16個國外鴨茅種質(zhì)材料引種與初步評價
2011-04-25 09:38:46才宏偉劉藝杉王赟文
草業(yè)科學(xué) 2011年4期
徐 倩,才宏偉,劉藝杉,王赟文
(1.中國農(nóng)業(yè)大學(xué)動物科學(xué)技術(shù)學(xué)院草業(yè)科學(xué)系,北京 100193; 2.中國農(nóng)業(yè)大學(xué)農(nóng)業(yè)與生物技術(shù)學(xué)院植物遺傳育種系,北京 100193; 3.北京克勞沃草業(yè)技術(shù)開發(fā)中心,北京100029)
鴨茅(Dactylisglomerata)是一種優(yōu)良的叢生型冷季禾本科鴨茅屬牧草,原產(chǎn)于歐洲、北非和亞洲溫帶地區(qū)[1],現(xiàn)在北美、歐洲、大洋洲和亞洲廣泛種植[2-3]。鴨茅葉量豐富,草質(zhì)柔嫩,牛、羊、馬、兔等均喜食,可用作放牧或制作干草[4],也可收割青飼或制作青貯飼料,具有很好的利用價值。鴨茅抗逆性較強(qiáng),表現(xiàn)出一定的耐旱、耐熱、耐瘠薄和耐蔭能力,是栽培草地建植、天然草地改良與植被恢復(fù)選用的重要草種之一,在我國的云貴高原、四川盆地、華北、西北、東北和華中地區(qū)均有種植,適應(yīng)性廣泛。
按染色體數(shù)目的不同,鴨茅有四倍體和二倍體兩種類型,兩種倍型分別包含多個亞種,相同倍型的不同亞種之間可自由雜交。根據(jù)起源、形態(tài)及生長特征,鴨茅又可分為歐洲型和地中海型,歐洲型的特點(diǎn)是夏季生長,冬季休眠;而地中海型則相反,夏季休眠,冬季生長。目前國外栽培利用的鴨茅品種主要是起源于歐洲的四倍體鴨茅亞種(D.glomeratasubsp.glomerata),二倍體鴨茅亞種栽培利用較少[6]。我國野生鴨茅資源主要分布于西南橫斷山區(qū),有二倍體和四倍體野生類型[7]。在新疆天山山脈的森林邊緣地帶也有分布,并散見于大興安嶺東南坡地[4]。我國野生鴨茅種質(zhì)資源的收集、評價與利用以西南地區(qū)分布的種質(zhì)材料為主,從形態(tài)學(xué)、遺傳學(xué)、農(nóng)藝學(xué)和分子標(biāo)記檢測等方面,開展了大量的遺傳多樣性與栽培品種選育研究[6-15]。除1個國外引進(jìn)品種外,我國選育獲得的鴨茅品種僅有3個,與日本和歐美等發(fā)達(dá)國家相比,還存在很大差距。
為豐富我國鴨茅種質(zhì)資源,積極利用國外選育獲得的品種或種質(zhì)材料,促進(jìn)我國鴨茅新品種選育工作,本研究參考多種牧草的種質(zhì)資源形態(tài)學(xué)研究的方法[16-18],選取16個國外引進(jìn)的野生鴨茅亞種或育成品種材料在北京種植,進(jìn)行初步的田間形態(tài)學(xué)性狀觀測,旨在為鴨茅不同倍性材料的遺傳學(xué)分析與選育利用提供依據(jù)。
1 材料與方法
1.1試驗(yàn)地自然概況 試驗(yàn)于2008-2009年在北京東郊克勞沃草業(yè)技術(shù)開發(fā)中心雙橋科研與生產(chǎn)基地進(jìn)行。試驗(yàn)地地勢平坦、開闊,光照、通風(fēng)條件良好,土壤質(zhì)地為砂質(zhì)土壤,土壤熟化程度高。試驗(yàn)地位于華北平原北端,39°34′ N,116°28′ E,海拔50 m,為典型的暖溫帶半濕潤大陸性季風(fēng)氣候,夏季高溫多雨,冬季寒冷干燥。年平均氣溫10~12℃,年平均降水量為595 mm,其中75%集中在6-8月[19]。
1.2試驗(yàn)材料 參試的16個種質(zhì)材料當(dāng)中,包括9個日本鴨茅育成品種,來自日本草地畜產(chǎn)種子協(xié)會飼料作物研究所,均為首次引入我國的種質(zhì)材料;7個野生鴨茅種質(zhì),來自美國植物引種試驗(yàn)站(表1),其中PI316209和PI538922國內(nèi)已有引進(jìn)與評價研究[12],其余5份材料目前在我國尚未有評價報道。
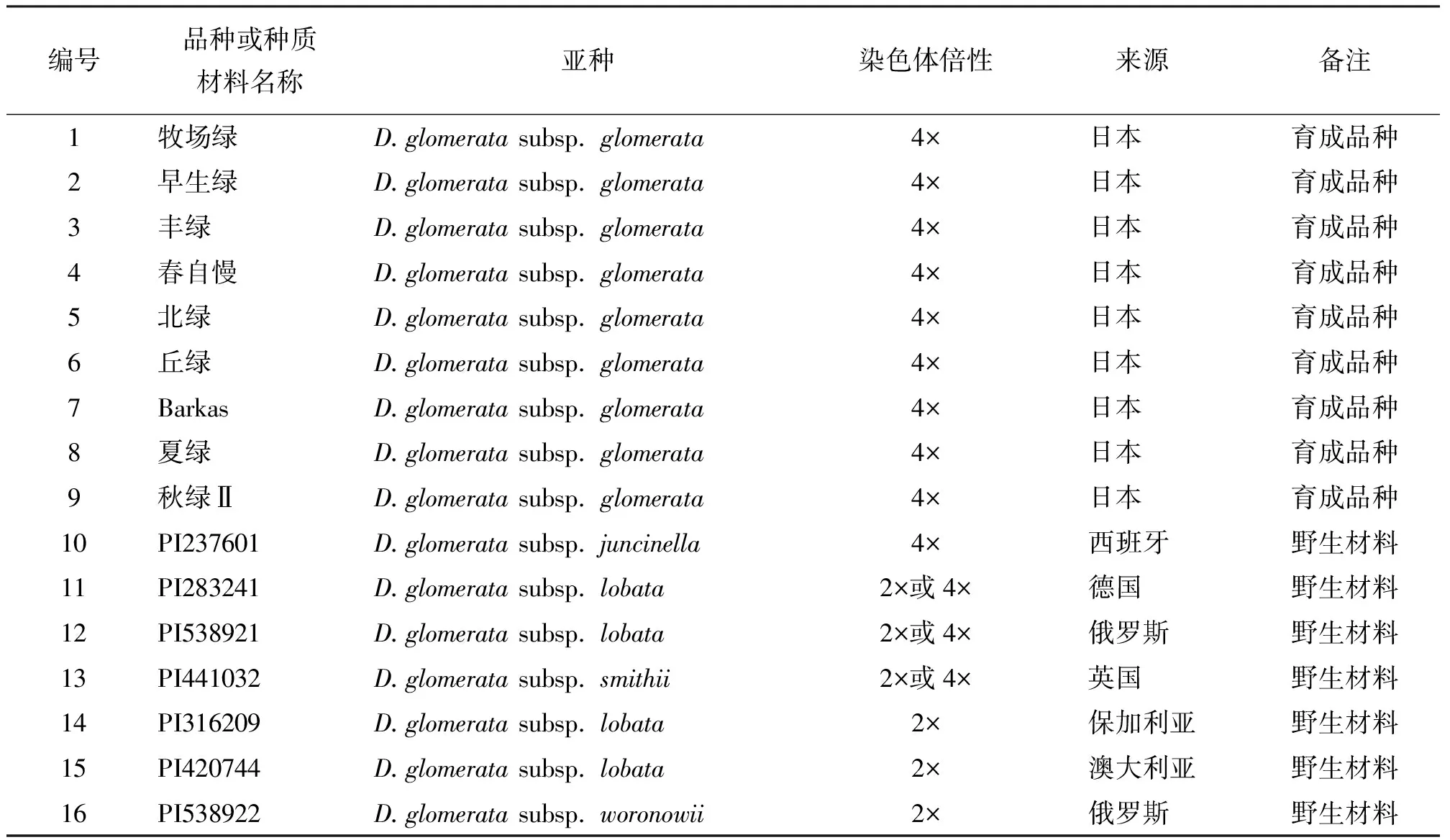
表1 試驗(yàn)材料及其來源
1.3試驗(yàn)設(shè)計 由于各參試材料收集到的種子數(shù)量少,試驗(yàn)按育苗后移栽的方式進(jìn)行,每個材料觀測評價50株,按15株為一個重復(fù),每個參試材料重復(fù)3次。具體試驗(yàn)過程為:2008年7月溫室育苗,當(dāng)年10月將幼苗移至北京東郊的克勞沃草業(yè)技術(shù)開發(fā)中心雙橋基地種植,株行距50 cm×50 cm。
1.4測定項(xiàng)目
1.4.1物候期 對每個品種2009年的返青期、拔節(jié)期、抽穗期、開花期和完熟期分別進(jìn)行觀測記錄,各生育期以50%的植株達(dá)到發(fā)育階段為依據(jù)。
1.4.2形態(tài)學(xué)性狀 參考彭燕等[11]選用的測定指標(biāo)和方法,對每個品種分別進(jìn)行以下性狀觀測:1)抽穗期營養(yǎng)器官性狀:葉長、葉寬、株高、節(jié)間長度、莖粗、旗葉長度、旗葉寬度;2)花序性狀:花序長度、花序?qū)挾龋?)穗部性狀:小穗長度、小穗寬度;4)種子性狀:種子長度、種子寬度、種子千粒重。
1.5數(shù)據(jù)分析 試驗(yàn)數(shù)據(jù)用SPSS 16.0統(tǒng)計軟件,對各參試材料不同觀測性狀的觀測值進(jìn)行單因素方差分析。根據(jù)14個性狀的平均值,在進(jìn)行標(biāo)準(zhǔn)化處理后,對其中15個參試材料進(jìn)行聚類分析。
2 結(jié)果與分析
2.1物候期 參試的16個國外鴨茅材料,2008年10月育苗移栽后,2009年3月開始返青。各參試材料在測試期間,均能安全越冬。除了從西班牙采集的野生鴨茅種質(zhì)PI237601外,其他的15份參試材料在北京地區(qū)均能完成全生育期,而PI237601材料在觀測當(dāng)年未結(jié)種子。參試的9個日本育成品種當(dāng)中,早生綠、春自慢和Barkas成熟期較早,均為97 d,而丘綠成熟期最晚,生育期為104 d(表2)。野生種質(zhì)當(dāng)中,PI283241和PI538921成熟最早,生育期均為93 d,二倍體的3份種質(zhì)材料PI316209、PI420744和PI538922成熟最晚,生育期為106~108 d(表2)。根據(jù)各參試材料已有的染色體倍性信息,初步分析參試的四倍體材料普遍比二倍體材料的返青期早(表2)。
2.2形態(tài)學(xué)性狀 參試的16份鴨茅種質(zhì)材料之間,14項(xiàng)觀測指標(biāo)均不同程度地達(dá)到極顯著(P<0.01)或顯著水平(P<0.05)(表3)。其中,來自西班牙的PI237601材料植株矮小,長勢較其他材料差,不能正常結(jié)實(shí),表現(xiàn)出明顯的不適應(yīng)性。從參試材料在14項(xiàng)觀測性狀上的表現(xiàn)來看,種子千粒重、花序?qū)挾取⑵烊~長度、葉片長度和節(jié)間長度等性狀變異范圍較大,而種子寬度性狀的變異范圍最小,變異系數(shù)為10.1%(表3)。
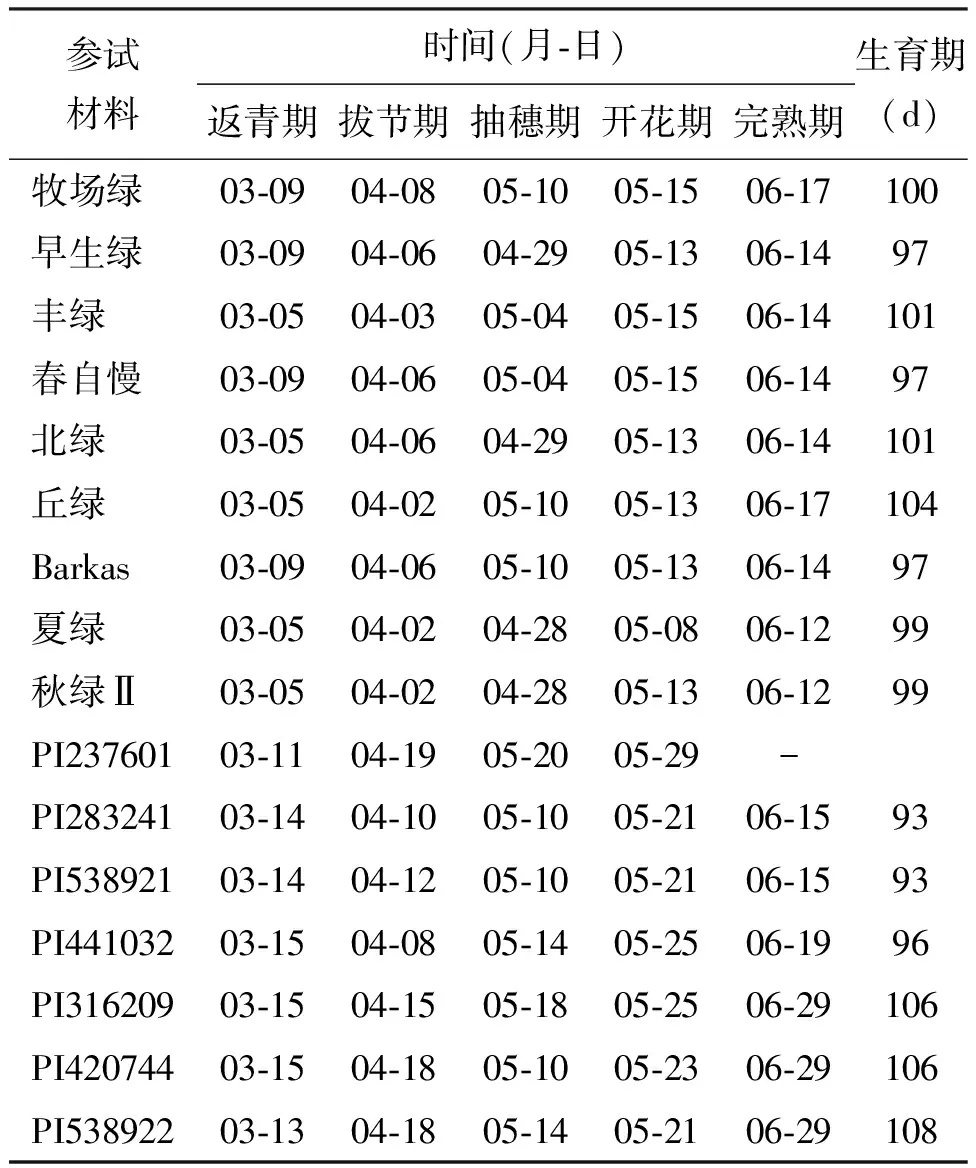
表2 2009年鴨茅參試材料物候期觀測
2.3參試材料形態(tài)學(xué)性狀的聚類分析 根據(jù)以上形態(tài)學(xué)觀測結(jié)果,剔出具有明顯不適性的PI237601材料后,對其余的15個參試材料的14個性狀數(shù)據(jù)進(jìn)行標(biāo)準(zhǔn)化處理,然后進(jìn)行聚類分析,可以初步劃分為四大類(圖1)。
A類:包括‘豐綠’和‘春自慢’2個日本育成品種,以及PI316209、 PI420744和 PI538922共3個野生材料,均為二倍體材料,其中前2個均為鴨茅亞種D.glomeratasubsp.lobata,第3個為鴨茅亞種D.glomeratasubsp.woronowii。
B類:為來自俄羅斯的鴨茅亞種D.glomeratasubsp.lobata野生材料PI538921,主要特點(diǎn)是旗葉較寬。
C類:包括‘Barkas’、‘丘綠’、‘早生綠’、‘北綠’、‘牧場綠’5個日本育成品種和屬于鴨茅亞種D.glomeratasubsp.smithii的PI441032,以及屬于鴨茅亞種D.glomeratasubsp.lobata的PI283241,這類的主要特點(diǎn)是植株較為高大,種子千粒重較高。
D類:包括‘夏綠’、‘秋綠Ⅱ’2個日本育成品種。
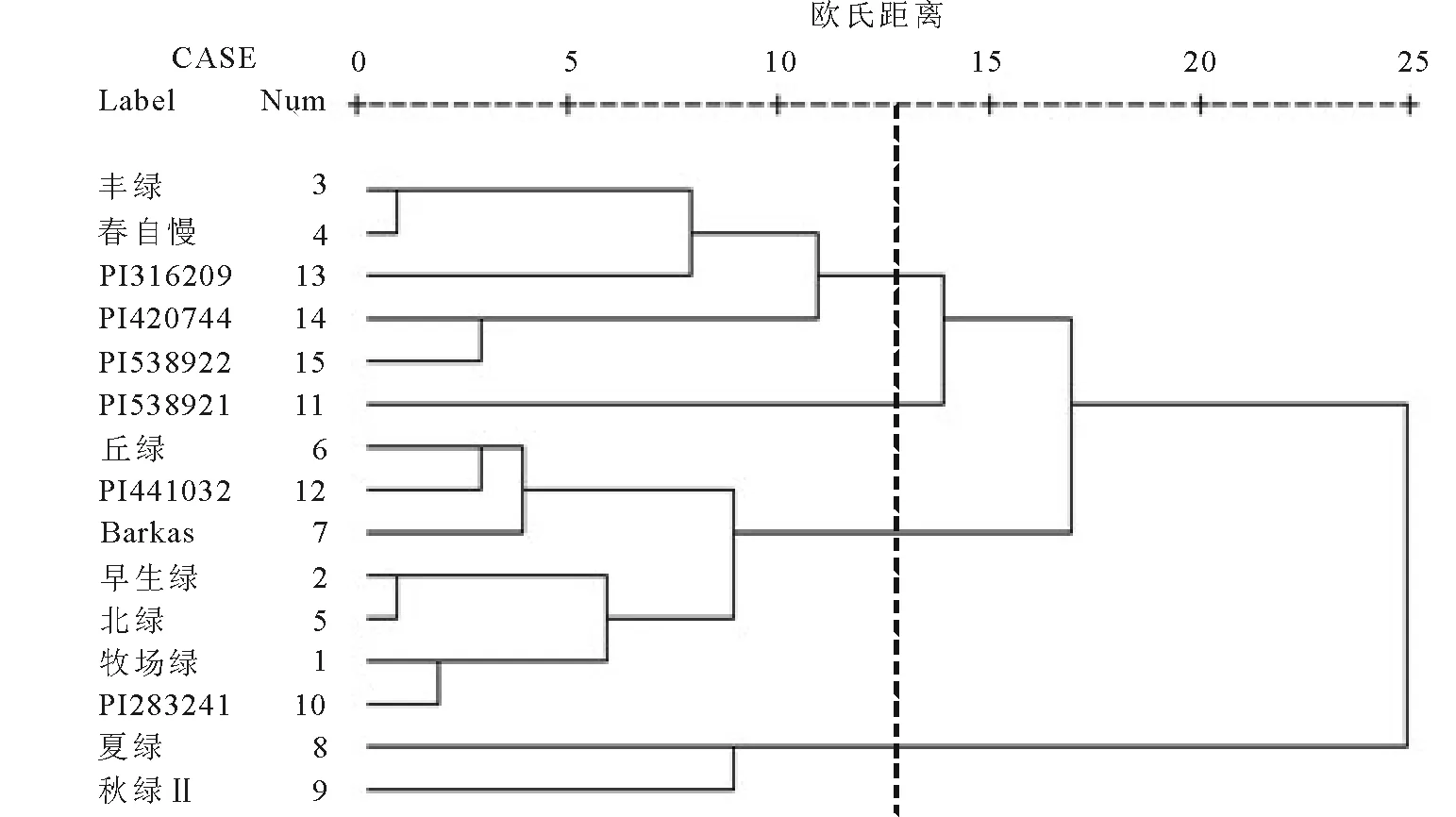
圖1 15個鴨茅表型性狀歐氏距離聚類圖
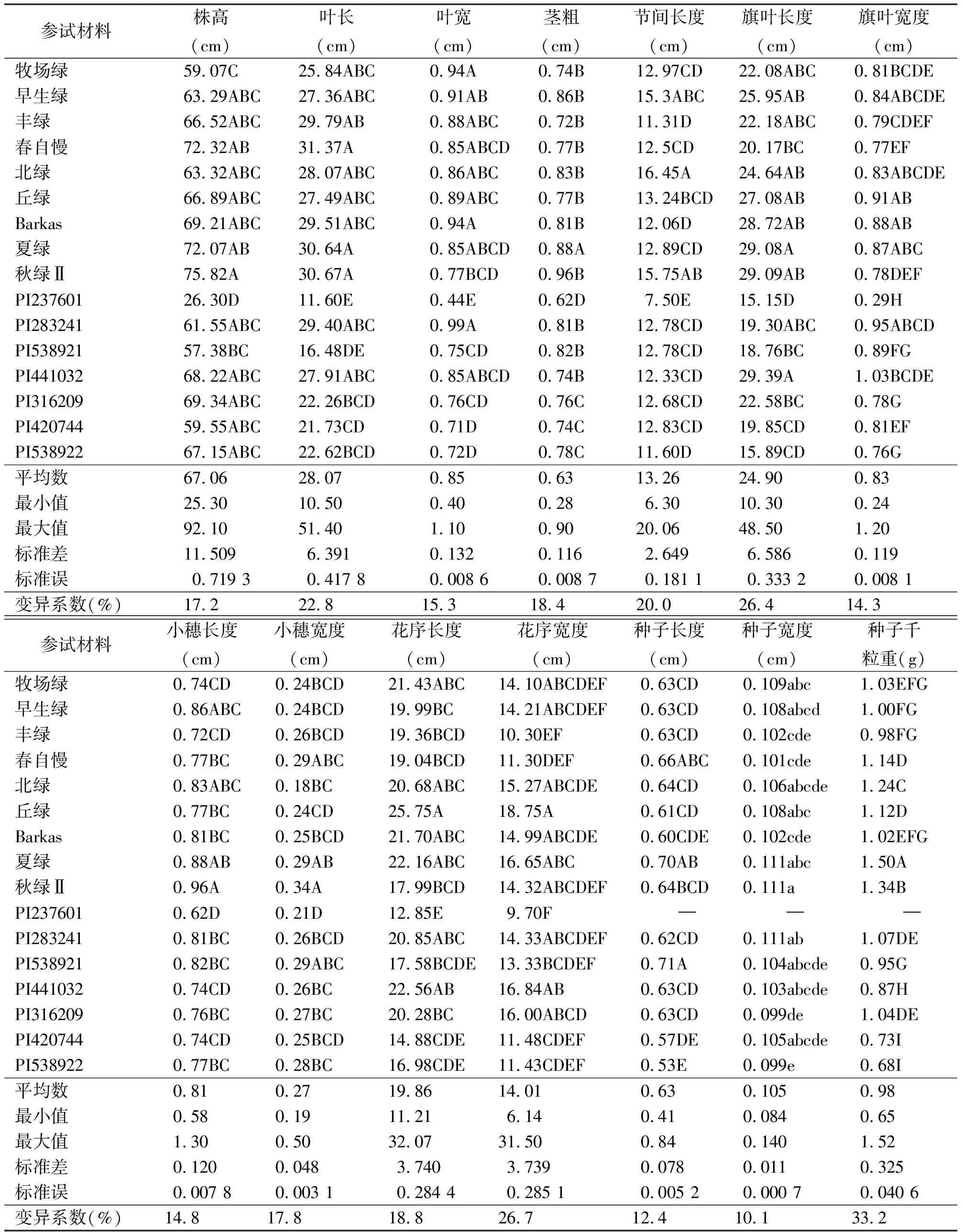
表3 鴨茅主要表型性狀差異分析
3 討論與結(jié)論
3.1物候期 對16份國外鴨茅種質(zhì)材料的引種種植與初步觀測發(fā)現(xiàn),參試的種質(zhì)材料大部分能適應(yīng)北京地區(qū),在移栽后第2年能完成整個生育周期,只有來源于西班牙的野生種PI237601不能結(jié)實(shí)。已知染色體倍性的材料當(dāng)中,四倍體材料的返青期、開花期總體早于二倍體,這與Lumaret和Gulilerm[20]研究的鴨茅四倍體比二倍體開花早的結(jié)論相符合。
3.2表型性狀 通過對這16份材料的表型性狀的觀測分析可以看出,參試的材料形態(tài)學(xué)差異較大。從測試的14項(xiàng)形態(tài)學(xué)指標(biāo)當(dāng)中,種子千粒重、花序?qū)挾取⑵烊~長度、葉片長度和節(jié)間長度等性狀變異范圍較大,變異系數(shù)均大于20%。這與彭燕等[11]對以國內(nèi)野生鴨茅種質(zhì)為主的形態(tài)學(xué)多樣性研究結(jié)果一致。種子長度和種子寬度變異最小,其次是旗葉寬度、小穗長度和葉片寬度變異較小。
3.3聚類分析 參試的16份鴨茅種質(zhì)材料當(dāng)中,除不能完成全生育期的PI237601之外,其余15份材料可劃分為4類,其中3個二倍體鴨茅種質(zhì)材料聚為一類,與其他四倍體材料存在明顯差異。謝文剛等[12]的研究結(jié)果表明,鴨茅的遺傳多樣性與地理來源和氣候類型存在一定的關(guān)系,我國收集的鴨茅野生種質(zhì)與亞洲、歐洲、美洲、大洋洲和非洲等其他地域收集的種質(zhì)之間存在明顯的遺傳差異。本研究與謝文剛等[12]的研究中均包括了PI316209和PI538922兩份種質(zhì)材料,本研究從形態(tài)學(xué)性狀將PI316209和PI538922劃分為一類的結(jié)果與謝文剛等用SSR分子標(biāo)記的分析結(jié)果基本一致。本研究引進(jìn)的鴨茅種質(zhì)對豐富我國鴨茅種質(zhì)資源,為進(jìn)一步開展鴨茅遺傳學(xué)及品種選育具有一定的作用。
[1] Gauthier P,Lumaret R,Bedecarrals A E.Differentiation and coexistence of two paratactic tetraploid subspecies of cocksfoot (Dactylisglomerata) in the Alps[J].New Phyotlogist,1998,139:741-750.
[2] Lumaret R.Cytologygenetics,evolution in the genusDactylis[J].Critical Reviews in Plant Science,1988(7):55-91.
[3] Casler M D,Fales S L,McElroy A R,etal.Genetic progress form 40 years of orchardgrass breeding in North America measured under hay management[J].Crop Science,2000,40:1019-1025.
[4] 潘全山,張新全.禾本科優(yōu)質(zhì)牧草——黑麥草、鴨茅[M].北京:臺海出版社,2000:103.
[5] 王棟.牧草學(xué)各論[M].南京:江蘇科技出版社,1989:76-78.
[6] 鐘聲,黃梅芬,段新慧.中國西南橫斷山區(qū)的野生鴨茅資源[J].植物遺傳資源學(xué)報,2010,11(1):1-4.
[7] 彭燕,張新全.鴨茅種質(zhì)資源多樣性研究進(jìn)展[J].植物遺傳資源學(xué)報,2003,4(2):179-183.
[8] 張新全,杜逸,鄭德成.鴨茅二倍體和四倍體生物學(xué)特性的研究[J].四川農(nóng)業(yè)大學(xué)學(xué)報,1996,14(2):202-206.
[9] 鐘聲,杜逸,鄭德成,等.二倍體鴨茅農(nóng)藝性狀的初步研究[J].草地學(xué)報,1997(1):54-61.
[10] 鐘聲,杜逸,鄭德成,等.野生四倍體鴨茅農(nóng)藝性狀的初步研究[J].草業(yè)科學(xué),1998,15(2):20-23.
[11] 彭燕,張新權(quán),曾兵.野生鴨茅植物學(xué)形態(tài)特征變異研究[J].草業(yè)學(xué)報,2007,16(2):69-75.
[12] 謝文剛,張新全,馬嘯,等.鴨茅種質(zhì)遺傳變異及親緣關(guān)系的SSR分析[J].遺傳,2009,31(6):654-662.
[13] 曾兵,張新全,楊武云,等.SSR在鴨茅種質(zhì)資源遺傳多樣性研究中的應(yīng)用初探[J].西南農(nóng)業(yè)學(xué)報,2006,19(6):1028-1033.
[14] 謝文剛,張新全,彭燕,等.鴨茅SSR-PCR反應(yīng)體系優(yōu)化及引物篩選[J].分子植物育種,2008,6(2):381-386.
[15] 張新全,杜逸,鄭德成,等.鴨茅染色體核型分析[J].中國草地,1994(3):55-57.
[16] 李培英,孫宗玖,阿不來提.偃麥草種質(zhì)資源外部性狀變異及其形態(tài)類型的初步研究[J].草業(yè)科學(xué),2010,27(1):71-78.
[17] 劉江,陳興福,楊文鈺,等.四川盆地麥冬的種質(zhì)資源的形態(tài)特征變異分析[J].草業(yè)學(xué)報,2010,19(1):143-150.
[18] 姚慶群,白昌軍,王文強(qiáng),等.22份禾本科牧草種質(zhì)適應(yīng)性評價[J].草業(yè)科學(xué),2009,26(6):84-89.
[19] 陳鵬,蘇德榮.地下滴灌對草坪土壤水分及根系分布的影響[J].灌溉排水學(xué)報,2008,27(6):48-50.
[20] Lumaret R,Gulilerm J L.Polyploidy and habitat differentiation inDactylisglomerataL. from Galicia [J].Oecologia,1987,73(3):436-446.
