G蛋白偶聯受體56胞外端融合蛋白的表達、純化及其多克隆抗體的制備①
2011-02-06 04:38:20趙去非韓紅輝劉俊晨潘新華錢杜
中國免疫學雜志 2011年5期
趙去非 韓紅輝 劉俊晨 潘新華 汪 磊 錢杜 冰
(華東師范大學生命醫學研究所,上海 200241)
G蛋白偶聯受體56胞外端融合蛋白的表達、純化及其多克隆抗體的制備①
(華東師范大學生命醫學研究所,上海 200241)
目的:制備G蛋白偶聯受體56(G protien-coupled recepter 56,GPR56)的高效抗體,為其功能及分子機制的深入研究提供靈敏、高效的免疫學檢測工具。方法:以肝癌細胞株MHCC-97H的cDNA為模板,利用PCR技術擴增GPR56 N端序列(661~1 073 bp),并構建重組表達載體pGEX4T-1-GPR56N。陽性克隆經酶切、測序鑒定正確后,轉化大腸桿菌BL21進行誘導表達。表達產物經Western blot初步鑒定后,用GST磁珠分離純化,純化后蛋白與免疫佐劑混合直接免疫家兔制備抗血清。陽性血清用ELISA、Western blot、免疫組織化學等技術對細胞或組織中的GPR56進行鑒定和分析。結果:成功構建重組表達載體pGEX4T-1-GPR56N,GPR56N與GST的融合蛋白(GST-GPR56)在原核系統中進行了高效的可溶性表達,制備的兔抗人GPR56抗體經Western blot和免疫組織化學分析結果表明,該抗體可同時對人源性和鼠源性的GPR56進行特異性檢測。結論:成功對GPR56蛋白N端序列進行了原核表達和純化,制備的抗GPR56多克隆抗體具有較高的特異性,為GPR56功能及分子機制的深入研究奠定了堅實的基礎。
GPR56;GST;原核表達;多克隆抗體
G蛋白偶聯受體(GPCRs)是真核生物中最大的一類跨膜受體家族,目前已報道的GPCRs多達2 000余種。這些結構各異功能豐富的GPCRs通過與相應的配體結合,能夠將細胞外的各種生物信息跨膜傳遞到細胞內,引起細胞內相應信號通路的變化,最終完成對細胞生理功能的調節,從而引起包括細胞生長、分化、凋亡、遷移以及腫瘤等生長在內的一系列生物學功能[1,2]。其中,G蛋白偶聯受體超家族中粘附亞家族所含成員超過30個[3],該家族有以下兩個特點:①胞外段即N端很長,含有很多不同的功能區域;②具有富含半胱氨酸的GPCR水解位點(簡稱GPS)。因此,G蛋白偶聯受體粘附亞家族在細胞間的粘附,以及細胞基質與細胞內的信號轉導過程中發揮了非常重要的作用[4,5]。
GPR56是GPCRs粘附家族中的重要成員之一,屬于GPCRs中類分泌素家族。它除了具有粘附家族成員普遍具有的特征之外,在胞外區段還含有一個類粘蛋白功能域。近期對GPR56功能研究表明,它可能在人體機能紊亂、腫瘤發生等疾病中發揮重要作用。最新研究發現,GPR56在遷移性黑色素瘤中的表達下調,通過相關的體內實驗證明,GPR56對黑色素瘤的生長和轉移有明顯的負調控作用[6,7]。
本文通過PCR的方法從人cDNA文庫中克隆出GPR56N端約400 bp大小的基因片段,并將其連入表達載體進行原核表達,融合蛋白經蛋白純化后免疫家兔得到多克隆抗體,制備得到的陽性血清可用于Western b lot、免疫組織化學等技術,能夠同時對人類和小鼠來源的細胞和組織中的GPR56進行特異性檢測,為GPR56的功能和作用機制的深入研究提供了良好的研究工具。
1 材料與方法
1.1 主要試劑和儀器 rTaq酶、4×dNTP預混液、限制性內切酶SalⅠ、SacⅠ、NotⅠ、T4DNA連接酶均購自大連寶生物工程有限公司。pGEX4T-1,大腸桿菌DH5α和BL21菌株為本實驗室保存。瓊脂糖凝膠DNA回收試劑盒、質粒DNA小量提取試劑盒購自天根生物公司。基因測序和引物合成由上海博尚生物技術有限公司完成。異丙基硫代-β-半乳糖苷(IPTG)購自德國Merck公司。弗氏佐劑購自美國Sigma公司。ELISA試劑盒購自美國BD生物公司。人 肝 癌 細 胞 株 Huh-7、SMMC-7701、MHCC97L、MHCC97H、SK-Hep1、BEL-7402等均購自中國科學院上海細胞庫。GPR56標準抗體購自Abcam(ab77515)。臨床組織芯片購自上海芯超生物科技有限公司。
1.2 方法
1.2.1 GST-GPR56重組蛋白的表達、純化及鑒定根據NCBI提供的GPR56的DNA序列,應用生物信息學軟件primer5.0設計上下游引物,擴增人源GPR56基因中661~1 073 bp片段。其上游引物及下游引物均設計為SalⅠ酶切位點,上游引物為5′-GCACCAGGAGGAGAGCCTGGCTCAG-3′,下游引物為5′-CGCTCTGCTCCTCCTCCTGCCGGGA-3′。 以肝癌 細胞MHCC-97H的cDNA為模板進行PCR擴增,PCR擴增條件為:94℃預變性5分鐘;94℃變性30秒,55℃退火30秒,72℃延伸 45秒 (30個循環);72℃再延伸8分鐘。瓊脂糖凝膠電泳并回收擴增的目的片段。回收片段及pGEX4T-1載體經SalⅠ酶切后,以適當比例將目的片段與載體連接并轉化大腸桿菌DH5α,陽性克隆經測序證明插入片段序列正確后抽提質粒,轉化大腸桿菌BL21。400 ml低鈉LB中加入2.5ml活化菌液,搖至OD600在0.4~0.6。加誘導劑IPTG(0.5mmol/L),低溫25℃誘導表達6小時。取大量表達后的菌液4℃4 000 r/min離心15~30分鐘;加含PMSF(終濃度 0.2 nmol/L)的 PBS 10m l重懸菌體,超聲破碎后,加0.5%Triton X-100,4℃混合30分鐘,12 000 r/min離心30分鐘,將上清與GST磁珠混合,4℃孵育2小時。用含 0.1%Triton的PBS洗磁珠(洗掉非特異吸附),4℃冰箱混合10分鐘,3 000 r/min離心5分鐘,重復洗四次,最后一次離心留少許上清。用谷胱甘肽洗脫液洗脫。取GST蛋白和GST-GPR56融合蛋白各2μl,SDS-PAGE電泳后用抗GST抗體檢測目的蛋白。
1.2.2 兔抗GST-GPR56N融合蛋白抗體的制備將一定量重組GPR56N-GST融合蛋白與等體積弗氏完全佐劑充分混勻并乳化后,每只家兔皮下多點注射500μg,4周后劑量減半進行再次免疫(不完全佐劑),4周后同樣條件加強免疫,1周后頸動脈取血,血漿于37℃放置2小時,4℃沉淀過夜后,離心分離血清,分裝后置-20℃保存。
1.2.3 兔抗人GPR56N抗體效價的測定 ELISA檢測GPR56抗血清效價:檢測板用GST-GPR56融合蛋白(1mg/L)包被,每孔100μl,4℃過夜。每孔加入200μl封閉液,37℃封閉2小時。每孔加入100μl以1∶4梯度稀釋的抗血清(陰性對照為正常兔血清,空白對照為封閉液),37℃孵育30分鐘。加入HRP標記羊抗兔IgG,37℃孵育30分鐘。TMB顯色后,酶標儀以450 nm波長讀值。
1.2.4 兔抗人GPR56融合蛋白抗體特異性的Western blot分析 將含有hGPR56胞外段序列的真核表達質粒和對照空質粒(pCDNA3.1)脂質體法轉入293T細胞,使其在293T細胞中過表達,收集細胞后加入適量SDS上樣buffer,100℃煮沸十分鐘,經SDSPAGE分離,并轉移至PVDF膜上,以3%脫脂奶粉于室溫封閉1小時后,加入1∶1 000稀釋的兔抗GPR56融合蛋白抗血清,室溫溫育2小時,以PBST室溫洗膜3次,每次 10分鐘,之后,加入 1∶2 000稀釋的HRP標記的羊抗兔IgG作為二抗溫育1小時,洗膜3次,每次10分鐘。顯影曝光。
1.2.5 兔抗人GPR56抗體的免疫組織化學分析將人肝癌臨床組織芯片樣品和BALB/c模型小鼠動脈血管組織石蠟切片,常規脫蠟,置0.01mol/L檸檬酸鹽緩沖液(pH 6.0)中抗原修復,分別滴加0.3%H2O2甲醇溶液,室溫避光孵育10分鐘,經BSA封閉后,用制備的抗GPR56多抗濕盒內孵育30~60分鐘,洗片,顯微鏡下控制DAB顯色(2~5分鐘),自來水沖洗25分鐘,蘇木精復染,封片,鏡檢、觀察。
2 結果
2.1 原核表達載體pGEX4T-1-GPR56的構建與鑒定經PCR擴增得到GPR56N端片段(661~1 073 bp)大小約為400 bp(圖1)。目的片段經SalⅠ酶切后,連入pGEX-4T-1質粒,并轉化DH5α。抽提質粒后采用目的片段781位置上SacⅠ和載體中NotⅠ位點進行酶切,酶切片段大小為300 bp左右。如圖2中質粒1、2為陽性克隆,與預期大小一致。
2.2 融合蛋白GST-GPR56的誘導表達及表達形式分析 經測序鑒定正確的質粒,轉化BL21后IPTG誘導4小時,超聲破碎后用GST磁珠純化可溶性融合蛋白。表達出的GST、GPR56 N端融合蛋白分子量約為41 kD。GST抗體Western blot(圖3)檢測目的蛋白,結果表明融合蛋白分子量與預期基本一致。
2.3 ELISA檢測兔抗人GPR56抗體效價 用重組GPR56N-GST蛋白包被檢測板,ELISA檢測兔抗人GPR56血清的效價為1∶10 000,Western blot分析中有效的稀釋比例為1∶1 000。
2.4 Western blot檢測兔抗人GPR56抗體特異性與對照未轉入GPR56N的細胞(圖4,泳道2)相比,轉入pEGFP-N2-GPR56質粒的293T細胞(圖4,泳道1),經我們制備的抗體檢測,在40 kD左右明顯出現一條特異性的蛋白條帶,其分子量與GPR56N端融合蛋白預期大小基本一致,表明抗體對GPR56的特異性結合能力(圖4)。
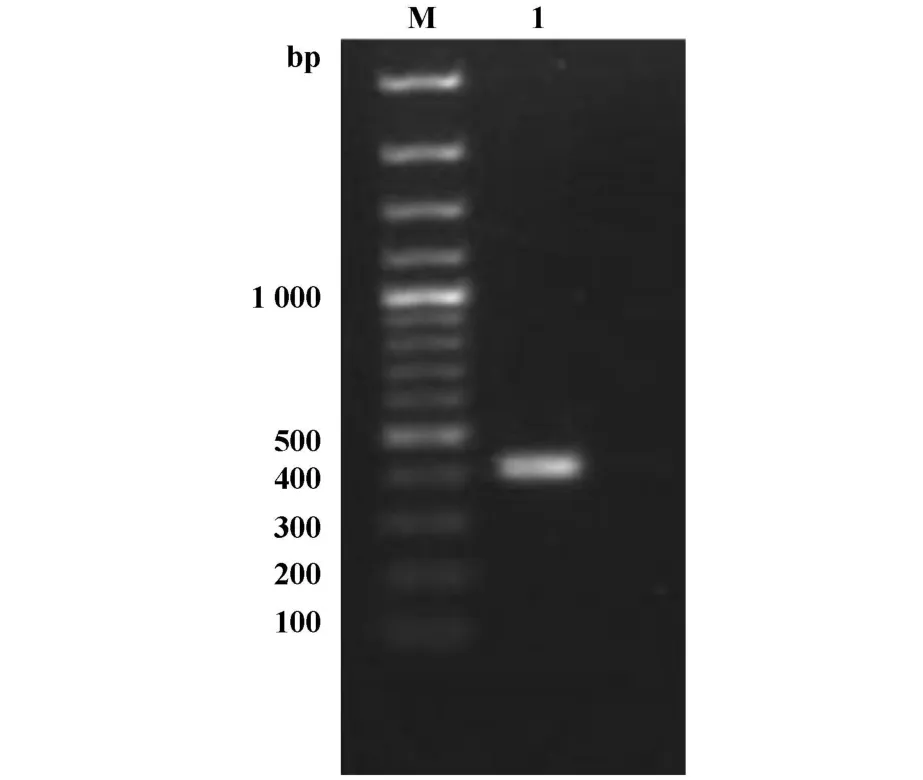
圖1 GPR56 N端片段PCR擴增結果Fig.1 PCR amplification of GPR56 N-term inal
2.5 兔抗人GPR56抗體檢測不同肝癌細胞系中GPR56的表達 收集人肝癌細胞系Huh-7、SMMC-7701 、MHCC97L 、MHCC97H 、SK-Hep1 、BEL-7402 的細胞裂解物,利用所制備的抗體進行Western blot檢測時發現,不同肝癌組織中GPR56的表達出現明顯差異(圖5),且所示條帶與所購買的商業化GPR56抗體(Abcam)的檢測結果基本一致。

圖2 pGEX4T-1-GPR56N酶切檢測Fig.2 Identification of recombinantp lasmid pGEX4T-1-GPR56N

圖3 GPR56N端融合蛋白Western blot分析Fig.3 W estern blot analysis for GPR56N term inal fusion p roteins

圖4 兔抗人GPR56抗體鑒定Fig.4 Identification of the rabbit anti-GPR56 polyclonal antibody by Western b lot

圖5 多克隆抗體anti-GPR56識別不同細胞系中的GPR56蛋白Fig.5 Identification of endogenous GPR56 from different cell lines using polyclonal antibody anti-GPR56
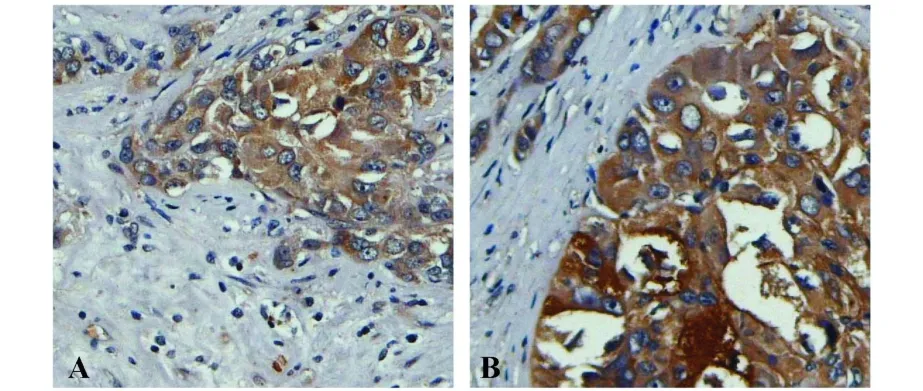
圖6 GPR56抗體對肝癌臨床組織芯片中的人源性的GPR56的檢測Fig.6 Immunohistochemistry analysis exp ression of GPR56 in liver cancer tissuem icroarray using polyclonalantibody anti-GPR56
2.6 兔抗人GPR56抗體檢測臨床組織芯片中GPR56的表達 為進一步驗證該自制兔抗人GPR56的有效性,我們選取自制抗體與購買的GPR56抗體,對肝癌臨床組織芯片中人源性的GPR56進行了檢測(圖 6)。從圖中可知,我們自制的兔抗人GPR56抗體(圖6B)與購買的GPR56抗體(圖6A)在組織切片中的著色部位基本相同,證明我們所制備的兔抗人GPR56抗體的特異性。
2.7 免疫組織化學檢測GPR56在鼠動脈血管中的表達 利用RT-PCR的方法我們也發現GPR56在血管內皮細胞中表達豐富(圖7),并有數據表明,人、小鼠和大鼠中GPR56的同源性達到88%。因此,我們取小鼠動脈血管,用免疫組化的方法檢測小鼠血管中鼠源GPR56的表達情況(圖 8)。同樣發現GPR56在小鼠動脈血管中的表達較為豐富,提示我們GPR56可能在血管新生和發育過程中扮演著重要角色。

圖7 GPR56在內皮細胞中的表達Fig.7 Expression of GPR56 in endothelial cell
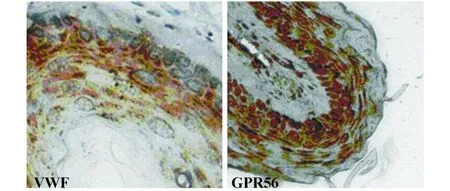
圖8 免疫組化檢測GPR56在鼠動脈血管中的表達Fig.8 Immunohistochem istry analysis expression of GPR56 in mouse arterial blood vessel using polyclonal antibody anti-GPR56
3 討論
大約40%~50%的臨床藥物都以GPCRs為治療靶點,因此對于GPCRs功能及分子機制的研究已經成為了目前的熱點[8]。GPCRs粘附家族作為其重要的家庭成員,其功能受到越來越多的關注,然而到目前為止大部分家庭成員,尤其是一些孤兒受體的功能仍不清楚。GPR56是近年研究較多的粘附家族成員之一,目前已發現其在腫瘤發生、惡化過程中發揮了至關重要的作用。有文獻報道,GPR56的作用機制是通過與細胞表面的CD9/CD81形成一個多分子復合體,從而發揮作用[9]。最近證實,一種普遍表達的細胞外基質蛋白轉谷氨酰胺酶(TG2)可能為GPR56的配體。有文獻證明,TG2能夠抑制血管新生和腫瘤生長,TG2被敲除的小鼠更容易發生腫瘤,它能與整合素、纖維連接蛋白等基質蛋白形成大的復合物,從而抑制腫瘤生長和遷移[10-13]。因此開展對GPR56詳細功能機制的研究具有十分重要的理論價值和應用前景。
抗體在蛋白質檢測及功能研究中都有著廣泛的應用,制備高效價的抗體是研究基因表達及其功能的重要工具。又由于G蛋白偶聯受體是一種具有7次跨膜結構的細胞膜蛋白,且胞外端結構復雜并伴隨有較高程度的糖基化修飾,同時胞外端的結構還起到了識別特異性配體的作用,因此這些G蛋白偶聯受體的特殊結構造成了其抗體制備的困難。本文以GPR56蛋白N端為目標,在原核表達系統中重組表達了GPR56-GST融合蛋白,并以該蛋白為免疫原,成功制備了特異性高和效價良好的兔抗人GPR56多克隆抗體。該抗體可同時識別內源性及外源性的GPR56,并能夠用于 ELISA、Western blot以及免疫組織化學的檢測,為今后深入研究GPR56的表達分布、細胞內定位及其與其它蛋白質間的相互作用提供了一個有用的檢測與鑒定工具。由于小鼠和人GPR56蛋白具有一定的同源性,免疫組織化學染色顯示該多抗不僅能夠識別人源性GPR56,還能夠識別小鼠體內GPR56,可用于對小鼠組織GPR56蛋白表達和定位研究,具有比較廣泛的應用價值。該抗體的獲得不僅能夠解決GPR56蛋白檢測的問題,還可能對相應配體結合機制的研究提供幫助,具有重要的應用前景,同時為進一步研究GPR56在多種疾病發生、發展過程中的作用和分子機制提供了必要的工具。
1 Bockaert J,Pin JP.Molecular tinkering of G protein coupled receptors:an evolutionary success[J].EMBO,1999;18:1723-1729.
2 Pierce K L,Premont R T,Lefkowitz R J.Seven-transmembrane receptors[J].Nat RevMolCell Biol,2002;3:639-650.
3 Fredriksson R,Gloriam D E,Hoglund PJetal.There existat least 30 human G-protein coupled receptors with long Ser/Thr-rich N-termini[J].Biochem Biophys Res Commun,2003;301:725-734.
4 Hayflick JS.A family ofheptahelical receptorwith adhesion-like domains:amarriagebetween two super fam ilies[J].Recept Signal Transduct Res,2000;20:119-131.
5 Van Lier RA,EichlerW,Hamann Jetal.The sevenspan trnasmembrane receptor CD97 has a cellular ligand(CD55,DAF)[J].Exp Med,1996;184:1185-1189.
6 Shashidhar S,Lorente G,Nagavarapu Uetal.GPR56 is a GPCR that is overexpressed in gliomas and functions in tumor cell adhesion[J].Oncogene,2005;24:1673-1682.
7 Xu L,Begum S,Hearn JDetal.GPR56,an atypicalG protein-coupled receptor,binds tissue transglutaminase,TG2,and inhibitsmelanoma tumor growth and metastasis[J].Proc Natl Acad Sci USA,2006;103:9023-9028.
8 Fredriksson R,Schioth H B.The repertoire of Gprotein coupled receptors in fully sequenced genomes[J].Mol Pharmacol,2005;67:1414-1425.
9 Little K D,HemlerM E,Stipp CSetal.Dynam ic regulation of a GPCR-tetraspanin-G protein complexon intact cells:central roleofCD81 in facilitating GPR56-Galphaq/11association[J].MolBiolCell,2004;15:2375-2387.
10 Lorand L,Graham R M.Transglutaminases:crosslinking enzymes with pleiotropic functions[J].Nat Rev Mol CellBiol,2003;4:140-156.
11 Iismaa SE,BeggG E,Graham RM.Cross-linking transglutaminaseswith G protein coupled receptor signaling[J].SciSTKE,2006;353:34-36.
12 Akimov SS,Krylov S,Fleischman LFetal.Tissue ransglutaminase isan integrin-binding adhesion coreceptor for ibronectin[J].JCell Biol,2006;148:825-838.
[收稿2010-11-29 修回2011-01-27]
(編輯 許四平)
Expression,purification and polyclonal antibody p reparation of G protein-coupled receptor 56(GPR56)extracellular domain fusion protein
ZHAOQu-Fei,HANHong-Hui,LIUJun-Chen,PANXin-Hua,WANGLei,QIANMin,DUBing.EastChinaNormal University,InstituteofBiomedicalScience,Shanghai200241,China
Objective:To prepare ahigh efficacious and sensitive polyclonal antibody against the N-terminal of human G protein-coupled receptor 56(GPR56)for the further functionalassay.Methods:The cDNA of human liver cancer cell line MHCC-97H was used as the templateto amplify the N-terminal of GPR56 by PCR.Then the target gene was cloned into pGEX4T-1 to construct recombinant plasmid pGEX4T-1-GPR56N.After enzyme analysis and sequencing,the vector was transformed into E.coli BL21 and induced by IPTG.Finally,the GPR56 and GST fusion proteinwas purified by GSTbeads to prepare the rabbitantiGPR56 polyclonalantibody.And the specificity of antibody was appraised by Western blotand immunohistochemistry assay.Results:The recombinant protein GST-GPR56N was expressed successfu lly in E.coli.Western blotand immunohistochem istry analysis demonstrated that the polyclonal antibody can recognize the overexp ressed or endogenousGPR56 specifically both in human andmouse tissues.And the titer of anti-GPR56 polyclonalantibody inWestern blotting assaywas about 1∶1 000.Conclusion:The fusion protein GPR56-GST was highly expressed in prokaryotic system.And the antibody was proven to have high titer and good specificity for the study of GPR56.
GPR56;GST;Prokaryotic expression;Polyclonal antibody
R392.33
A
1000-484X(2011)05-0450-05
10.3969/j.issn.1000-484X.2011.05.016
①本文受上海市科委生物醫藥領域重點科技攻關專項(09JC1405200)和中央高校基本科研業務費專項資金資助
趙去非(1985年-),女,生物醫學碩士;
及指導教師:杜 冰(1979年-),男,博士,講師,主要從事免疫學方面的研究,E-mail:bdu@bio.ecnu.edu.cn。
·臨床免疫學·
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
今日農業(2021年19期)2022-01-12 06:16:36
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年11期)2021-08-22 03:15:44
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
學苑創造·A版(2020年9期)2020-10-13 09:41:02
現代出版(2020年3期)2020-06-20 07:10:34
海峽科技與產業(2016年3期)2016-05-17 04:32:12
