Bt殺蟲基因?qū)@Wo現(xiàn)狀與趨勢
2011-06-12 00:58:44束長龍張春鴿
植物保護 2011年3期
關(guān)鍵詞:研究
張 杰, 束長龍, 張春鴿
(中國農(nóng)業(yè)科學院植物保護研究所,植物病蟲害生物學國家重點實驗室,北京 100193)
在1996-2010年的15年間,全球轉(zhuǎn)基因植物種植面積累計首次超過10億hm2。2010年全球轉(zhuǎn)基因作物種植面積達到1.48億hm2,占全球耕地總面積的10%,比1996年增長了87倍,與之相對應(yīng)的是,2010年轉(zhuǎn)基因作物種子的全球市場價值為112億美元,轉(zhuǎn)基因玉米、大豆以及棉花產(chǎn)品創(chuàng)造價值約為1500億美元,這些轉(zhuǎn)基因作物中約39%是具有抗蟲性狀的,主要是轉(zhuǎn)入了Bt殺蟲基因[1]。面對Bt殺蟲基因的巨大市場前景,近年來,西方發(fā)達國家及其農(nóng)業(yè)生物技術(shù)跨國公司不斷加大投入,進行新型Bt殺蟲基因發(fā)掘和相關(guān)技術(shù)開發(fā),并利用知識產(chǎn)權(quán)將這些基因和技術(shù)轉(zhuǎn)化為自己獨占的權(quán)利,以期在未來的競爭中掌控主動權(quán)[2-4]。我國于20世紀90年代后期開始了相關(guān)的研究工作,與西方發(fā)達國家由企業(yè)主導(dǎo)的研發(fā)不同,我國Bt殺蟲基因發(fā)掘的主體是國家科研院所與大專院校,目前在國家“863”計劃、“轉(zhuǎn)基因生物產(chǎn)業(yè)化重大專項”等項目的資助下,呈現(xiàn)出加速發(fā)展的良好勢態(tài)。
本文主要分析Bt殺蟲蛋白基因(cry基因)的克隆命名情況,并對目前受專利保護的基因分布狀況進行統(tǒng)計,借此為我國相關(guān)研究人員在轉(zhuǎn)基因技術(shù)研發(fā)、產(chǎn)業(yè)化應(yīng)用過程中,合理有效地規(guī)避知識產(chǎn)權(quán)陷阱,有效利用Bt資源提供決策參考。
1 Bt殺蟲蛋白分類
Bt是蘇云金芽胞桿菌(Bacillusthuringiensis)的簡稱,在1901年作為一種昆蟲病原菌被發(fā)現(xiàn),從1940年起被大規(guī)模開發(fā)成微生物殺蟲劑,用于害蟲防治。Bt特異的殺蟲作用一直受到人們的關(guān)注,其殺蟲物質(zhì)主要是殺蟲晶體蛋白(insecticidalcrystal proteins,簡稱ICPs),由cry基因和cyt基因編碼;隨著研究的深入,從Bt中不斷分離到新的殺蟲活性蛋白,例如營養(yǎng)期殺蟲蛋白(vegetativeinsecticidal protein,簡稱Vips)等[5-7]。
ICPs是Bt主要的毒性物質(zhì),具有特異的殺蟲活性。1981年,Schnepf首次克隆到cry基因,并證實了其殺蟲活性[8]。截止到目前,全球已經(jīng)克隆并被Bt殺蟲毒素國際命名委員會命名的殺蟲晶體蛋白基因有 605個,按照命名規(guī)則分為72個大類(cry1~cry68,cyt1~cyt4),包括230種模式基因。目前Bt殺蟲毒素使用的命名規(guī)則是1998年Neil Crickmore博士提出的,殺蟲毒素蛋白按照其編碼的氨基酸序列相似性進行命名。按照目前的命名規(guī)則,所有毒素都可以獲得一個包含4個等級的命名(圖1)。如果現(xiàn)在有一種新獲得的毒素,它與目前所有毒素氨基酸序列相似性都小于45%,屬于第1等級的新蛋白,它則會被命名為 Cry69Aa1;與Cry68Aa1的一致性在45%~78%之間則會被命名為Cry68Ba1,屬于第2等級的新蛋白;與Cry68Aa1的一致性在 78%~95%之間則會被命名為Cry68Ab1,屬于第3等級的新蛋白;而與Cry68Aa1的一致性大于95%則會被命名為Cry68Aa2,屬于第4等級的蛋白,以此類推[9]。上述前3類的新毒素與目前已命名毒素序列一致性小于95%,與之相對應(yīng)的基因通常稱為模式基因(holo-typegene),若其殺蟲活性高,即可直接申請發(fā)明專利。
這些已發(fā)現(xiàn)的Cry毒素對無脊椎動物中4個門和節(jié)肢動物門中9個目的500余種有害生物具有殺蟲活性,并且這些基因擁有各自獨特的殺蟲譜,如Cry1、Cry3、Cry4、Cry5可分別毒殺鱗翅目、鞘翅目、雙翅目等有害昆蟲,以及線蟲、肝吸蟲等;Cry2對鱗翅目、雙翅目昆蟲有活性;Cry1B、Cry1I對鞘翅目、鱗翅目有活性[10]。目前已有十余種cry基因應(yīng)用于玉米、棉花、馬鈴薯和番茄等抗蟲作物構(gòu)建[4]。
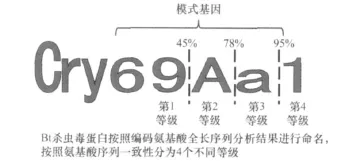
圖1 BtCry毒素命名規(guī)則
營養(yǎng)期殺蟲蛋白(Vips)是蘇云金芽胞桿菌在細胞營養(yǎng)期分泌的一種非晶體的胞外殺蟲蛋白。Estruch等人于1996年和1998年分別從Bt菌株和蠟狀芽胞桿菌(B.cereus)菌株培養(yǎng)物的上清液中分離到Vip殺蟲蛋白[5,11]。該類蛋白從對數(shù)生長中期開始分泌,直到穩(wěn)定前期達到最高峰;一般不形成伴胞晶體,且與已知的ICPs序列相似性很低,最重要的是其殺蟲作用機理與ICPs不同。該蛋白的發(fā)現(xiàn)使Bt在殺蟲活性、殺蟲譜方面得到很大突破,并在一定程度上克服了多種害蟲對ICPs不敏感的弊端。目前Vips命名規(guī)則分為4大類(Vip1、Vip2、Vip3和Vip4),其中Vip1與Vip2是一種二元毒素,兩者需要協(xié)同作用,目前研究顯示這類毒素對鞘翅目害蟲具有殺蟲活性(US5770696,US6656908,US6605701等專利)。Vip3類是一大類對鱗翅目高效的殺蟲蛋白,特別是Vip3Aa類蛋白對草地貪夜蛾、小地老虎等害蟲有較好殺蟲活性,克服了BtCry蛋白對其毒力較低的弱點;Vip4類蛋白是新發(fā)現(xiàn)的一類蛋白,其具體功能還不清楚。
除此之外Bt還編碼其他殺蟲蛋白及其毒力相關(guān)的物質(zhì),例如幾丁質(zhì)酶、蛋白酶、細菌素、增強素、抗菌肽等,這些物質(zhì)在Bt殺蟲過程中都有重要的作用[12]。此外,Sip1A與S-層蛋白是2006年發(fā)現(xiàn)的新型殺蟲蛋白[13-14],兩者都對葉甲科的害蟲有殺蟲活性,其殺蟲機理尚不清楚。
2 Bt殺蟲基因克隆與專利保護現(xiàn)狀
2.1 數(shù)據(jù)來源與分析方法
由于目前應(yīng)用最廣泛的Bt毒素是Cry、Cyt和Vip,本文將主要分析這3種毒素的克隆與專利保護的相關(guān)信息。有關(guān)基因的克隆與命名主要依據(jù)英國薩塞克斯大學(SussexUniversity)生命科學學院Bt毒素命名網(wǎng)站(http:∥www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/)。國外 Bt殺蟲基因?qū)@Wo情況是通過分析GenBank中專利保護氨基酸(PATAA)數(shù)據(jù)庫獲得(http:∥www.ncbi.nlm.nih.gov/genbank/)。PATAA數(shù)據(jù)庫收集了來自美國專利局(USPTO)以及通過歐洲分子生物學實驗室(EMBL)、日本DNA數(shù)據(jù)庫(DDBJ)中的歐洲專利、日本專利所涉及的氨基酸序列。分析已經(jīng)獲得正式命名的Bt模式殺蟲基因,建立本地殺蟲基因數(shù)據(jù)庫,利用本地Bt模式殺蟲基因數(shù)據(jù)庫與PATAA數(shù)據(jù)庫中氨基酸序列進行Blastp比對,挑選PATAA數(shù)據(jù)庫中與Bt模式殺蟲基因相比得分(Score)在60以上的氨基酸序列進行分析,進一步去除無關(guān)序列,獲得目前專利保護的所有Bt殺蟲蛋白序列。cry基因序列在國內(nèi)的專利數(shù)據(jù),通過對中國專利數(shù)據(jù)庫進行檢索,數(shù)據(jù)庫的檢索日期是2011年4月1日,包括數(shù)據(jù)庫中此前已公開的所有專利申請文件。采用關(guān)鍵詞進行檢索,檢索詞如下:蘇云金(桿菌)、蘇云金芽胞桿菌、芽胞桿菌、Bt(基因)等,在摘要、說明書和權(quán)利要求書中同時檢索,并經(jīng)過人工判讀排除不相關(guān)專利。
2.2 Bt殺蟲基因的克隆與命名
自從1981年第1個殺蟲基因被克隆開始,截至2011年4月,有701個殺蟲基因被命名,其中cry基因570個,cyt基因35個,vip基因96個,全球Bt基因克隆的速度總體呈現(xiàn)加速上升的趨勢(圖2)。這些基因來自30多個國家和地區(qū),其中我國最多,有259個。我國Bt基因克隆工作近年來有較大突破,2000年中國農(nóng)業(yè)科學院植物保護研究所克隆到我國第1個模式基因cry1Ie1;2010年我國克隆到的模式基因有31個,占當年全球模式基因總數(shù)的89%。
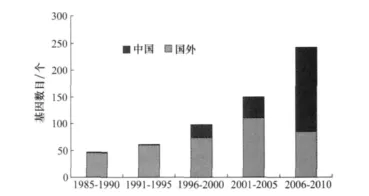
圖2 歷年我國克隆的Bt基因所占比例
2.3 Bt殺蟲基因在國外的專利保護情況
通過對GenBankPATAA數(shù)據(jù)庫氨基酸序列進行比對分析,結(jié)果顯示目前已授權(quán)專利涉及的Bt殺蟲基因氨基酸序列有768個,主要分布在對鱗翅目、鞘翅目害蟲有活性的 cry1、cry2、cry3、cry8、cry9、cry34和cry35中(圖3)。分析結(jié)果還顯示出一個現(xiàn)象,這些序列中有364條目前已命名的模式基因序列相似性小于95%,按照目前命名規(guī)則應(yīng)屬于新型模式基因,占所有受保護序列的47%。這些新基因沒有在任何正式的學術(shù)刊物上發(fā)表,而只是以專利形式出現(xiàn),這些結(jié)果說明國外相關(guān)研究機構(gòu)和企業(yè)已經(jīng)對殺蟲基因命名與發(fā)表文章不感興趣,而致力于占有新基因的知識產(chǎn)權(quán)。

圖3 受專利保護的Bt基因序列類型與數(shù)量
近年來,在國家“863”、“轉(zhuǎn)基因?qū)m棥钡软椖康闹С窒?國內(nèi)不少研究機構(gòu)正在開展Bt基因的分離與轉(zhuǎn)基因工作。由于目前國外大量受專利保護的Bt基因并未提交Bt新毒素國際命名委員會命名,序列等信息處于隱藏狀態(tài),而這批基因與已知的模式基因的一致性都小于95%。這種情況就給我國許多從事Bt新基因分離與轉(zhuǎn)基因研究的人員帶來潛在的知識產(chǎn)權(quán)隱患。因為大家很可能不清楚自己所掌握的新型模式基因是否已經(jīng)被國外專利所覆蓋和保護,極易落入知識產(chǎn)權(quán)陷阱,進而導(dǎo)致在與國外的知識產(chǎn)權(quán)競爭中處于絕對劣勢。隨著國外大型企業(yè)申報中國專利等知識產(chǎn)權(quán)申請的逐年增多,對其專利所保護基因信息的準確掌握顯得尤為重要。
本文按照目前Bt基因命名規(guī)則對已授權(quán)專利中與目前已報道的模式基因的一致性小于95%的Bt基因進行分類,統(tǒng)計相關(guān)專利號,以供國內(nèi)從事Bt基因相關(guān)研究人員參考。表1列出了與目前已命名的模式基因序列相似性大于78%小于95%的序列相關(guān)的專利,按照現(xiàn)行命名規(guī)則,應(yīng)屬于第3等級的新模式基因。表2列出了與目前已命名的模式基因序列相似性大于45%小于78%的序列相關(guān)的專利,按照現(xiàn)行命名規(guī)則,應(yīng)屬于第2等級的新模式基因。表3列出了與目前已命名的模式基因序列相似性小于45%的序列相關(guān)的專利,按照現(xiàn)行命名規(guī)則,應(yīng)屬于第1等級的新模式基因。
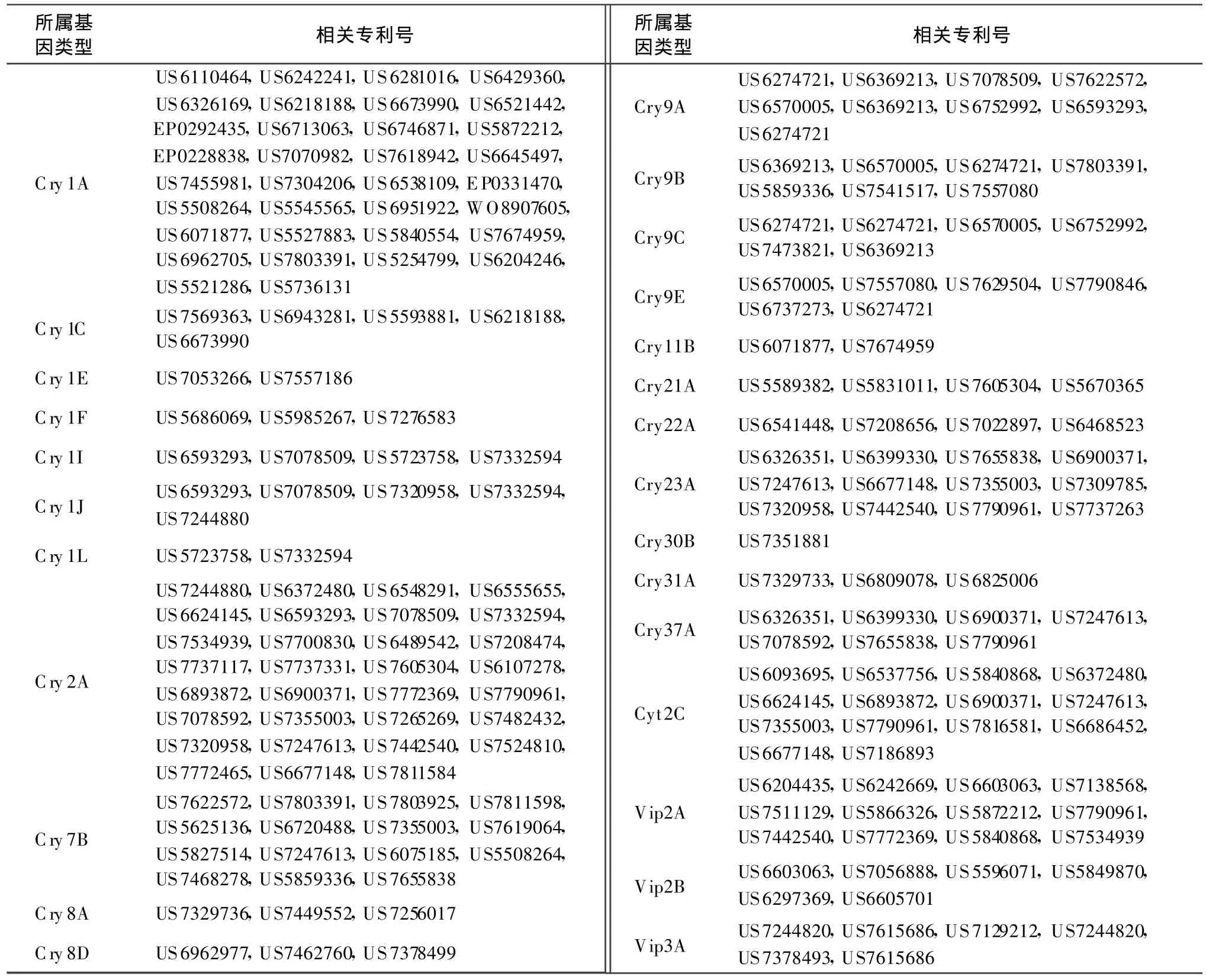
表1 涉及第 3等級的 Bt新基因的專利
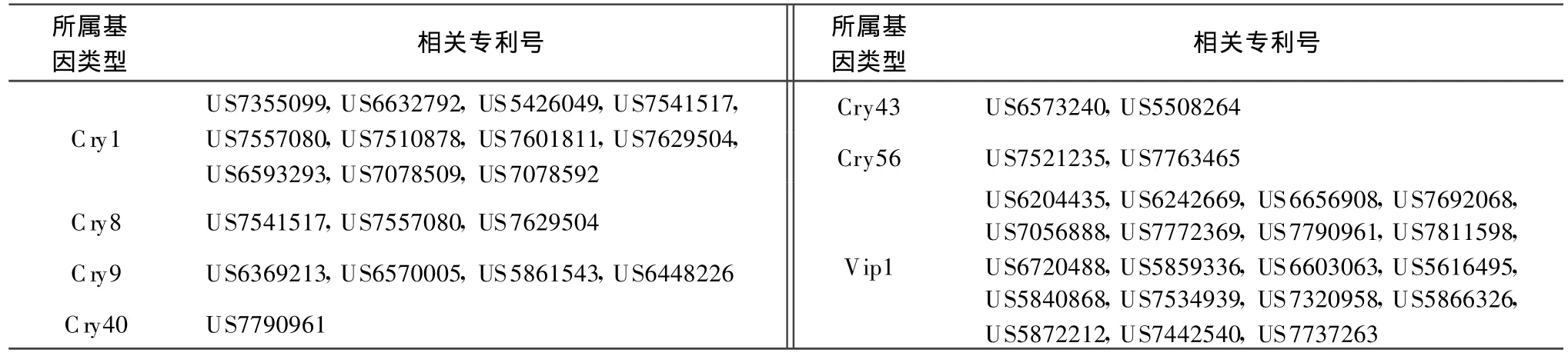
表2 涉及第 2等級的 Bt新基因的專利
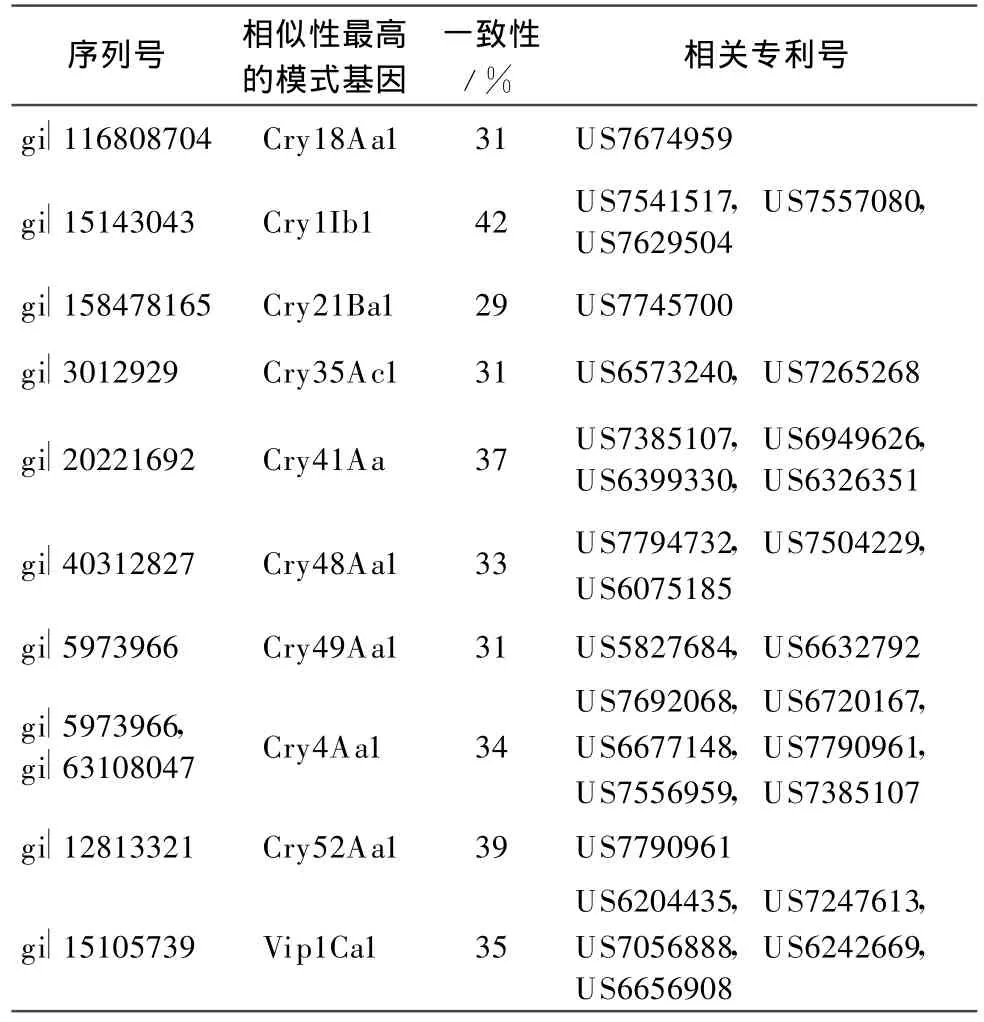
表3 涉及第1等級的Bt新基因及其專利
2.4 Bt基因在國內(nèi)的專利保護現(xiàn)狀
通過檢索,獲得Bt相關(guān)專利260件,剔除Bt菌株、基因改造及Bt殺蟲劑相關(guān)專利,獲得Bt殺蟲基因相關(guān)專利47件,其中國內(nèi)研究機構(gòu)申請的有21件,占44.7%。在受保護基因類型標注清楚程度來看,國內(nèi)研究機構(gòu)申請的專利標注明顯、準確,通過專利判讀,可以準確了解專利所保護的基因類型;而國外研究機構(gòu)申請的專利對其權(quán)利要求基因往往不標注,或者標注不準確,這種現(xiàn)象近年來尤為突出。比如2011年先鋒國際良種公司在華申請的5個專利中,在摘要、題名甚至專利全文都沒有說明其保護準確基因名稱。而2004年拜爾生物科學公司專利(zl1484702)所要求的“cry2Ae” ,“cry2Af” ,“cry2Ag”3 個基因中 ,只有cry2Ae與目前毒素命名網(wǎng)站(http:∥www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/)命名一致,而“cry2Af”、“cry2Ag”則與命名網(wǎng)站對應(yīng)模式基因序列有較大差異,屬于公司個體行為,不能按目前基因命名規(guī)則對其準確描述。此外,從申請專利數(shù)量上看,國外研究機構(gòu)加速了在中國知識產(chǎn)權(quán)的布局,2010年申請的8件專利中,國內(nèi)外各占4件,與之前比例相當;而2011年4月份之前申請的6件專利中,國外就占到5件,整體呈現(xiàn)速度加快的趨勢。表4列出的是外國研究機構(gòu)在中國申請專利保護的基因情況,以供相關(guān)研究人員參考。

表4 2000-2011年國外企業(yè)在中國申請的Bt基因?qū)@?/p>

續(xù)表4
3 總結(jié)與展望
通過對Bt基因命名網(wǎng)站的數(shù)據(jù)分析統(tǒng)計,結(jié)果顯示我國相關(guān)研究取得了令人矚目的成就,有大量新型模式基因被分離克隆,特別是近年來所命名的新基因占全球新基因的絕大部分。而當我們分析國外授權(quán)專利所保護的Bt殺蟲基因的時候發(fā)現(xiàn),雖然國外研究機構(gòu)命名的Bt基因數(shù)目在減少,但是其專利保護的新型Bt基因序列數(shù)量卻是非常巨大的,并呈上升趨勢。這些基因通常不在正式的學術(shù)刊物上發(fā)表,只有通過檢索相關(guān)專利數(shù)據(jù)庫才能獲得相關(guān)信息;即便檢索到相關(guān)專利,由于其專利保護基因標注不明確,無法獲取這些新型基因的更多信息,這些情況容易導(dǎo)致國內(nèi)相關(guān)研究人員在與國際同行競爭過程中陷入被動挨打的境地。
針對這種情況,密切跟蹤、整理和分析國外主要農(nóng)業(yè)生物技術(shù)研究機構(gòu)和跨國公司所擁有的Bt基因?qū)@麪顩r是當前一項刻不容緩的工作。國內(nèi)Bt基因研究人員應(yīng)當密切跟蹤外國主要農(nóng)業(yè)生物技術(shù)研究機構(gòu)的專利申請動態(tài),通過生物信息學、情報學研究方法對其進行整理,形成易讀的研究報告,為我國相關(guān)管理部門和科研人員提供決策依據(jù)和參考。
目前,國內(nèi)Bt基因?qū)@麛?shù)量與質(zhì)量都處于劣勢,要改變目前狀況需要從多方位進行創(chuàng)新[4]。首先,國外企業(yè)在申請專利的推動力方面占有較大優(yōu)勢。獲得具有開發(fā)前景的Bt基因、并將這些基因和技術(shù)轉(zhuǎn)化為獨占的權(quán)利從而獲得最大化的經(jīng)濟利益是國外大型企業(yè)的推動力,是其生存與發(fā)展的根本。而國內(nèi)研究主體是科研院所與大專院校,主要依靠國家研究項目推動Bt基因研究,這使得國內(nèi)研究機構(gòu)在Bt基因知識產(chǎn)權(quán)方面動力不足,難以在競爭中占據(jù)優(yōu)勢。目前,主要國外公司掌握了全世界70%以上的Bt殺蟲基因?qū)@?并且被批準商業(yè)化的cry、vip3A基因轉(zhuǎn)化事件幾乎全被孟山都、陶氏益農(nóng)、先鋒、先正達、拜耳等公司掌控。其次,國內(nèi)單位上中下游脫節(jié),缺乏整體有機的密切配合機制。國外大型企業(yè)不僅掌握了絕大部分基因,并且這些基因涉及的專利質(zhì)量大多高出國內(nèi)單位的專利,其權(quán)利要求范圍涵蓋較寬、技術(shù)全面,包含了基因、載體、宿主菌、轉(zhuǎn)化方法等Bt基因應(yīng)用的各個方面,從上游至下游技術(shù)都囊括其中。這樣的專利文件最大限度地保證了其相關(guān)基因在應(yīng)用過程中的生命力,避免了競爭對手在其中某一環(huán)節(jié)對其基因?qū)@M行絞殺。形成這樣的專利需要一個集成了Bt菌株分離與基因克隆平臺、農(nóng)業(yè)害蟲飼養(yǎng)與測定等功能驗證平臺、基因改造修飾技術(shù)平臺、轉(zhuǎn)基因植物技術(shù)平臺,甚至作物品種資源與選育技術(shù)平臺的大型研究機構(gòu)進行全面系統(tǒng)的合作。在國外,孟山都、先鋒等公司都具備這樣的實力,相比之下國內(nèi)研究機構(gòu)和國內(nèi)企業(yè)顯得勢單力薄,沒有一家研究單位或企業(yè)可以包攬所有的環(huán)節(jié)流程,而又缺乏高效的合作機制,從而很難同國外企業(yè)競爭。因此,創(chuàng)新上中下游合作機制,讓企業(yè)成為成果產(chǎn)業(yè)化主體,協(xié)調(diào)基因克隆、遺傳轉(zhuǎn)化和轉(zhuǎn)基因新品種培育之間的緊密協(xié)作,形成利益共享、風險共擔的合作機制,加大Bt基因在各個環(huán)節(jié)中的知識產(chǎn)權(quán)保護力度,讓市場成為推動國內(nèi)Bt基因研究與知識產(chǎn)權(quán)保護真正動力,是保障我國的轉(zhuǎn)基因植物產(chǎn)業(yè)化早日走上良性發(fā)展的必由之路。
[1]James C.Global status of commercialized biotech/GM crops:2010[G].International Service for the Acquisition of Ag ri-biotech Applications,2010,Brief 42.
[2]Shu C,Zhang J.Current patents related to Bacillus thuringiensis insecticidal crystal proteins[J].Recent Pat DNA Gene Seq,2009,3(1):26-28.
[3]鄒吉承.中國Bt基因相關(guān)知識產(chǎn)權(quán)專利保護現(xiàn)狀及趨勢[J].中國棉花,2007(7):2-5.
[4]宋敏,林祥明,劉麗軍.Cry基因家族的專利分布研究[J].生物技術(shù)通報,2010(1):1-8.
[5]Schnepf E,Crickmore N,Van R J,et al.Bacillusthuringiensis and its pesticidal crystal proteins[J].Microbiol M ol Biol Rev,1998,62(3):775-806.
[6]Bravo A,Likitvivatanavong S,Gill S,et al.Bacillus thuringiensis:A story of a successful bioinsecticide[J].Insect Biochem Mol Biol,2011(2):1-9.
[7]Mohamed A I,Natalya G,Matthew J,et al.Bacillus thuringiensis A genomics and proteomics perspective[J].BioengineeredBugs,2010,1(1):31-50.
[8]Schnepf H E,Whiteley H R.Cloning and expression of the Bacillus thuringiensis crystal protein gene in Escherichia coli[J].Proc Natl Acad Sci USA,1981,78(5):2893-2897.
[9]Crickmore N,Zeigler D R,Feitelson J,et al.Revision of the nomenclature for the Bacillus thuringiensis pesticidal crystal proteins[J].Microbiol Mol Biol Rev,1998,62(3):807-813.
[10]de Maagd R A,Bravo A,Crickmore N.How Bacillus thuringiensis has evolved specific toxins to colonize the insect world[J].Trends Genet,2001,17(4):193-199.
[11]Estruch J J,Warren G W,Mullins M A,et al.Vip3A,a novel Bacillus thuringiensis vegetative insecticidal protein with a wide spectrum of activities against lepidopteran insects[J].Proc Natl Acad Sci USA,1996,93(11):5389-5394.
[12]Raymond B,Johnston P R,Nielsen-Leroux C,et al.Bacillus thuringiensis:an important pathogen?[J].T rends in Microbiology,2010,18(5):189-194.
[13]Donovan W P,Eng leman J T,Donovan J C,et al.Discovery and characterization of Sip1A:A novel secreted protein from Bacillus thuringiensis with activity against coleopteran larvae[J].Appl Environ Microbiol,2006,72(4):713-719.
[14]Pena G,Miranda-Rios J,de La R G,et al.A Bacillus thuringiensis S-layer protein involved in toxicity against Epilachna varivestis(Coleoptera:Coccinellidae)[J].Appl Environ Microbiol,2006,72(1):353-360.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務(wù)財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
