軸突生長抑制因子重組疫苗預防免疫減輕阿爾茨海默癥模型小鼠行為學損傷
2011-02-01 09:28:34張玲玲馬全紅楊武林馬春梅徐艷峰肖志成
中國比較醫學雜志 2011年6期
張玲玲,馬全紅,楊武林,劉 穎,馬春梅,黃 瀾,徐艷峰,朱 華,鄧 巍,肖志成,秦 川
(1.北京協和醫學院比較醫學中心,中國醫學科學院醫學實驗動物研究所,衛生部人類疾病比較醫學重點實驗室;北京協和醫學院比較醫學中心,中國醫學科學院醫學實驗動物研究所,國家中醫藥管理局人類疾病動物模型三級實驗室,北京 100021;2.蘇州大學神經科學研究所,蘇州 215123;3.蒙納士大學免疫干細胞實驗室,維多利亞,澳大利亞 3800;4.昆明醫學院分子臨床醫學研究所,昆明 650031)
阿爾茨海默癥(A lzheimer's disease,AD)又名老年癡呆癥是引發癡呆的最常見疾病。隨著人口老齡化的加重,阿爾茨海默癥越來越受到人們的重視。雖然目前有大量阿爾茨海默癥的相關研究報道但并未闡明該病病因機理,而一系列藥物也并不能起到實質性治療作用。軸突損傷,神經元丟失不可再生是老年癡呆癥患者學習記憶漸進性損傷并最終導致癡呆的直接原因。而腦內神經元不可再生主要是由腦內環境造成的,其中髓鞘相關軸突生長抑制因子蛋白(NGI)在抑制神經元,軸突再生過程中扮演重要角色[1-3]。NGI是一系列以抑制軸突生長為主要功能的蛋白。軸突生長抑制因子蛋白重組疫苗能夠促進脊髓橫斷損傷小鼠軸突生長,恢復脊髓功能[4-5],該研究結果為該疫苗在神經退行性變疾病中的應用提供了可能。基于以上研究,本研究構建NGI重組疫苗并觀察NGI重組疫苗對阿爾茨海默癥轉基因模型小鼠的預防效果。
1 材料和方法
1.1 質粒
NGI重組DNA質粒包含小鼠 Tenascin-R(TNR)和Tenascin-C(TN-C)蛋白的表皮生長因子樣區域,以及人受體樣蛋白酪氨酸磷酸酶β的cDNA,每個亞克隆序列末端加入三個丙氨酸作為連接[6]。以上三個片段順次連接后經 Bg1II和 XbaI酶切位點插入 pcDNA3.1+載體(pcDNA-NGI),產物經過PCR和測序驗證。PcDNA3.1+和pcDNA-NGI質粒在大腸桿菌中擴增,質粒大提試劑盒(美國Promega公司)提取無內毒素質粒,制備成1μg/μL溶解在磷酸鹽溶液中,-20℃凍存備用。
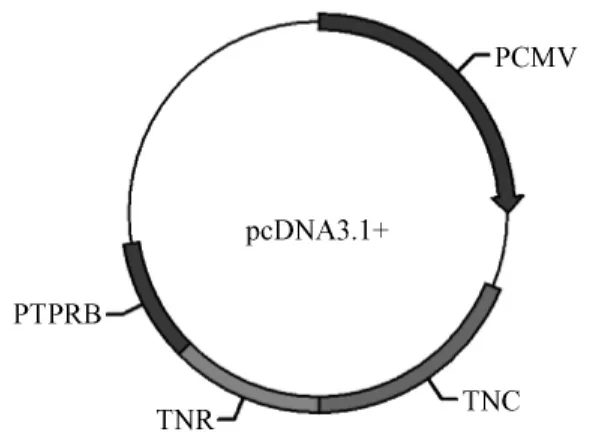
圖1 重組DNA疫苗示意圖Fig.1 recombinant DAN vaccine
1.2 H is蛋白的表達及純化
設計帶有Pet28載體(李萬波教授饋贈)限制性酶切位點的 Tenascin R引物,構建質粒 Pet28-Tenascin R,將質粒轉染入 BL21(DE3)細胞,LB固體培養基過夜培養,挑取單克隆菌落加入 LB液體培養基,37℃振蕩培養至OD值為0.6~0.9,加入終濃度1 mM IPTG,25℃誘導6~8 h,超聲破碎菌體,SDS-PAGE電泳分析蛋白表達。GE 1m L HP His蛋白純化柱純化蛋白。
1.3 實驗動物及分組
APP/PS1轉基因小鼠由我所遺傳中心實驗室構建,該轉基因小鼠三月齡出現認知行為學改變,4.5月腦內出現老年斑沉積[7-8]。無任何行為學改變及腦內老年斑沉積的4~5周小鼠隨機分為四組:野生對照組(注射PBS);模型對照組(注射PBS);空載體對照組(注射pcDNA3.1質粒);疫苗組(注射pcDNA-NGI)。每組12只,雌雄各半。實驗動物飼養室溫18℃ ~25℃,相對濕度(30% ~70%),每日光照12 h,自由攝食引水,實驗動物使用及操作均嚴格按照中國醫學科學院實驗動物倫理委員會標準進行(證號ILAS-PL-2010-003)。
1.4 免疫和抗體測定
小鼠免疫采用DNA疫苗肌肉注射免疫,具體如下:4~5周小鼠肌肉注射質粒100μg/只,兩周一次,連續免疫直至實驗結束。四只小鼠用來進行抗體檢測,免疫前及免疫后每兩周取血,分離血清,-80℃凍存,檢測抗體使用。
抗體檢測采用雙夾心Elisa法,具體步驟簡述如下:純化蛋白his-TNR 10 ng/孔包被板(美國corning公司)4℃過夜孵育,5%脫脂奶粉與0.5%BSA混合液37℃1 h封閉,血清樣品1∶200稀釋,37℃1 h孵育,山羊抗小鼠二抗(中衫金橋PS-6002)室溫1 h孵育,TBS發光液室溫孵育15 min,含有2M硫酸的終止液終止顯色,酶標儀450 nm讀數。
1.5 行為學檢測:M orris水迷宮
利用荷蘭Noldus ethvision XT行為學分析軟件及監測系統對小鼠學習記憶能力進行測試與數據分析。Morris水迷宮 DMS-2系統由中國科學院藥物研究所提供,包括一個圓形不銹鋼水池,直徑100 cm,高50 cm;將水池等分為4個象限;目標象限的中央放置一直徑為9 cm,高27 cm的圓形隱藏平臺,整個實驗期間其位置保持不變;水池中水面高于平臺且加入奶粉,以隱藏平臺并使水不透明,水溫保持(26±1)℃;實驗期間迷宮外設足夠的參照物,且始終保持不變。迷宮上方安置帶有顯示系統的攝像機,同步記錄小鼠運動軌跡,Morris水迷宮數據采集和分析軟件記錄相關數據及圖象結果。整個實驗過程分為隱藏平臺獲得實驗和空間搜索實驗兩部分。
1.5.1 隱藏平臺獲得實驗:用于測量動物在水迷宮中的學習和記憶能力。實驗歷時5 d。正式實驗每天訓練4次,每次60 s,隨機從東、西、南、北4個入水點選擇一個,將動物面向池壁放入水中,記錄動物尋找并爬上平臺所需時間即潛伏期(latency)。如果動物在60 s內未找到平臺,由實驗者將其引至平臺,潛伏期記為60 s;動物在平臺上停留20 s(此種停留對在水中高度緊張的小鼠非常重要,它可避免小鼠再跳回水中,以保證實驗的成功)。每次訓練之間和4次訓練完成后,迅速將動物洗凈擦干,置于加熱器旁,以防動物低體溫。每只動物共計訓練20次。計算每天各組4次逃避潛伏期的平均值。
1.5.2 空間搜索實驗:用于測量動物對平臺空間位置準確記憶,即記憶保持能力。第6天撤除平臺,任選一個入水點將動物放入水中,動物在水中游泳60 s,測量60 s內:(1)動物在目標象限(原平臺所在象限)(target quadrant)的游泳時間;(2)穿越目標(原平臺所在區域及象限)次數。Noldus ethvision XT行為學分析軟件記錄數據后,采用SPSS13.0統計學軟件進行數據分析。
2 結果
2.1 抗體檢測
在裸DNA免疫兩次后抗體OD平均值由免疫前的0升至0.3并一直維持在0.3左右直至實驗結束(圖2)。結果顯示裸DNA疫苗注射在整個實驗過程都產生了有效的抗體。
2.2 水迷宮檢測
5天的隱藏平臺獲得實驗中轉基因組和空載體組潛伏期雖然在第2,3天縮短,但第4,5天的訓練卻出現潛伏期再次延長的現象,表明其學習能力受損。疫苗組潛伏期在第3天出現平臺期(較第2天并無縮短)但隨著訓練次數的增加在第4,5天顯著縮短并較模型對照組和空載體對照組有顯著差異(P<0.05),疫苗組潛伏期隨訓練天數變化趨勢與野生組一致,表明疫苗對癡呆小鼠的學習能力起到了改善作用(圖3)。在探索實驗中,轉基因和空載體組穿越原平臺及原平臺所在象限的次數,停留時間均較野生組縮短表明其記憶功能受損。疫苗組穿越次數增加停留時間延長表明預防免疫能夠改善APP/PS1的記憶功能(圖3)。
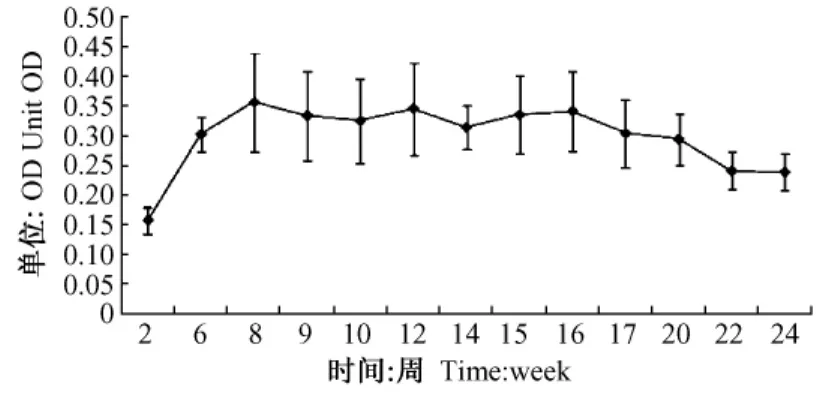
圖2 抗體動態檢測示意圖Fig.2 Fig.re of antibody
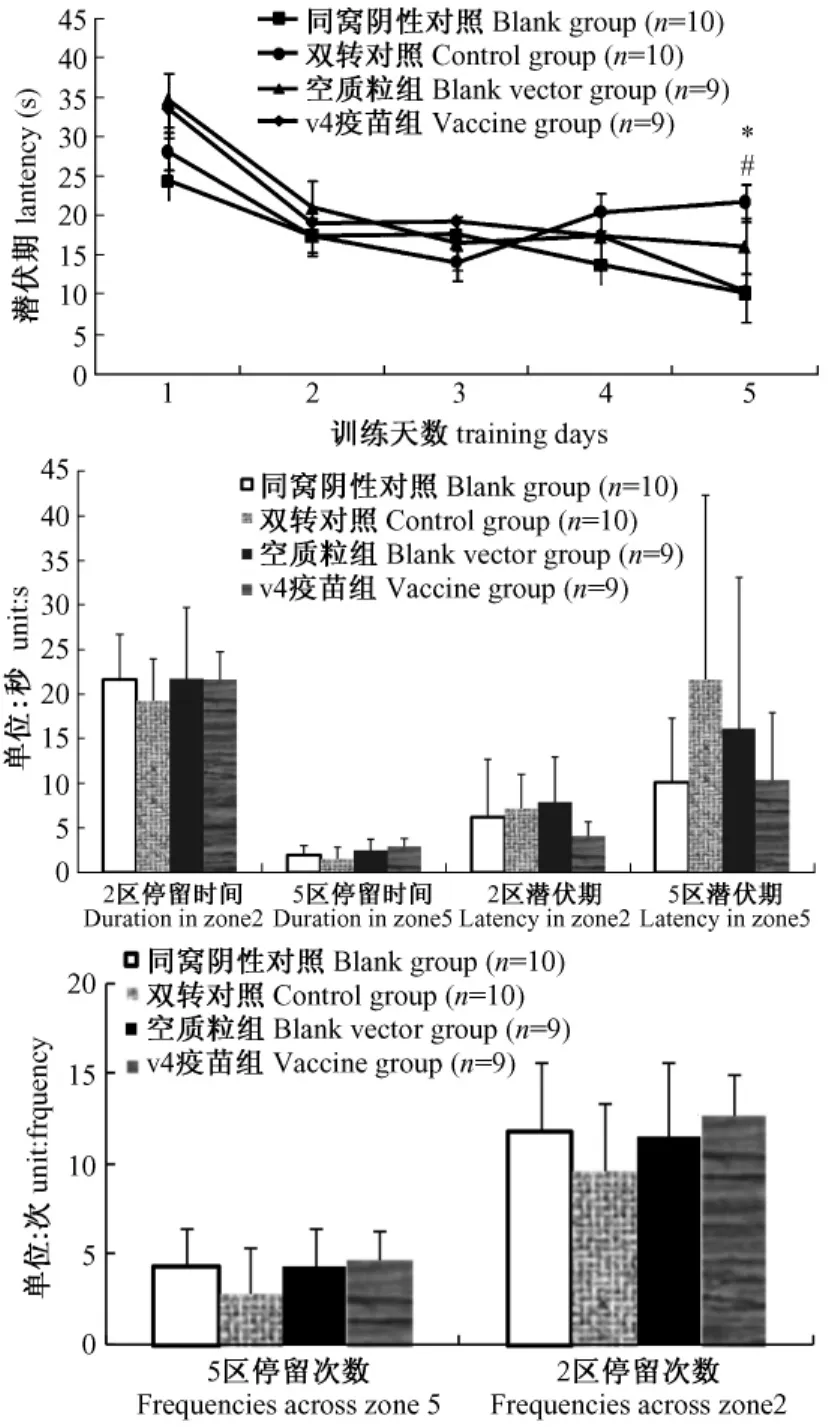
圖3 Morris水迷宮實驗結果(*P<0.05,P<0.01野生對照組VS雙轉對照組;#P<0.05空質粒對照組VS疫苗組)Fig.3 Open field test result(*P<0.05,WT group vs placebo group;#P<0.05,vehicle vs vaccine group)
3 討論
阿爾茨海默癥是以神經元丟失為病變基礎的神經退行性變疾病,神經元的損傷不可再生是導致病人出現各種癥狀的根本原因。神經元的不可逆性損傷大部分歸咎于髓鞘及少突膠質細胞相關的軸突生長抑制因子蛋白。Tenascin R,Tenascin C,受體樣蛋白酪氨酸磷酸酶β,均為髓鞘相關軸突生長抑制因子蛋白[11-12]。髓鞘相關軸突生長抑制因子重組疫苗能夠應用于神經退行性疾病治療[1-3,13],由MAG Ig1-5區域,TN-R的EGFL區域,NogoA的66氨基酸胞外區域及N端構成的DNA疫苗能在大鼠體內產生有效抗體,促進脫髓鞘大鼠的皮質脊髓束軸突再生[4],增加脊髓損傷大鼠體內神經生長因子蛋白的表達[5],并無脫髓鞘副反應的產生[4]。本文中我們檢驗了軸突生長抑制因子疫苗是否也能有效的改善阿爾茨海默癥的癥狀。DNA疫苗便于構建,生產方便,且DNA疫苗注射可產生有效抗體[4]。我們采用裸DNA疫苗連續注射的免疫方式。實驗結果表明此方法在免疫期間產生了雖然不高但持續有效的抗體。APP/PS1雙轉小鼠是由過表達人的 APP swedon雙突變(K594N/ M595L)和 PS1單突變(deltaE9)小鼠雜交而成,兩種突變均為家族性老年癡呆癥的患病基因,雙轉小鼠的老年斑隨年齡的增長而不斷增多,學習記憶損傷也不斷加重[7-8,9]。行為學檢測方法水迷宮實驗表明疫苗能有效改善模型小鼠的行為學損傷。到目前為止針對阿爾茨海默癥的治療藥物只能取得癥狀的改善,疫苗治療也多聚焦于 Aβ蛋白的被動或主動免疫以期待減少Aβ蛋白的表達,而對于與老年癡呆癥狀學習記憶損傷直接相關的軸突改善卻被人忽略。本文中髓鞘相關軸突生長抑制因子重組疫苗預防免疫能改善模型小鼠的行為學,可能機制推測為該疫苗預防阿爾茨海默癥導致的軸突損傷,為阿爾茨海默癥的免疫治療提供了新契機。其具體機制尚待我們進一步實驗研究證實。本實驗中DNA免疫雖產生有效抗體,但需連續免疫,為疫苗發展前景帶來阻礙,我們會進一步在免疫劑量,方式方面進行改善,期待得到更加有效簡便的免疫方式。另外,本實驗中的疫苗免疫在小鼠產生行為學癥狀前進行,至于該疫苗是否對小鼠有治療效果還需我們進一步實驗證實。
致謝:感謝李萬波教授、劉江寧博士對實驗中質粒構建及蛋白表達純化的幫助和指導。
[1]Ang BT,Xu G,Xiao ZC.Therapeutic vaccination for central nervous systemrepair[J].Clin Exp Pharmacol Physiol,2006,33 (5-6):541-5.
[2]Ma QH,Yang WL,Nie DY,et al.Physiological roles of neurite outgrowth inhibitors in myelinated axons of the central nervous system——implications for the therapeutic neutralization of neurite outgrowth inhibitors[J].Curr Pharm Des,2007,13 (24):2529-37.
[3]Nie DY,Xu G,Ahmed S,et al.DNA vaccine and the CNS axonal regeneration[J].Curr Pharm Des,2007,13(24):2500-6.
[4]Gang Xu,Duyu Nie,Jutao Chen,et al.Recombinant DNA vaccine encoding multiple domains related to inhibition of neurite outgrowth:a potential strategy for axonal regeneration.[J].Neurochem,2004,91:1018-1023.
[5]Zhang Y,Hao CG,Hu LQ,et al.Recombinant DNA vaccine against inhibition of neurite outgrowth promotes functional recovery associated with endogenous NGF expression in spinal cord hemisected adult rats[J].Neurochem Res.2009 Sep;34 (9):1635-41.
[6]高偉,張玲玲.NGI重組DNA疫苗對轉基因AD小鼠的治療作用[J].中國比較醫學志,2009,19(6):13-16.
[7]Jankowsky JL;Fadale DJ;Anderson J et al.Mutant presenilins specifically elevate the levels of the 42 residue beta-amyloid peptide in vivo:evidence for augmentation of a 42-specific gamma secretase[J].Hum Mol Genet,2004,13(2):159-70.
[8]宗園媛,王曉映.APP/PPS雙轉基因阿爾茨海默病小鼠模型的老年斑及行為學動態分析[J].中國比較醫學雜志,2008,18(9):8-12.
[9]袁樹民,張曉娟.阿爾茨海默病模型小鼠自主活動能力的分析[J].中國比較醫學雜志,2010,20(4):5-8.
[10]Zhao Y,Pei J,Ji Z,et al.Effect of amyloid precursor protein 17mer peptide on microtubule structure and tau protein hyperphosphorylation in hippocampal neurons of experimental diabetic m ice[J].NeuroReport,2003,14(20):61-66
[11]Prinjha R,Moore SE,Vinson M,et al.Inhibitor of neurite outgrowth in humans[J].Nature,2000,403(6768):383-4.
[12]Pesheva P,Spiess E,Schachner M.J1-160 and J1-180 are oligodendrocyte-secreted nonperm issive substrates for cell adhesion[J].JCell Biol,1989,109(4):1765-1778.
[13]Strittmatter SM. Modulation of axonal regeneration in neurodegenerative disease:focus on Nogo[J].JMol Neurosci,2002,19(1-2):117-21.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
七彩語文·畫刊(2012年3期)2012-04-29 00:00:00
