香蒲擬發(fā)網(wǎng)菌12S rDNA的PCR擴增及序列分析1)
2011-03-29 08:06:40張旭潘景芝劉福杰王琦李玉
東北林業(yè)大學學報 2011年7期
關(guān)鍵詞:研究
張旭 潘景芝 劉福杰 王琦 李玉
(食藥用菌教育部工程研究中心(吉林農(nóng)業(yè)大學),長春,130118)(長春市傳染病醫(yī)院)(食藥用菌教育部工程研究中心(吉林農(nóng)業(yè)大學))
黏菌(Myxomycetes)在生物界中是介于原生動物與真菌之間的一個特殊類群,其名稱、類群范圍及分類地位,在學者們中的見解始終不一致。雖然現(xiàn)代生物分類學的發(fā)展日益趨于多界系統(tǒng),但黏菌仍然是一個存在爭議的類群[1]。黏菌的分布是世界性的,其生境最常見于林中陰涼濕潤的地方。黏菌在原生質(zhì)團時期生活在潮濕的木頭縫隙和樹皮里面,爬行攝食,形成子實體時移到較干燥的基物表面,如腐朽木段、枯枝落葉、樹皮草莖。本研究在其它黏菌分子生物學研究的基礎(chǔ)上[2-7],將現(xiàn)代分子生物學方法引入香蒲擬發(fā)網(wǎng)菌的系統(tǒng)分類,采用CTAB法對基物培養(yǎng)[8-12]獲得的研究標本進行DNA提取,用White等[13]設(shè)計的引物對其rDNA片段進行擴增和序列測定分析。探討了香蒲擬發(fā)網(wǎng)菌的分子系統(tǒng)學關(guān)系,為其系統(tǒng)演化研究提供理論依據(jù)。
1 材料與方法
香蒲擬發(fā)網(wǎng)菌的基物培養(yǎng):將所采基物放于鋪滿濾紙的搪瓷盤中,用無菌水充分濕潤浸透,加塑料布遮蓋,給予一定散射光,在室溫(15~25℃)下培養(yǎng),觀察黏菌原生質(zhì)團和子實體的發(fā)育過程,至子實體成熟時,收集,陰干后放于標本盒中保存,鏡檢鑒定。
總DNA的提取:①CTAB DNA提取液的配制。CTAB 20 g/L,Tris-HCl(pH值8.0)50 mmol/L,Na2EDTA 10 mmol/L,NaCl 0.7 mmol/L。②TE緩沖液的配制。10 mmol/L Tris,1 mmol/L EDTA(pH值8.0)。③破壁。取少量孢子(0.1 mg)置于1.5 mL離心管中,加入100 μL CTAB提取液,用玻璃研磨杵研磨。④消化。向離心管中再加入300 μL CTAB提取液,輕輕搖動。將管置于65℃水浴中消化1.0~1.5 h,在此過程中輕輕搖動2~3次。⑤抽提:加入等體積預冷的氯仿—異戊醇(V(氯仿)∶V(異戊醇)=24∶1),劇烈振蕩使之形成懸乳狀。12 000 r/min離心10 min,將上清液移至一干凈的離心管中,重復2次。⑥沉淀。加入3 mol/L NaAc(100~130 μL)使溶液終濃度為0.1 mol/L。加入2倍體積預冷的無水乙醇,上下轉(zhuǎn)動混勻,放入-20℃冰箱過夜,沉淀DNA。⑦洗滌。12 000 r/min離心15 min,棄去上清液,用預冷的75%乙醇洗滌DNA沉淀,然后棄去冷乙醇。共洗滌2次。管中乙醇倒凈后,將管口傾斜倒置在濾紙上,使乙醇完全揮發(fā)。⑧溶解。加入30 μL TE緩沖液溶解DNA沉淀,-20℃保存?zhèn)溆谩?/p>
PCR擴增:PCR擴增引物由北京三博遠志生物技術(shù)有限責任公司合成,12S上游引物(5'-AAG GAG CCG GTA TCA AGT A-3'),12S下游引物(5'-TAG AGG GAT GTG AAG TGC C-3')[13]。擴增反應體系為10倍PCR Buffer 5.0 μL,25 mmol/L MgCl25.0 μL,dNTP 5.0 μL,引物各2.0 μL,Taq酶0.6 μL,模板4~6 μL,最后以ddH2O水補足至總體積50 μL。PCR反應條件為94℃變性50 s;48~55℃退火50 s,72℃延伸1 min,共循環(huán)35次,最后再72℃延伸10 min。在上述PCR的反應條件下,將DNA模板原液用TE緩沖液按1∶5,1∶10,1∶15梯度稀釋,改變引物濃度及各項反應條件,從而獲取最佳的DNA擴增條件。
PCR擴增產(chǎn)物的檢測:取PCR反應產(chǎn)物2.5 μL,在經(jīng)溴化乙錠染色的1%的瓊脂糖凝膠上電泳25 min(200 V)后取出,在FR-200紫外與可見電泳分析裝置上觀察,并照相。
PCR產(chǎn)物的序列測定:將PCR原液送至北京三博遠志生物技術(shù)有限責任公司進行序列測定。
序列比對和系統(tǒng)發(fā)育樹的構(gòu)建:采用Clustal X 1.83對NCBI上已注冊的黏菌12S rDNA基因序列片段進行比對,經(jīng)BioEdit version7.0.9.0人工調(diào)整后,運用分析軟件MEGA4中基于缺失位點百分數(shù)的臨位相連法構(gòu)建系統(tǒng)發(fā)育樹,同時進行2000次Bootstrap自舉法檢驗。
2 結(jié)果與分析
2.1 香蒲擬發(fā)網(wǎng)菌子實體及其孢絲、孢子形態(tài)
孢囊群生,有柄,窄圓柱形,頂端稍窄或鈍圓,直立,少數(shù)彎曲,全高3~5 mm,寬0.5~2.0 mm。囊被凋落。柄黑褐色,長為全高的1/3~1/2,囊軸黑色,向上漸細。孢絲稠密,彎曲,分枝并聯(lián)結(jié)。基質(zhì)層膜質(zhì),褐色。孢子成堆時淺紫褐色,球形,有小疣,直徑7~9 μm。根據(jù)《中國真菌志——黏菌卷二》[14]鑒定為香蒲擬發(fā)網(wǎng)菌(圖1)。
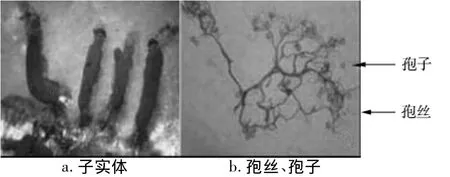
圖1 香蒲擬發(fā)網(wǎng)菌及其孢絲、孢子
2.2 DNA提取
通過CTAB法得到高質(zhì)量的黏菌DNA溶液,可以用于下一步試驗。DNA電泳結(jié)果見圖2。
2.3 PCR擴增
PCR反應適宜條件:所用DNA模板濃度以1∶5稀釋擴增效果最好。最佳的反應體系為10倍PCR Buffer 5.0 μL,25 mmol/L MgCl25.0 μL,dNTP 5.0 μL,引物各2.0 μL,Taq酶0.6 μL,模板4 μL,最后以ddH2O補足至50 μL。
最佳反應溫度與時間:94℃變性50 s,在退火溫度為48℃、時間50 s時出現(xiàn)清晰的擴增條帶(圖3),然后72℃延伸1 min,共35個循環(huán),最后72℃再延伸10 min。
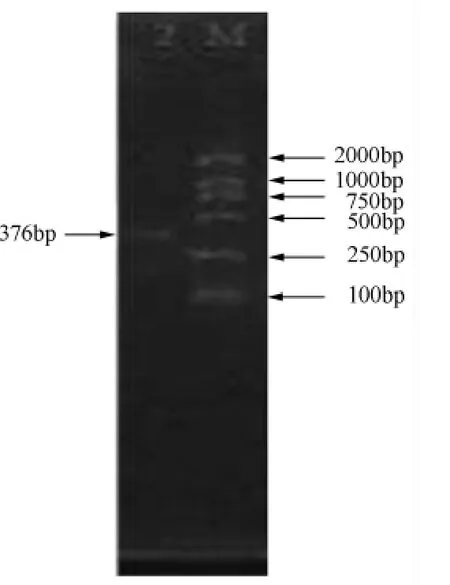
圖3 香蒲擬發(fā)網(wǎng)菌PCR擴增結(jié)果
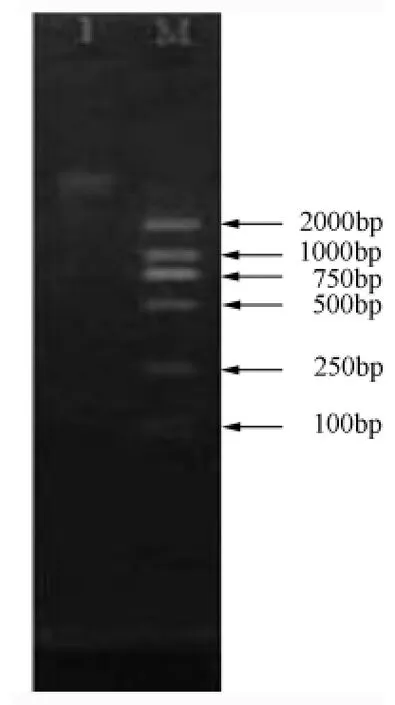
圖2 香蒲擬發(fā)網(wǎng)菌DNA電泳結(jié)果
2.4 測序結(jié)果
香蒲擬發(fā)網(wǎng)菌12S rDNA片斷序列見圖4。該序列全長376 bp,GenBank登錄號為HM102318。

圖4 香蒲擬發(fā)網(wǎng)菌12S rDNA片斷序列
2.5 序列比對和系統(tǒng)發(fā)育樹的構(gòu)建
香蒲擬發(fā)網(wǎng)菌與NCBI上查到的9個黏菌的12S rDNA序列片斷比對結(jié)果見圖5。
粉瘤屬的大粉瘤菌和筒菌屬的筒菌聚在一起,但筒菌的分化早于大粉瘤菌。粉瘤屬的大粉瘤菌,絨泡菌屬的圈絨泡菌、鈣皮菌屬的黑柄鈣皮菌和半網(wǎng)菌屬的棒形半網(wǎng)菌聚集在一起,其Bootstrap支持率為100%,顯示出十分密切的親緣關(guān)系。團網(wǎng)菌屬的灰團網(wǎng)菌與雙皮菌屬的輻射雙皮菌聚在一個分支上。擬發(fā)網(wǎng)菌屬的香蒲擬發(fā)網(wǎng)菌與煤絨菌屬的煤絨菌及半網(wǎng)菌屬蛇形半網(wǎng)菌聚在一個分支上,其支持率分別為66%和46%。
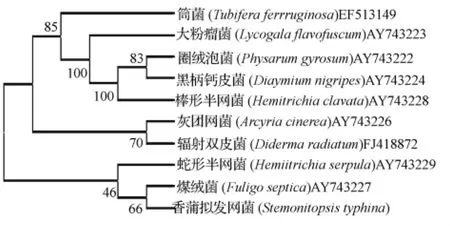
圖5 基于12S rDNA序列分析得到的NCBI已注冊黏菌的系統(tǒng)發(fā)育樹
3 結(jié)論與討論
基物培養(yǎng)是黏菌研究的一種必備手段,通過它可以豐富所考察地區(qū)的黏菌分布,彌補野外采集工作的不足。本試驗所采用的濕室培養(yǎng)方法簡便,無需對器皿和試劑進行消毒滅菌,使得以后的培養(yǎng)工作易于進行。從培養(yǎng)條件上看,變溫條件下濕室培養(yǎng)產(chǎn)生黏菌的頻率較高,溫度恒定或過于極端則不利于黏菌子實體的產(chǎn)生,可能是由于變溫的條件更接近于采集地自然的溫度,有利于黏菌子實體的形成。培養(yǎng)中對光照無苛求,只要保證水份濕度,讓其在自然的變溫和散射光條件下就可生長。
在提取黏菌DNA過程中,最重要因素即為細胞壁破壁過程,適當增加研磨時間可以提高提取DNA的質(zhì)量。相對于玻片壓碎法[3]及勻漿器研磨法[15],應用玻璃研磨杵在離心管中直接研磨黏菌孢子[16],既可以避免勻漿器研磨后轉(zhuǎn)入離心管過程中標本的損失,又可以彌補玻片壓碎法中玻片僅能對黏菌孢子破壁的不足,從而可以更好地利用有限的標本資源,提取高質(zhì)量黏菌DNA。
線粒體DNA是生物體內(nèi)的核外遺傳信息載體,系共價閉環(huán)的環(huán)狀分子,分子量小,基因組中一般沒有間隔序列,結(jié)構(gòu)簡單,為嚴格的母系遺傳,幾乎不發(fā)生倒位、易位等畸變與重組,使之容易被檢測。因而線粒體DNA越來越廣泛地被應用于各類群的系統(tǒng)進化研究。而12S rDNA基因是線粒體DNA上2個rRNA基因之一,在結(jié)構(gòu)上存在4個結(jié)構(gòu)域,第三結(jié)構(gòu)域是相當保守的。該結(jié)構(gòu)域包括32至48號莖,高度保守的側(cè)翼序列使它成為12S rDNA基因中最常被擴增的區(qū)域。同時,12S rDNA基因進化速度較16S、18S、28S rDNA快。因此12S rDNA基因序列適用于研究生物類群的種屬間系統(tǒng)發(fā)生關(guān)系[17]。
目前,GenBank中關(guān)于黏菌12S序列的報道僅有10個種,且關(guān)于香蒲擬發(fā)網(wǎng)菌分子生物學方面的研究尚未見報道。本研究首次成功地對香蒲擬發(fā)網(wǎng)菌的12S rDNA片段進行了PCR擴增。該序列的測得為進一步研究黏菌的系統(tǒng)演化提供了依據(jù)。由于部分黏菌的地域性及其培養(yǎng)條件的不成熟在一定程度上限制了黏菌標本的來源,進而限制了其分子生物學方面的發(fā)展,雖然基物培養(yǎng)在一定程度上解決了這個問題,但其培養(yǎng)條件及基物的選擇還有待進一步研究。
與傳統(tǒng)的形態(tài)學分類不同,在對已知的10個黏菌12S rDNA的序列比對中可以看出團網(wǎng)菌屬、半網(wǎng)菌屬、粉瘤屬、筒菌屬、絨泡菌屬、鈣皮菌屬和雙皮菌屬黏菌的關(guān)系較近,在演化的過程中交替出現(xiàn)。擬發(fā)網(wǎng)菌屬與煤絨菌屬、半網(wǎng)菌屬聚在一個分支上,但是其Bootstrap支持率不高,只能作為參考。黏菌系統(tǒng)發(fā)育的研究需要大量的黏菌基因序列對其分類進行支持,為以后的研究提出了思路。選用不同的分子標記也會造成截然不同的結(jié)論,這種現(xiàn)象在分子系統(tǒng)學研究中并不少見,可能是因不同的DNA標記在進化的各階段所起作用不同,或是統(tǒng)計分析方法中存在的某種缺陷所致,仍有待于作進一步研究。在研究某一特定類群的詳細系統(tǒng)親緣關(guān)系時,單憑某一種或某幾種方法是不夠的,必須綜合多方面資料(形態(tài)、生理、生化、古生物等)進行全面的考查與分析。DNA序列比對結(jié)果做為研究中的證據(jù)之一,還須結(jié)合形態(tài)學等多種手段,相互印證,才可得出符合客觀規(guī)律的結(jié)論,從而為物種的系統(tǒng)發(fā)育、生態(tài)學等研究提供重要的信息。
[1]Kirk P M,Cannon P F,Minter D W,et al.Ainsworth&Bisby's dictionary of the fungi[M].10th ed.U K:CABI Bioscience,CAB International,2008.
[2]劉淑艷,李玉.首次用PCR法擴得長尖團毛菌小亞基rDNA片段[J].吉林農(nóng)業(yè)大學學報,2004,26(3):291-292.
[3]劉淑艷,李玉.首次用一對引物擴得大粉瘤菌小亞基rDNA長片段[J].菌物系統(tǒng),2003,22(1):74-76.
[4]劉淑艷,李玉.黑發(fā)菌核糖體DNA小亞基片段的序列測定[J].菌物研究,2003,1(1):5-8.
[5]白秀娟,李玉,劉淑艷.粘菌基因組DNA提取方法的研究[J].吉林農(nóng)業(yè)大學學報,2003,25(2):158-160.
[6]Kamono A,F(xiàn)ukui M.Rapid PCR-based method for detection and differentiation of Didymiaceae and Physaraceae(myxomycetes)in environmental samples[J].Journal of Microbiological Methods,2006,67(3):496-506.
[7]Kamono A,Matsumoto J,Kojima H,et al.Characterization of myxomycete communities in soil by reverse transcription polymerase chain reaction(RT-PCR)-based method[J].Soil Biology&Biochemistry,2009,41(6):1324-1330.
[8]王琦.中國團毛菌科四個屬的分類研究及雙毛菌屬DITRICHIA的建立[D].沈陽:沈陽農(nóng)業(yè)大學,1994.
[9]王琦,李玉.中國團毛菌目黏菌分支分類研究[J].吉林農(nóng)業(yè)大學學報,2005,27(6):617-620.
[10]周宗璜,張志澄,劉宗麟.從基物培養(yǎng)獲得的幾種粘菌[J].吉林農(nóng)業(yè)大學學報,1981,3(2):1-9.
[11]趙曰鋒.粘菌基物培養(yǎng)的研究[J].吉林農(nóng)業(yè)大學學報,1983,5(2):67-74.
[12]陳雙林,李玉.粘菌濕室培養(yǎng)的初步研究[J].吉林農(nóng)業(yè)大學學報,1995,17(3):33-37.
[13]White T J,Bruns T,Lee S,et al.Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[M]//Innis M A,Gelfand D H,Sninsky J J,et al.PCR Protocols:A guide to methods and applications.San Diego,California:Academic Press,1990:315-322.
[14]李玉,李惠中,王琦,等.中國真菌志:2卷[M].北京:科學出版社,2007:127-128.
[15]劉樸,白秀娟,王琦.筒菌12S rDNA的PCR擴增及序列測定[J].吉林農(nóng)業(yè)大學學報,2007,29(6):633-635.
[16]潘景芝,張旭,劉福杰,等.藍灰團網(wǎng)菌ITS rDNA的PCR擴增及序列測定[J].東北林業(yè)大學學報,2010,38(10):103-105.
[17]鄭冬,劉學東,馬建章.12S rRNA基因及其二級結(jié)構(gòu)在系統(tǒng)學研究中的應用[J].東北林業(yè)大學學報,2003,31(3):59-61.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務(wù)財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
