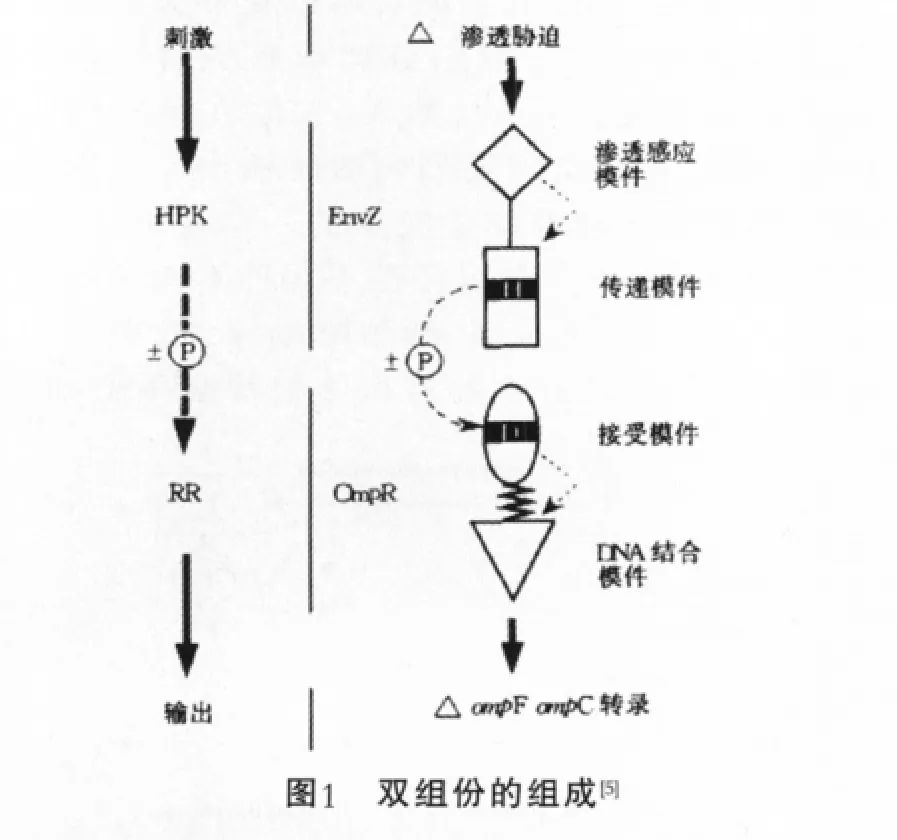
HPK通常是由一個信號輸入模件(input module)和一個自身激酶模件(catalytic module)組成,其中自身激酶模件可分為兩個子域:組氨酸磷酸轉移酶子域和ATP結合子域。HPK的外界信號輸入模件,與蛋白激酶的催化模件相連接,這一結構特點使得HPK能很靈敏地感受外界環境的變化[6]。序列分析表明,這類蛋白質C末端區域是高度保守的,而且由大約240個氨基酸連接數量不變的殘基組成。組氨酸殘基蛋白質自磷酸化作用消耗ATP,稱為H結合位點。G1、G2結合位點是由大量的甘氨酸組成,其序列類似與其他蛋白質的核苷酸結合基序。F、N結合位點通過序列分析,其功能還不是很清楚,大概與蛋白質的催化活性有關(圖2)。組氨酸蛋白激酶中的激酶,如EnvZ和NtrB起磷蛋白磷酸酶的作用,OmpR和NtrC在反應調節器中加速去磷酸化的作用。組氨酸蛋白激酶可分為兩類:跨膜蛋白和細胞質蛋白,跨膜蛋白能感應環境中的信號,而細胞質蛋白則感應細胞內的信號。許多革蘭氏陽性菌種中發現的組氨酸蛋白激酶都為跨膜蛋白。
RR通常是由調節區域,即感受器,和DNA結合區域,即輸出器組成。當激酶的輸入模件檢測到外部的信號時使自身激活,激酶的這個功能是通過組氨酸殘基經由ATP水解供能發生磷酸化產生的[7]。隨后磷酸基團傳遞到應答調節蛋白的保守天冬氨酸位點上,使其發生磷酸化。磷酸化的應答調節蛋白,能與DNA或其它信號蛋白發生相互作用,進而調節下游的信號轉導。在細菌中,大多數應答調節蛋白是轉錄因子,能夠調節相關基因表達。
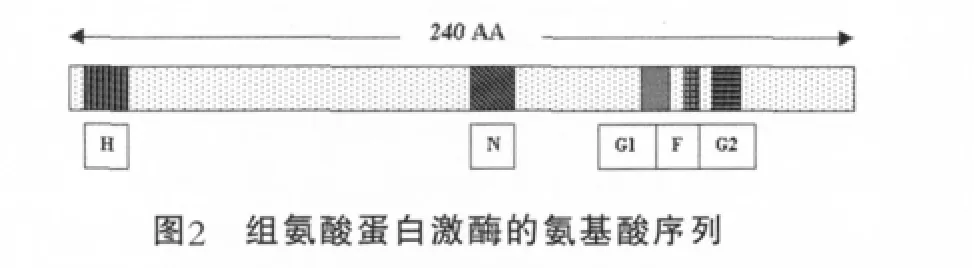
圖2中,H為組氨酸結合位點;N為天冬氨酸結合位點;G1和G2是不同甘氨酸的結合位點;F為苯丙氨酸的結合位點。
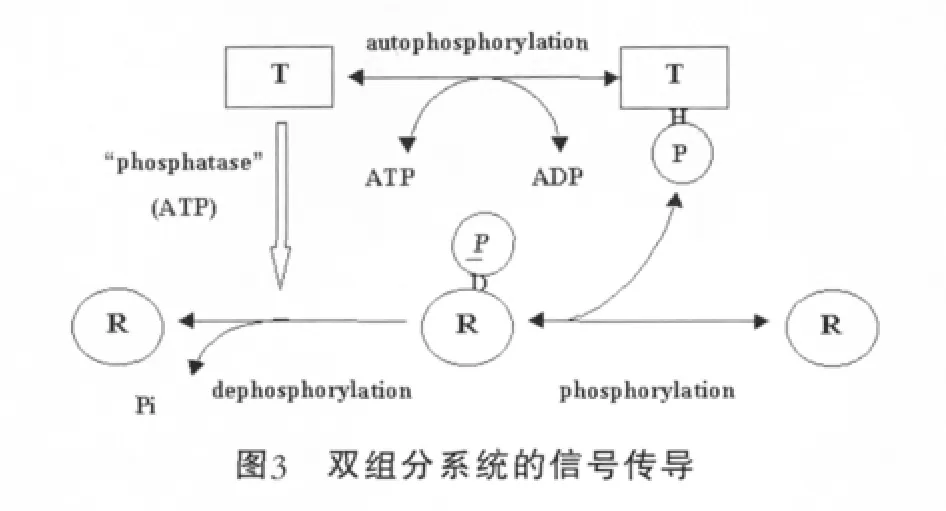
HPK和同源的RR是通過磷酸化和去磷酸化反應來進行溝通的(圖3),雙組分調節系統信號轉導的潛在分子學機理是簡單的,然而,它們在是怎樣收集環境信號、確信在感受到強烈刺激時輸出反應器能夠順利進行等方面的機理卻是復雜的。迄今為止,發現了大量的HPK和RR,它們存在于50種不同的細菌和真核生物中,包括鏈孢霉屬、釀酒酵母等。這就可以推論出這種雙組分系統在生物有機體中普遍存在。
2 羊毛硫抗生素合成的調節
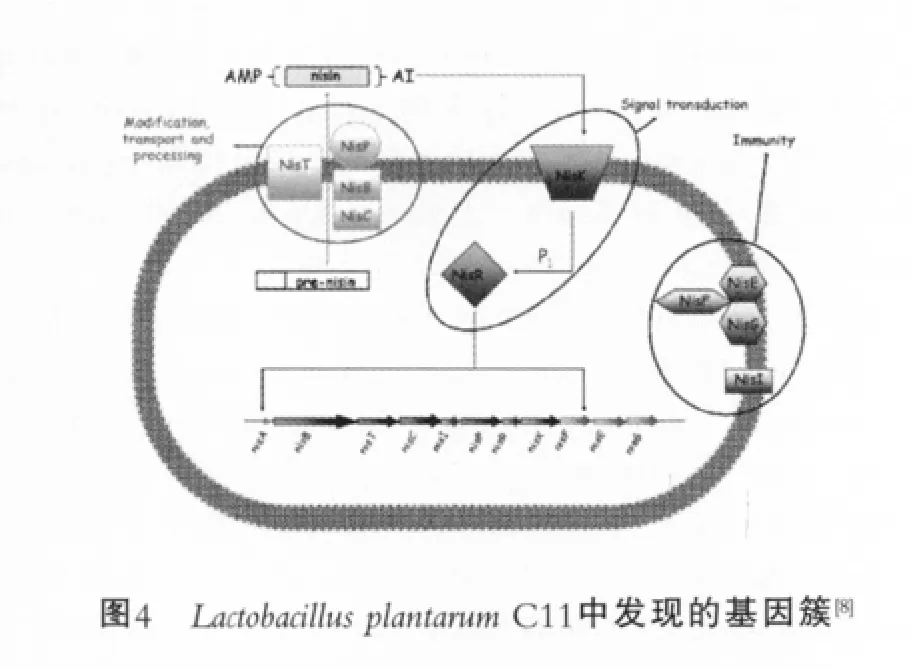
羊毛硫抗生素是小的熱穩定肽,結構中起作用的物質是硫醚環修飾的氨基酸,這類細菌素中最有名的是nisin。羊毛硫抗生素的生物合成、分泌和免疫所需的基因是在群體中構成的,即形成基因簇。nisin的產生開始于對數生長期,在到達細菌密度最高的穩定期時達到最高水平[8]。
基因簇nisABTCIPRKFEG編碼nisin的生物合成[9],除了結構基因、加工處理基因和免疫基因外,基因簇中的nisR(RR)和niskK(HPK)組成了與nisin生物合成有關的雙組分調節系統。高產nisin的Lc.Lactis菌株,其nisA基因缺少4個堿基對,稱為ΔnisA基因,它不但降低nisin的產生量,而且終止nisA的轉錄。在加入nisin后,ΔnisA恢復轉錄功能[10],因此,nisin的作用不僅是作為細菌素,而且作為一種分泌信號分子來誘導與自身的生物合成有關的基因進行轉錄,這種信號轉導以NisK和NisR為媒介。在B.subtilis中發現了與產nisin菌株Lc. Lactis擁有同樣的群體感應機制,基于基因組序列分析,在枯草芽胞桿菌的雙組分調節系統(TCS04)中,基因對SpaK/SpaR與產nisin菌株的基因對NisK/NisR具有高度的相似性[11]。
基因簇nisABTCIPRKFEG編碼nisin;NisB和NisC是與細胞內翻譯信號有關的修飾反應器蛋白;NisT推斷為ABC轉位分子族中的運輸蛋白;NisP是轉移前導肽的胞外蛋白酶;AI為自身誘導;NisK是跨膜信號轉導蛋白;NisR為反應調節器;NisF、NisE和NisG是一種從細胞到細胞產生免疫性的輸出系統;NisI是促成發射器的脂蛋白[12]。
3 第二類細菌素(ClassⅡAMPs)的合成調節
ClassⅡAMPs(Klaenhammer,1993)是小的熱穩定的不含羊毛硫氨酸殘基的膜活性多肽,它不含有被修飾的氨基酸殘基。作為前體肽的ClassⅡAMPs在合成過程中N-末端的延伸在肽分泌后立即進行。ClassⅡAMPs都具有在分離位點前含有兩個甘氨酸殘基的特征,而不同種類的ClassⅡAMPs對應的分泌機制、免疫機制以及與細菌素產生有關的調節機制已經得到報道,如LactobacillusplantarumC11、LactococcuslactisMG1363、Lactobacillus acidophilusNCFM等菌株在產生細菌素過程中雙組分調節系統的作用。
3.1 Lactobacillus plantarum C11產細菌素的調節
L.plantarumC11產細菌素是一個可誘導的過程,由調節系統中的一個外信息素肽plantaricin A觸發[13]。經由同源的HPK和RR蛋白的作用產生誘導會導致在細菌活性生長期中5個pln操縱子進行協調轉錄,在完全進入穩定期前5個操縱子的轉錄能力開始下降,然后是完全終止轉錄,隨帶的是停止產生細菌素[14]。
3.1.1 與細菌素產生有關的5個操縱子的表達調控
一種稱為基于外信息素肽信號轉換網絡對Lacto-bacillus plantarumC11與產細菌素相關的5個操縱子進行表達調控。在這個調節網絡結構中,外信息素肽(PlnA)誘導細菌素的產生,主要是通過組氨酸蛋白激酶(HPK)和兩種對抗的反應調節器(RR)起作用。細菌素的基因座包含5種操縱子:plnEFI操縱子和plnJKLR操縱子編碼細菌素和免疫蛋白;plnGHSTUV操縱子包含有兩個甘氨酸N-殘基,它作為ABC的轉運系統;操縱子plnABCD作為信號轉換途徑;最后一個操縱子plnMNOP含有一個基因,它在細菌素合成中的功能還不得而知[15]。
在調控操縱子(plnABCD)中,plnA編碼外信息素肽PlnA,plnB編碼HPK,而plnCD編碼兩種RR。所有的pln操縱子在不產生細菌素的條件下都處于被抑制狀態,當PlnA誘導時,所有轉錄的5種操縱子以相互聯系的方式被調節:它們在菌體的對數生長期都具有活性,但有的在穩定期前活性下降。這種表達使5種操縱子促成了一種通常所說的調節網絡[15]。
與Lactobacillus plantarumC11產生細菌素相關的調節操縱子plnABCD所編碼的4種不同蛋白分別是:陽離子前肽(PlnA)、組氨酸蛋白激酶(PlnB)和兩種高度同源性的反應調節器(PlnC和PlnD)(圖4)。成熟的PlnA產物(plantaricin A)作為一種胞外信息素來誘導細菌素的產生,而plnBCD在細菌素產生中所起的具體作用還沒有得到實驗性的證實[2]。
為了證明了plnABCD操縱子編碼自動調節基因使啟動子的活性被激活,用一個包含gusA的指示系統結合plnA啟動子的方法來研究plnABCD;誘導基因plnA和激酶基因plnB產生蛋白需要自體活化。而后幾個基因編碼的蛋白(PlnC和PlnD)作為外信息素肽的感受器,這兩種反應調節器展現出完全不同的功能,與操縱子plnC和plnD過表達影響細菌素產生的機理相一致:PlnC激活轉錄并產生細菌素,而PlnD則抑制這兩種作用即PlnD在細菌素的合成過程中起負調節作用。PlnD是第一種對產生細菌素直接起負調節作用的蛋白,它與PlnC是由位于同一操縱子上的基因編碼的,但卻表現出完全相反的功能特性[2]。
3.1.2L.plantarumC11調節操縱子中plnC和plnD的過表達對細菌素產生的影響
L.plantarumC11的五個負責細菌素產生的pln操縱子的轉錄是被plnA誘導啟動的,一旦被誘導,轉錄在指數生長期的整個過程中發生,但有一些在進入穩定期時立即停止。這意味著存在一個有效的減量調節轉錄機制,操縱子plnD或獨立或結合plnC發揮作用,但在基因活化上被認為是可有可無的,在自身啟動子的活化上更是不需要的。因此懷疑它在細菌素的產生過程中起著與plnC不同的作用,換句話說就是作為一個負調節物。在產細菌素菌株中過分表達plnD來證明其是否抑制plnABCD的轉錄和伴隨的細菌素的產生,所得到的結果與只有plnC對基因活化起作用這一模型相一致[2]。
首先,plnC不管plnD是否一起被表達都表現出十分強烈有效的活性,也意味著這兩個調節器之間沒有明顯的協同作用。其次,當兩個反應調節器各自過表達時,只有plnC的過表達加強了報道基因的表達。一些實驗還表明plnC的功能與HPK的存在有直接的聯系。
這些表明plnC在細菌素的產生過程中起積極的調節作用,相反的,plnD則起負調節作用[2]。
3.2 Lactococcus lactis MG1363產細菌素的調節
在Lactococcus lactisMG1363的染色體中已經鑒定出6種雙組分調節系統,這些雙組分調節系統都包含有一個組氨酸激酶和編碼基因的反應調節器。對6種雙組分調節系統的轉錄分析表明,其中兩個系統顯示出高效的組成性表達,同時剩余的4種在生長期顯示出表達。通過插入誘變確定了兩個組成性表達的雙組分調節系統對于普通細胞的存活和生長以及產生細菌素都是必需的,而通過對剩余4個調節系統的突變分析表明,它們與極端pH值的敏感性、滲透或氧化條件、乳酸乳球菌的磷酸酶活性調節有關[16]。
3.3 Lactobacillus acidophilus NCFM產細菌素的調節
Lactobacillus acidophilusNCFM存在著雙組分寡肽轉換系統,推斷共有九種,其中有一些對細菌素的產生和耐酸性起決定性的作用,每個雙組分系統都是由組氨酸激酶和相應的反應調節器組成。其中有兩個是反應傳感器對,有一對顯現出與細菌素lactacin-B的產生有關,另一對是類似于李斯特菌屬的耐酸性雙組分調節系統,其它雙組分調節系統則對細菌素的產生基本沒有影響[17]。
3.4 Carnobacterium piscicola LV17 B產細菌素的調節
群體感應系統調節Carnobacterium piscicolaLV17B,至少產生兩種細菌素,分別為carnobacteriocin B2 (CB2)和carnobacteriocin BM1(CBM1),主要應用在肉的真空包裝以延長保存期。61 kb的質粒(pCP40)在CB2和CBM1的合成過程中影響很大,precarnobacteriocin B2(cbnB2)與其免疫蛋白(cbiB2)的遺傳因子位于pCP40上,而precarnobacteriocin BM1(cbnBM1)與其免疫蛋白(cbiBM1)的遺傳因子則位于染色體上。部分pCP40經分析含有在細菌素產生過程中所必需的4種基因(cbnKRTD),CbnT和CbnD的功能與細菌的獨立分泌系統有關,而CbnK和CbnR的機能可能是作為雙組分信號轉換系統[9]。
4 結束語
乳酸菌基因組研究的廣泛開展,為人們在分子水平上系統闡述乳酸菌的生理及代謝機制提供了可能[18]。研究者通過生物信息學手段,對已知乳酸菌的基因組進行預測和分析,發現乳酸菌中廣泛存在著雙組分系統,這些雙組分系統中有一些對乳酸菌產細菌素具有直接的調節作用,有一些則通過調控細菌耐酸、抗氧化及耐滲透壓能力來調控細菌素的產生。但對于如何控制外界因素來調節雙組分系統進而調節細菌素的產生還沒有見報道過,因此設想如果能利用外界因素來控制雙組分系統中有利于細菌素產生的成分對于大量工業化生產細菌素具有重大的意義。
[1]CHANG C,STWWART R C.The Two Component System:Regulation of Diverse Signaling Pathways in Prokaryotes and Eukaryotes[J]. Plant Physiol,1998,117(3):723-731
[2]DIEP D,JOHNSBORG O,ARNE P.Evidence for Dual Functionality of The Operon plnABCD in The Regulation of Bacteriocin Production in Lactobacillus plantarum[J].Molecular Microbiology,2001, 41(3):633-644.
[3]NOVICK R P.Regulation of Pathogenicity in Staphylococcus aureus by a Peptide-Based Density-Sensing Mechanism.In Cell–Cell Signaling in Bacteria.Dunny,G.M.,and Winans,S.C.(eds)[J].Washington,DC:American Society for Microbiology Press,1999,pp.129–146.
[4]AZCARATE-PERIL M,MCAULIFFE O,ALTERMANN E,et al.. Microarray Analysis of a Two-Component Regulatory System Involved in Acid Resistance and Proteolytic Activity in Lactobacillus acidophilus[J].Applied and Environmental Microbiology,2005:5794-5804.
[5]邱全勝.雙組分系統——細胞識別滲透脅迫信號的感應器[J].生物化學與生物物理進展,2000,27(6):593-595.
[6]類延寶,宋松泉,傅家瑞,等.植物的雙組分信號系統[J].植物學通報,2004,21(2):216-227.
[7]PFEILER E,ANDREA M,KLAENHAMMER T.Characterization of a Novel Bile-Inducible Operon Encoding a Two-Component Regulatory System in Lactobacillus acidophilus[J].Journal of Bacteriology, 2007:4624-4634.
[8]KLEEREBEZEM M,Quadri L,Kuipers O,et al.Quorum Sensing by Peptide Pherpmones and Two-Component Signal-Transduction Systems in Gram-positive Bacteria[J].Molecular Microbiology,1997,24 (5):895-904.
[9]KLEEREBEZEM M,KUIPERS O,ET AL.A Two-Component Signal-Transduction Cascade in Carnobacterium piscicola LV17B: Two Signaling Peptides and One Sensor-Transmitter[J].Peptides, 2001,22:1597-1601.
[10]KUIPERS O P,BEERTHUIZEN,M.M.,DE RUYTER,ET AL. Autoregulation of Nisin Biosynthesis in Lactococcus Lactis by a Signal Transduction[J].The Journal of Biological Chemistry,1995,270: 27299-27304.
[11]HOLS P,HANCY F,FONTAINE L,ET AL.New Insights in The Molecular Biology and Physiology of Streptococcus Thermophilus Revealed by Comparative Genomics[J].FEMS Microbiology Reviews,2005,29:435-463.
[12]QUARDRI L E N.Regulation of Antimicrobial Peptide Production by Autoinducer-Mediated Quorum Sensing in Lactic acid Bacteria[J]. Antonie van Leeuwenhoek,2002,82:133-145.
[13]DIEP D B,HAVARSTEIN L S.,and NES I F.A Bacteriocin-Like Peptide Induces Bacteriocin Synthesis in Lactobacillus Plantarum C11 [J].Mol Microbiol,1995,18:631-639.
[14]DIEP D B,VARSTEIN H.Characterization of The Locus Responsible for The Bacteriocin Production in Lactobacillus Pantarum C11[J]. J Bacteriol,1996,178:4472-4483.
[15]DIEP D,MYHRE R,JOHNSBORG O,ET AL.Inducible Bacteriocin Production in Lactobacillus is Regulated by Differential Expression of the pln Operons and by Two Antagonizing Response Regulators,the Activity of Which is Enhanced Upon Phosphorylation[J]. Molecular Microbiology,2003,47(2):483-494.
[16]CONNELL-MOTHERWAY M,SINDEREN D,MOREL-DEVILLE F,ET AL.Six Putative Two-Component Regulatory Systems Isolated from Lactococcus lactis subsp.cremoris MG1363[J].Microbiology,2000,146:935-947.
[17]ALTERMANN E,MICHAEL W,ANDREA M,et al.Complete Genome Sequence of the Probiotic Lactic Acid Bacterium Lactobacillus acidophilus NCFM[J].PNAS,2004,102(11):3906-3912.
[18]MAKAROVA K S,KOONIN E V.KOONIN Evolutionary Genomics of Lactic Acid Bacteria[J].J Bacteriol,2007,189(4):1199-1208.
Effect of two-component regulatory system involved in bacteriocin production in lactic acid bacteria
YANG Yu-hong1,WANG Hui2
(1.Heilongjiang Wondersun Dairy Co.Ltd.,Harbin 150078,China;2.Key Laboratory of Dairy Science,Ministry of E-ducation,and Food Science&Technology College,Northeast Agricultural University,Harbin 150030,China)
This review aims at describing the composition of two-component regulatory systems and the regulation of two-component regulatory systems in two class bacteriocin synthesis,understanding the mechanism of two-component regulatory systems involved in bacteriocin production inlactic acid bacteria.
two-component;bacteriocin;histidine protein kinase;response regulators
Q936
B
1001-2230(2011)06-0051-04
2011-03-01
楊郁葒(1968-),女,工程師,從事發酵乳制品的研究與開發。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
鴨綠江(2021年35期)2021-04-19 12:24:18
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
裝備制造技術(2019年12期)2019-12-25 03:06:46
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
家庭影院技術(2017年9期)2017-09-26 03:41:45
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
