鹽脅迫對雷竹生理生化特性的影響
2011-05-09 05:41:33何奇江周文偉傅懋毅
浙江林業科技 2011年1期
關鍵詞:植物
何奇江,李 楠,王 波,周文偉,傅懋毅
(1. 中國林業科學研究院,北京 100091;2. 浙江省林業科學研究院,浙江 杭州 310023)
鹽脅迫對雷竹生理生化特性的影響
何奇江1,2,李 楠2,王 波2,周文偉2,傅懋毅1
(1. 中國林業科學研究院,北京 100091;2. 浙江省林業科學研究院,浙江 杭州 310023)
應用盆栽法,研究了在不同NaCl濃度(0、0.1%、0.3%、0.5%)梯度脅迫下雷竹葉片內的超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性和丙二醛(MDA)含量的變化。結果表明:①不同鹽脅迫下雷竹都保持了較高的SOD活性成分,且在脅迫后期SOD活性成分高于對照或與對照持平;②高鹽脅迫后期葉片中POD活性成分和對照相比略有下降,但在初期持平,甚至在中期顯著高于對照,中鹽脅迫下的葉片中POD活性成分在中期高于對照,后期和對照持平,而低鹽脅迫和對照相比變化不是很明顯;③鹽脅迫下MDA的含量與對照差異不顯著,其中在后期低鹽脅迫下MDA的含量較對照低;④不同處理條件下,各項生理生化指標間的相關性不顯著。綜合來看,中鹽脅迫下SOD和POD活性均保持較高水平,能抑制膜脂質過氧化作用,并使MDA含量保持在較低水平,表明雷竹能在短期內忍受Na+濃度在0.3%的脅迫。
雷竹;鹽脅迫;抗氧化酶活性;丙二醛(MDA)
雷竹(Phyllostachys praecoxcv.prevevnalis)是浙江西北地區的主要筍用竹種之一,因其出筍早、產量高、筍味好而受到廣泛栽培。近年來通過覆蓋早出豐產技術,取得了很大的經濟效益,目前已成為竹區農民提高經濟收入的重要手段,并通過引種廣泛栽培到其它地區。近幾年在沿海地區也有所發展,但沿海有些地區種植地是近海圍墾地,含鹽量較高,致使個別引種地出現雷竹死亡現象,而有些地方雖然成活,但生長也嚴重不良。
植物在長期進化過程中,為保護自身膜系統免受傷害而形成了一整套相應的抗氧化保護系統。鹽分條件下,膜系統的變化分成兩個階段:首先,表現為鹽分對膜系統的破壞,也反映其對鹽分的忍耐程度;然后是植物對膜系統的修復。膜系統的修復與SOD、POD和CAT等酶活性的升高是分不開的[1]。SOD的主要功能是清除自由基、控制脂質過氧化和減少細胞膜系統的傷害,其活性變化直接影響植物體內O2與H2O2的含量,植物組織中SOD活性的高低以及在逆境中的變化在一定程度上可以反映植物抗逆性的強弱;POD也是抗氧化系統中的重要一員,主要清除H2O2形成體內活性氧清除鏈的重要一環;CAT能夠在逆境脅迫或衰老過程中清除植物體內的過氧化氫,保護細胞膜結構,減輕有毒物質對活細胞的毒害,延遲或阻止細胞結構的破壞,從而使植物能在一定程度上忍耐、減緩或抵抗逆境的脅迫,其CAT活性高低與植物的抗逆性密切相關[2]。三種酶能夠相互協調,清除鹽脅迫下膜脂過氧化作用產生的MDA,保護膜結構的穩定。王愛國等[3]研究證實,MDA含量多少可代表膜損傷程度的大小。因此,植物體內的活性氧清除系統活性或含量的高低是反映植物抗性的重要標志之一。鹽分脅迫對雷竹生長和代謝的影響是多方面,目前對雷竹進行鹽分脅迫處理尚未見報道。本文研究了在不同鹽分脅迫程度下,雷竹超氧化物歧化酶(SOD)活性、過氧化物酶(POD)活性和丙二醛(MDA)含量的變化與鹽分脅迫的關系,為雷竹的耐鹽機理提供一定的理論依據。
1 材料和方法
1.1 試驗材料與處理
盆栽試驗在浙江省林業科學研究院竹類植物園內進行,試驗的雷竹于2009年3月盆栽到內徑約60 cm,深50 cm的陶瓷盆里,盆內土壤取自浙江省林業科學研究院竹類植物園雷竹林內,雷竹年齡為1年生竹苗(2008年的雷竹),地徑基本一致,為3 cm左右,共栽培40盆,并于7 d后對死亡的雷竹進行重新種植,種植后進行日常管理。2009年10月30號對盆栽竹子用不同濃度NaCl溶液進行澆灌,共設4種處理:濃度梯度分別為0(ck)、0.1%、0.3%、0.5%,每種處理10盆。每2 d澆1次,每次澆灌至飽和,并在處理的第6、第12、第18、第24、第30天即11月4、10、16、22、28日取樣,采用混合采樣法采集植物功能葉片,每次每種處理取3盆,低溫處理下帶回實驗室進行各項指標測定,每項指標重復測定3次。
1.2 測定方法
SOD活性測定采用氮藍四唑(NBT)光還原法,以抑制氯化硝基氮藍四唑(NBT)光化還原50%為一個酶活性單位;POD活性測定采用愈創木酚法,以每分鐘內A470變化0.01為1個過氧化物酶活性單位;MDA含量測定采用TBA法[4]。
1.3 數據統計分析方法
用Excel軟件進行數據統計,用SPSS16.0統計軟件
進行方差分析、多重比較和相關分析。
2 結果與分析
2.1 鹽分脅迫對SOD活性的影響

圖1 NaCl脅迫對竹葉SOD活性的影響Figure 1 Effect of NaCl on SOD activities in leaves ofPh. Praecoxcv.prevernalis
SOD是活性氧清除反應過程中第一個發揮作用的抗氧化酶,SOD對于清除氧自由基,防止氧自由基破壞細胞的組成、結構和功能,保護細胞免受氧化損傷具有十分重要的作用[5]。它能將超氧化物陰離子自由基快速歧化為過氧化氫(H2O2)和分子氧。圖1顯示,4種處理的雷竹葉片中SOD活性成分都有一定的波動。經方差分析和多重比較,對照變化較為平緩;低鹽和中鹽處理的SOD活性成分在脅迫前期下降較快,中期的下降趨勢有所緩解,到脅迫后期低鹽處理中的SOD活性成分趨于穩定,而中鹽中的活性成分則開始下降;高鹽處理SOD活性成分呈上升趨勢。與對照相比(如表1所示),鹽脅迫初期低鹽和中鹽處理SOD活性成分比對照高,說明雷竹在鹽處理后葉片中的SOD活性成分就迅速上升,此時高鹽處理和對照的差異不明顯;鹽脅迫的第12天,高鹽處理的SOD活性成分迅速升高,且顯著高于對照,低鹽處理也較對照高,且差異明顯;在脅迫的第18天,中鹽和高鹽處理的SOD活性成分和對照無顯著差異,低鹽處理的SOD活性成分略有下降,且達到顯著差異;在脅迫的第24天,各種鹽處理的SOD活性成分均保持了較高水平,并顯著高于對照;在鹽脅迫的后期,低鹽和高鹽處理SOD活性成分最高,顯著高于對照,中鹽處理的SOD活性成分和對照相似。
總體來看,不同鹽脅迫下雷竹都保持了較高的SOD活性成分,且在脅迫后期SOD活性成分高于對照或與對照持平。說明在整個鹽脅迫過程中,SOD活性隨著鹽脅迫時間的延長和加重呈升高的趨勢。
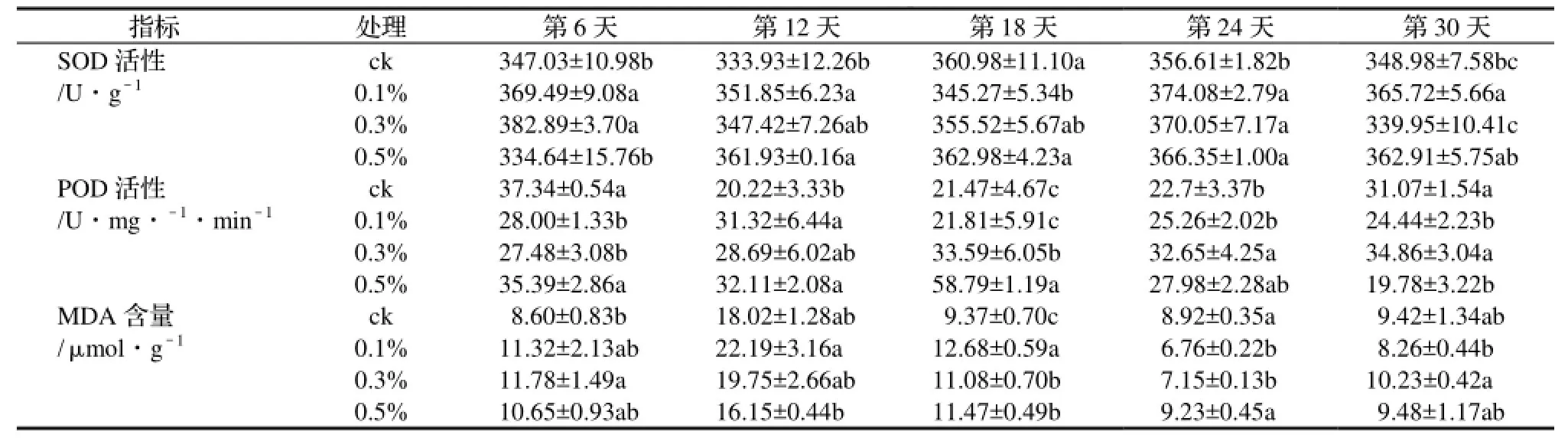
表1 不同鹽濃度處理對竹葉各項生理生化指標的影響Table 1 Effect of different NaCl treatments on the physiochemical index in leaf
2.2 鹽分脅迫對POD活性的影響
SOD清除超氧陰離子后,又會產生氧化能力極強的過氧化氫、羥自由基和單線態分子氧,這些物質也對細胞造成很大的傷害。POD是植物體內擔負清除H2O2的主要酶類之一,能夠催化H2O2氧化產生H2O[6],從而降低膜內不飽和脂肪酸的過氧化程度,維持了細胞膜的穩定性和完整性,使植物耐鹽性進一步提高。由圖2可以看出,4種處理的雷竹葉片中POD活性成分也有一定的波動。經方差分析和多重比較,中鹽處理中POD活性變化較為平緩;對照和低鹽處理中POD活性變化趨勢相似,在初期下降較快,后期又有所回升;高鹽處理中POD活性,表現為先上升并在第18天達到最大值,而后開始迅速下降,并低于對照。與對照相比(如表1所示),在鹽脅迫初期低鹽和中鹽處理POD活性成分比對照低,高鹽處理與對照沒有差異;在第12天各種鹽脅迫處理中POD活性成分均較對照高,其中低鹽和高鹽處理的POD活性成分顯著高于對照;第18天到第22天,中鹽和高鹽脅迫處理中POD活性成分均較對照高,低鹽和對照沒有顯著差異;到脅迫后期,中鹽處理和對照POD活性成分沒有顯著差異,且均比低鹽和高鹽處理高。

圖2 NaCl脅迫對雷竹葉片保護酶POD活性的影響Figure 2 Effect of NaCl on POD activities in leaves ofPh. praecoxcv.prevernalis
總體來說,高鹽脅迫后期葉片中POD活性成分和對照相比略有下降,但在初期持平,甚至在中期顯著高于對照;中鹽脅迫下的葉片中POD活性成分在中期高于對照,后期和對照持平;而低鹽脅迫和對照相比變化不是很明顯。
2.3 鹽分脅迫對MDA含量的影響
鹽分能增加細胞膜透性,加強脂質過氧化作用,最終導致系統的破壞。丙二醛(MDA)是膜脂過氧化的最終產物之一,會嚴重損傷生物膜,其含量的高低反映了細胞膜脂過氧化水平[7]。由圖3所示,4種處理的雷竹葉片中MDA的含量的變化趨勢相似,均呈現出先上升后下降的趨勢,并在后期趨于穩定。經方差分析和多重比較,在初期各種處理MDA的含量均迅速上升,在第12天又迅速下降,到后期變化趨于平緩。與對照相比(如表1所示),在鹽脅迫初期中鹽處理的MDA的含量顯著高于對照,低鹽和高鹽處理與對照無顯著差異;第12天各種處理的MDA的含量均達到最大,其中低鹽處理的MDA的含量最大,較對照和中鹽處理無顯著差異,并顯著高于高鹽處理,第18天各鹽處理的MDA的含量均顯著高于對照,說明鹽脅迫引發了雷竹的膜脂過氧化作用,并隨著鹽脅迫時間的延長,其MDA的含量也呈升高的趨勢;第24天高鹽處理的MDA的含量和對照相似,低鹽和中鹽處理低于對照水平;到后期,各種鹽脅迫下的MDA的含量與對照持平,低鹽處理的MDA的含量最低。
總的來說在各個時期,鹽脅迫下MDA的含量與對照差異不顯著,其中在后期低鹽脅迫下MDA的含量較對照低。雷竹在鹽脅迫下MDA含量雖然在前期迅速增加,但和對照沒有達到顯著差異,且沒有隨著鹽脅迫時間的延長發生明顯的上升趨勢,說明盡管雷竹體內可能存在鹽脅迫而誘導自由基的傷害,但能夠通過一定的自我調控機制來降低鹽分脅迫所引發的膜脂過氧化作用。

圖3 NaCl脅迫對雷竹葉片MDA含量的影響Figure 3 Effect of NaCl on MDA content in leaves ofPh. praecoxcv.prevernalis
2.4 鹽分脅迫下雷竹葉片內SOD活性、POD活性與MDA含量關系
鹽脅迫對雷竹葉片內的SOD、POD活性和MDA含量均產生了一定的影響,三者間的關系如表2。
由表2可知,不同處理條件下,各項生理生化指標間的相關性不顯著。其中對照、低鹽和中鹽處理下的SOD活性和MDA含量呈負相關,高鹽脅迫下SOD活性和MDA含量呈正相關,這表明較高的SOD活性能夠抑制膜脂質過氧化作用,使MDA含量保持在較低水平。而隨著脅迫的加深,氧自由基的積累,使膜系統的脂質過氧化作用增強,MDA含量也隨之升高。POD活性和MDA含量在對照和中鹽脅迫下也呈負相關,低鹽和高鹽處理下呈正相關。SOD活性與POD活性之間,除了低鹽脅迫下呈負相關外,其余處理下均呈負相關。
3 結論與討論

表2 不同鹽濃度處理下竹葉各項生理生化指標間的相關關系Table 2 Correlations of physiochemical indexes among different treatments
(1)活性氧清除系統對植株抗鹽生理有十分重要的作用,其中SOD是抗氧化系統中一種極為重要和在生物體內普遍存在的金屬酶。在酶促保護系統中,SOD處于核心地位,是活性氧清除反應過程中第一個發揮作用的抗氧化酶。在本實驗中,SOD首先對鹽脅迫所造成的膜傷害作出反應,可以看到雷竹在鹽脅迫初期低鹽和中鹽處理的SOD活性成分就迅速上升,且明顯高于對照。不同鹽脅迫條件下雷竹都保持了較高的SOD活性成分,表明雷竹在抗鹽機理方面對鹽脅迫的適應性。隨著鹽脅迫時間的延長和加重,酶促防御系統機能有所降低,高鹽脅迫下雷竹葉片中POD活性成分要明顯低于對照,但在初期持平,甚至在中期顯著高于對照;中鹽脅迫下的葉片中POD活性成分在中期高于對照,后期和對照持平;而低鹽脅迫和對照相比變化不是很明顯。何開躍等[8]研究表明,隨著鹽脅迫濃度的加重,白竹(Phyllostachys dulcis)筍體內的SOD、POD活性先升高后降低,紅竹(Phyllostachy iridenscens)則為先降低后升高,而翠竹(Arundinaria pygmaea)葉片酶活性則變化平緩。
(2)隨著鹽脅迫時間的延長,各種處理下MDA的含量與對照差異不顯著,其中低鹽脅迫下MDA的含量在后期要低于對照。雖然在鹽脅迫的前期MDA含量迅速增加,但沒有和對照達到顯著差異,也沒有隨著鹽脅迫時間的延長發生明顯的上升趨勢,說明雷竹能夠通過一定的自我調控機制來降低鹽分脅迫所引發的膜脂過氧化作用。這也表明雷竹在一定的鹽濃度范圍及時間內能夠通過自身誘導抗氧化酶系統以提高適應能力而忍受逆境。有研究表明不同濃度NaCl脅迫下,隨著時間的增加,菲白竹(Pleioblastus fortune)、鋪地竹(Pleioblastus argenteostriatus)葉片的MDA含量呈上升趨勢[9],紅竹和早竹(Phyllostachys praecox)呈先降后升趨勢,而白哺雞竹的MDA相對含量變化不大[10],可見各竹種間的耐鹽能力不同。
(3)相關分析表明,各項生理生化指標間的相關性不顯著。只有高鹽處理下SOD活性和MDA含量呈正相關,說明隨著脅迫的加深,氧自由基的積累,使膜系統的脂質過氧化作用增強,MDA含量也隨之升高。POD活性和MDA含量在對照和中鹽脅迫下也呈負相關。SOD活性與POD活性之間,除了低鹽脅迫下呈負相關外,其余處理下均呈負相關。
(4)綜合來看,中鹽脅迫下SOD活性和POD活性均保持了較高的水平,能夠抑制膜脂質過氧化作用,并使MDA含量保持在較低水平。表明雷竹能在短期內忍受Na+濃度在0.3%的脅迫。
[1] 于秀俊,張兆英. 植物抗鹽性評價生理指標的分析[J]. 滄州師范專科學校學報,2006,22(4):51-53.
[2] 劉金文,佟丹丹,關刺婭. NaCl脅迫對不同苜蓿品種超氧化物歧化酶同工酶的影響[J]. 黑龍江八一農墾大學學報,2006(6):4-7.
[3] 王愛國. 丙二醛作為脂質過氧化指標的探討[J]. 植物生理學通訊,1986(2):55-57.
[4] 鄭柄松. 現代植物生理生化研究技術[M]. 北京:氣象出版社,2006.
[5] 馬旭俊,朱大海. 植物超氧化物歧化酶(SOD)的研究進展[J]. 遺傳,2003,25(2):225-231.
[6] Lu Q,Zheng R. M embranes lipid peroxi-dation change and lipidremoval deduced by drought and activated oxygen[J]. Sci China (SeriC),1996(26):26-29.
[7] 汪貴斌,曹福亮. 土壤鹽分和水分脅迫對落羽杉葉片中幾種酶活性的影響[J]. 南京林業大學學報(自然科學版),2006,30(6):32-36.
[8] 何開躍,郭春梅. 鹽脅迫對3種竹子體內 SOD, POD活性的影響[J]. 江蘇林業科技,1995,22(4):11-14.
[9] 萬賢崇,宋永俊. 鹽脅迫及其鈣調節對竹子根系活力和丙二醛含量的影響[J]. 南京農業大學學報,1995,19(3):16-20.
[10] 李善春. NaCl鹽脅迫下5種地被觀賞竹生理特性的研究[D]. 南京:南京林業大學,2005.
Effect of NaCl Treatment on Phisiochem ica l Properties o fPhyllostachys praecoxcv.prevernalis
HE Qi-jiang1,2,LI Nan2,WANG Bo,ZHOU W en-wei2,FU Mao-yi1
(1. Chinese Academy of Forestry, Beijing 100091, China; 2. Zhejiang Forestry Academy, Hangzhou 310023, China)
Determ ination was conducted on SOD and POD activity and MDA content in leaf of pottedPhyllostachys praecoxcv.prevernalisunder different NaCl treatments(0, 0.1%, 0.3%,0.5%). The result showed that different NaC l treatedPh. praecoxcv.prevernaliscould hold high SOD activity, and even higher than the control at the later period. POD activity in leaf under high stress decreased a little than that of the control at the later stage, but similar at earlier stage and even higher evidently at middle state. POD activity in leaf under medium stress was higher and similar that that of the control at middle and later stage. POD under low salt stress had no evident difference of both treated bamboo and the control. MDA content in leaf of treated bamboo had no great difference w ith that in the control. It had no close relationship among different physiochemical properties under different salt stress. The result demonstrated that medium salt stress treated bamboo could hold high SOD and POD activity and low MDA content, indicatingPh. praecoxcv.prevernalishad salt tolerance under 0.3% of Na+ during short time.
Ph. praecoxcv.prevernalis; salt stress; SOD; POD; MDA
S718.43
A
1001-3776(2011)01-0044-05
2010-08-11;
2010-12-13
“十一五”國家科技支撐計劃專題“亞熱帶基巖質海岸防護林體系構建技術試驗示范”(2006BAD 03A 1406)
何奇江(1976-),男,浙江諸暨人,副研究員,從事竹林培育研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
