內(nèi)毛細胞帶狀突觸結(jié)構(gòu)及功能的研究進展
2011-07-28 03:21:06陳麗平綜述商秀麗審校
醫(yī)學綜述 2011年19期
關(guān)鍵詞:結(jié)構(gòu)功能
陳麗平(綜述),商秀麗(審校)
(中國醫(yī)科大學附屬第一醫(yī)院神經(jīng)內(nèi)科,沈陽110001)
耳蝸能夠編碼不同頻率和強度的聲音信號,這一過程由內(nèi)毛細胞及外毛細胞參與。外毛細胞對聲音信號具有放大作用,使耳蝸對聲音頻率有高度的敏感性和選擇性[1]。內(nèi)毛細胞將不同頻率、強度、時間性質(zhì)的聲音信號傳遞給中樞神經(jīng)系統(tǒng)。內(nèi)毛細胞將聲波的振動由化學信號轉(zhuǎn)換為電信號,形成電勢差,刺激神經(jīng)遞質(zhì)釋放到相應(yīng)的螺旋神經(jīng)元,再由螺旋神經(jīng)元投射到中樞神經(jīng)系統(tǒng)。聽感覺突觸——帶狀突觸對信號的傳遞必需準確,并且能維持很長一段時間。毛細胞上帶狀突觸的形態(tài)和數(shù)量對聽覺系統(tǒng)具有重要作用[2,3]。帶狀突觸在形態(tài)學和功能學上具有獨特性:其活動區(qū)只位于幾個固定的區(qū)域,即快速囊泡釋放區(qū),但其主要特征是被稱為帶狀突觸的結(jié)構(gòu)。帶狀突觸通過動態(tài)作用精確、快速而又持續(xù)地釋放囊泡,該過程非常復(fù)雜[4]。但是其精確的功能和作用方式是如何完成的仍不清楚。
1 帶狀突觸的超微結(jié)構(gòu)
帶狀突觸指位于突觸前膜的特殊結(jié)構(gòu),因其呈帶狀而被命名[5]。內(nèi)耳的內(nèi)毛細胞通過帶狀突觸將不同的聲音信號傳遞給聽覺傳入神經(jīng)。內(nèi)毛細胞形成帶狀突觸的突觸前膜,傳入神經(jīng)形成具有突觸后電子致密體的突觸后膜。帶狀突觸的囊泡分為三種:①胞質(zhì)內(nèi)游離囊泡;②與帶狀突觸相連的囊泡;③在帶狀突觸小體的底端為錨定突觸前膜的囊泡(圖1)。不同類型的細胞,帶狀突觸的結(jié)構(gòu)、數(shù)量及大小不同,即使在同一個毛細胞上,帶狀突觸的形狀也不相同。帶狀突觸主要存在于毛細胞、視網(wǎng)膜上[6]。在視網(wǎng)膜上,視桿細胞、視錐細胞和雙極細胞形成帶狀突觸;內(nèi)耳帶狀突觸位于內(nèi)毛細胞及前庭毛細胞下[5]。在不同動物耳蝸軸上的不同位置以及不同發(fā)育階段,帶狀突觸的形態(tài)、大小及數(shù)量均有所不同,甚至在同一個毛細胞的不同活性區(qū)域上帶狀突觸的閾值、形態(tài)、大小及數(shù)量也不同[4]。隨著聽力水平的變化,帶狀突觸的形態(tài)和數(shù)量也發(fā)生變化。越來越多的研究表明,即使在一個哺乳動物耳蝸的毛細胞內(nèi),不同帶狀突觸上的鈣離子通道數(shù)量也不同[7]。因而,帶狀突觸后傳入纖維對聲音信號的敏感性相差很大。在不同動物及不同部位毛細胞上,毛細胞的傳入神經(jīng)支配數(shù)量相差很大,如前庭傳入纖維形成分支連接毛細胞上大約有20個甚至上百個帶狀突觸信息的釋放與傳入。相反,哺乳類動物耳蝸內(nèi)毛細胞上的一個帶狀突觸通常只與Ⅰ型傳入纖維的一個樹突相連[8]。毛細胞的帶狀突觸(也叫致密小體)具有形態(tài)特異性,可以為球形、平面或長方體形,大小不一,聚集20~400個囊泡。非哺乳類毛細胞上的帶狀突觸可能更大(直徑為200~400 nm),但哺乳類的毛細胞帶狀突觸的長度≤200 nm,與視網(wǎng)膜上雙極細胞的帶狀突觸大小相近。哺乳類動物耳蝸毛細胞帶狀突觸聚集100~200個囊泡,在每個帶狀突觸上伴隨100~200個電壓-門控鈣通道[7]。
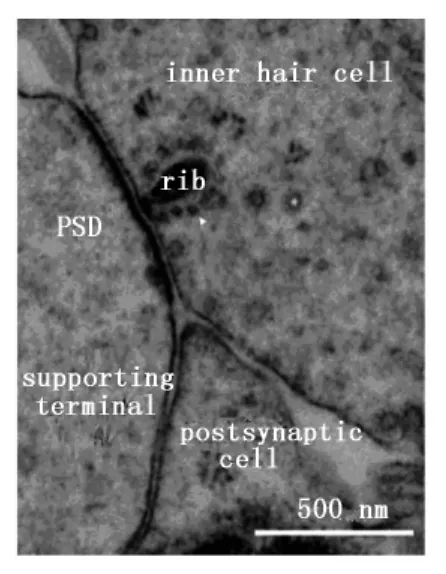
圖1 內(nèi)毛細胞下帶狀突觸的結(jié)構(gòu)
2 帶狀突觸的分子組成
RIBEYE是帶狀突觸上所知的唯一結(jié)構(gòu)蛋白[9],可能具有酶活性,并具有C-末端結(jié)合蛋白1(C-terminal binding protein 1,CtBP1)[10]轉(zhuǎn)錄共抑制及溶血磷脂酸乙酰輔酶轉(zhuǎn)移酶活性[11]。已經(jīng)克隆出RIBEYE的cDNA,RIBEYE具有一個特異的氨基末端富含脯氨酸的A區(qū)域,這是在其他細胞上所沒有的蛋白結(jié)構(gòu),其與CtBP2具有基因同源性。
CtBP1與RIBEYE的B區(qū)域相關(guān),是帶狀突觸的一個組成部分[5]。CtBP1與高爾基的分裂相關(guān),但其在帶狀突觸上的功能至今尚不清楚。CtBP1和CtBP2/RIBEYE(B)-區(qū)域連接至位于帶狀突觸活性區(qū)域的胞質(zhì)蛋白Bassoon上。Bassoon與Piccolo一樣,都是位于活性區(qū)域的細胞基質(zhì)大分子物質(zhì),很有可能與活性區(qū)域上帶狀突觸的錨定有關(guān)[12,13]。帶狀突觸減少可造成突觸病理源性的聽力損傷,即耳蝸的擴音能力正常,而聽覺誘發(fā)電位異常。Rab3反應(yīng)蛋白對于啟動囊泡釋放和Ca2+依賴性囊泡釋放具有重要的作用[14]。KIF3A是驅(qū)動蛋白Ⅱ的構(gòu)成部分[15]。因為在帶狀突觸上只找到了KIF3A,而未找到驅(qū)動蛋白Ⅱ的其他構(gòu)成部分,因此對帶狀突觸的具體功能至今還不清楚。在視網(wǎng)膜的動力蛋白還有MyosinVa和MyosinⅦa,MyosinⅤa基因突變時,將出現(xiàn)畸形帶狀突觸[4]。
在耳蝸毛細胞帶狀突觸上含有一個C2區(qū)域的跨膜蛋白Otoferlin。OTOF基因有28個編碼外顯子,在成年鼠的內(nèi)毛細胞和壺腹晴的Ⅰ、Ⅱ型毛細胞中都有表達(Ⅱ型毛細胞中表達微弱),其編碼的蛋白質(zhì)是Otoferlin,由1997個氨基酸組成,相對分子質(zhì)量量為140.5 ×103[16]。它是帶狀突觸信號釋放所必需的結(jié)構(gòu)成分,而在一些耳聾患者中Otoferlin也發(fā)生了突變[17]。研究發(fā)現(xiàn),Otoferlin位于與帶狀突觸相連的囊泡上,它的主要功能是將Ca2+與水溶性N-乙基馬來酰亞胺敏感因子黏附蛋白受體突觸融合蛋白1和突觸連接蛋白體連接在一起,使突觸囊泡膜與突觸前膜融合,通過胞吐作用釋放神經(jīng)遞質(zhì)[13]。目前,對于這些帶狀突觸所特有的復(fù)合物的具體功能還不是很清楚。盡管Otoferlin和其他很多鈣結(jié)合蛋白在耳蝸毛細胞中起重要作用,但具體功能還不確定[18,19]。而且,不同細胞類型的帶狀突觸引起突觸囊泡釋放鈣感受器的敏感性不同。鈣感受器具有高親和性和低協(xié)同性,這就允許了個別鈣通道的任意開放,從而興奮信號能使突觸后靶點除極,引起突觸囊泡釋放[20]。相反,耳蝸毛細胞具有低親和性和高協(xié)同性,適合于高同步性釋放的特征[21]。
3 帶狀突觸的可塑性
即使在一種細胞類型上,帶狀突觸的結(jié)構(gòu)、功能也可能不同。帶狀突觸具有動態(tài)結(jié)構(gòu),其結(jié)構(gòu)改變可由自身改變或外界環(huán)境改變所引起。在光照下,某種結(jié)構(gòu)從小鼠視桿細胞帶狀突觸上解離出來,無光照時,該物質(zhì)又回到帶狀突觸上。帶狀突觸結(jié)構(gòu)的改變反映了其功能的變化,且與突觸的活性相關(guān)。當光感受突觸釋放大量神經(jīng)遞質(zhì)時(在黑暗中),帶狀突觸延長;在光照時帶狀突觸縮短,胞吐功能降低[22]。毛細胞帶狀突觸的形狀與其功能性質(zhì)也密切相關(guān)。在耳蝸高頻和低頻區(qū)域上帶狀突觸的結(jié)構(gòu)不同導(dǎo)致胞吐作用對Ca2+的依賴性不同[12],而在高頻聽力的帶狀突觸表面積更大。這些反映了帶狀突觸的數(shù)量與帶狀突觸的功能具有相關(guān)性。
4 帶狀突觸的胞吐作用
現(xiàn)在關(guān)于帶狀突觸功能的認識主要基于兩類實驗:毛細胞突觸前膜電容的電壓-驅(qū)動的改變和對傳入神經(jīng)興奮性突觸后電位的記錄。它們的主要差別在于電容是記錄整個細胞上所有帶狀突觸的變化,而興奮性突觸后電位顯示的是單個帶狀突觸的釋放。突觸前電容記錄與突觸后興奮性突觸后電位記錄都反映了瞬時釋放與鈣離子內(nèi)流的增多呈線性關(guān)系。因此,鈣通道打開后興奮性突觸后電位增高,給予強的鈣離子阻滯劑后興奮性突觸后電位降低。
通過高分辨率顯像技術(shù)發(fā)現(xiàn),雖然在帶狀突觸上囊泡釋放位置不是嚴格局限于帶狀上,但連在帶狀上的囊泡是胞吐“熱點”[23]。通過膜片鉗技術(shù)可以詳細地記錄到突觸遞質(zhì)的釋放始于電壓門控鈣通道的開啟[24]。在哺乳類動物內(nèi)耳毛細胞上的100~200個鈣通道中,僅需一小部分(數(shù)十個)鈣通道的開放就可引起突觸囊泡釋放。因此,在活動區(qū)域鈣通道的總數(shù)量對囊泡的釋放具有重要意義。
5 帶狀突觸的特性
同光感受器細胞一樣,內(nèi)毛細胞傳入神經(jīng)突觸缺少突觸素、突觸體素以及突觸結(jié)合蛋白1和2[25]。突觸素在突觸的囊泡運動中發(fā)揮作用,突觸體素幾乎存在于所有的神經(jīng)終末,而突觸結(jié)合蛋白在中樞神經(jīng)系統(tǒng)和神經(jīng)內(nèi)分泌細胞的囊泡融合中起鈣傳感器的作用[26]。
研究發(fā)現(xiàn),帶狀突觸對突觸囊泡的持續(xù)融合起重要作用,帶狀突觸的每個活動域都擁有一個準備囊泡釋放池,以維持囊泡的持續(xù)釋放[5]。依據(jù)細胞類型的不同,帶狀突觸準備囊泡釋放池數(shù)也不同。電鏡觀察到,突觸囊泡的釋放至少有兩個動力部分組成:①快速釋放,在幾毫秒內(nèi)就能釋放。②慢釋放,在強刺激時能維持數(shù)百毫秒的釋放。刺激強度越低,釋放時間越長。每個準備釋放池有數(shù)十個囊泡,可在幾微秒內(nèi)釋放。這意味著每個活動域都可能同時釋放大量囊泡,從而獲得快速的遞質(zhì)釋放。
6 帶狀突觸的胞吞作用
胞吞作用與胞吐作用的平衡對避免突觸囊泡耗竭具有重要意義[27]。突觸末端細胞膜的再攝取非常困難,因為此過程需要大的胞膜翻轉(zhuǎn)[28]。通過電生理分析顯示在視網(wǎng)膜和內(nèi)毛帶狀突觸上至少有兩種動力模式參與胞吞過程[27]:快速攝取機制(每次只持續(xù)1~2 s)和緩慢模式(10~30 s)。快速過程受突觸前Ca2+的調(diào)節(jié)和細胞內(nèi)Cl-選擇性抑制的調(diào)節(jié),緩慢胞吞作用具有Ca2+依賴性。電子顯微鏡跟蹤分析顯示帶狀突觸前膜管狀內(nèi)陷入胞內(nèi)[29]。
7 問題與展望
關(guān)于帶狀突觸的報道雖然很多,但其中仍有很多問題不明確,如囊泡是如何連接至帶狀突觸上的?它們間又是如何調(diào)節(jié)的?作為囊泡的傳送帶,啟動裝置帶狀突觸是如何影響這些功能的?不同成分對帶狀突觸精確、快速釋放的作用?要回答這些問題,需要遺傳學、細胞生物學、生理學及成像技術(shù)等多個學科的共同參與。應(yīng)用新的技術(shù)(如在活細胞的高分辨率的光學成像、捕獲囊泡的超微方法及電生理實驗中改變帶狀突觸功能等方法)有助于理解和觀察突觸的特性。
[1]Dallos P,Corey ME.The role of outer hair cell motility in cochlear tuning[J].Curr Opin Neurobiol,1991,1(2):215-220.
[2]Liu K,Li S,Jiang X.Quantitative analysis of the ribbon synapse number of cochlear inner hair cells in C57BL/6J mice using the three-dimensional modeling method[J].Sci China C Life Sci,2009,52(9):807-812.
[3]施磊,趙寧,陳麗平,等.C57BL/6J小鼠耳蝸內(nèi)毛細胞傳入神經(jīng)突觸的免疫學標記觀察及其意義[J].中華耳科學雜志,2010,8(4):456-460.
[4]LoGiudice L,Matthews G.The role of ribbons at sensory synapses[J].Neuroscientist,2009,15(4):380-391.
[5]Nouvian R,Beutner D,Parsons TD,et al.Structure and function of the hair cell ribbon synapse[J].J Membr Biol,2006,209(2/3):153-165.
[6]Thoreson WB.Kinetics of synaptic transmission at ribbon synapses of rods and cones[J].Mol Neurobiol,2007,36(3):205-223.
[7]Meyer AC,F(xiàn)rank T,Khimich D,et al.Tuning of synapse number,structure and function in the cochlea[J].Nature Neurosci,2009,12(4):444-453.
[8]Matthews G,F(xiàn)uchs P.The diverse roles of ribbon synapses in sensory neurotransmission[J].Nat Rev Neurosci,2010,11(12):812-822.
[9]Zenisek D,Davila V,Wan L,et al.Imaging calcium entry sites and ribbon structures in two presynaptic cells[J].J Neurosci,2003,23(7):2538-2548.
[10]tom Dieck S,Altrock WD,Kessels MM,et al.Molecular dissection of the photoreceptor ribbon synapse:physical interaction of Bassoon and RIBEYE is essential for the assembly of the ribbon complex[J].J Cell Biol,2005,168(5):825-836.
[11]Valente C,Spano S,Luini A,et al.Purification and functional properties of the membrane fissioning protein CtBP3/BARS[J].Methods Enzymol,2005,404:296-316.
[12]Khimich D,Nouvian R,Pujol R,et al.Hair cell synaptic ribbons are essential for synchronous auditory signaling[J].Nature,2005,434(7035):889-894.
[13]Martinez-Dunst C,Michaels RL,F(xiàn)uchs PA.Release sites and calcium channels in hair cells of the chick's cochlea[J].J Neurosci,1997,17(23):9133-9144.
[14]Castillo PE,Schoch S,Schmitz F,et al.RIM1alpha is required for presynaptic long term potentiation[J].Nature,2002,415(6869):327-330.
[15]Muresan V,Lyass A,Schnapp BJ.The kinesin motor KIF3A is a component of the presynaptic ribbon in vertebrate photoreceptors[J].J Neurosci,1999,19(3):1027-1037.
[16]Yasunaga S,Grati M,Cohen-Salmon M,et al.A mutation in OTOF,encoding otoferlin,a FER-like protein,causes DFN9,a nonsyndromic form of deafness[J].Nature Genetics,1999,21(4):363-369.
[17]Roux I,Safieddine S,Nouvian R,et al.Otoferlin,defective in a human deafness form,is essential for exocytosis at the auditory ribbon synapse[J].Cell,2006,127(2):277-289.
[18]Johnson SL,F(xiàn)ranz C,Kuhn S,et al.Synaptotagmin Ⅳ determines the linear Ca2+dependence of vesicle fusion at auditory ribbon synapses[J].Nature Neurosci,2010,13(1):45-52.
[19]Pangrsic T,Lasarow L,Reuter K,et al.Hearing requires otoferlindependent efficient replenishment of synaptic vesicles in hair cells[J].Nature Neurosci,2010,13(7):869-876.
[20]Duncan G,Rabl K,Gemp I,et al.Quantitative analysis of synaptic release at the photoreceptor synapse[J].Biophys J,2010,98(10):2102-2110.
[21]Beutner D,Voets T,Neher E,et al.Calcium dependence of exocytosis and endocytosis at the cochlear inner hair cell afferent synapse[J].Neuron,2001,29(3):681-690.
[22]Spiwoks-Becker I,Glas M,Lasarzik I,et al.Mouse photoreceptor synaptic ribbons lose and regain material in response to illumination[J].Eur J Neurosci,2004,19(6):1559-1571.
[23]Midorikawa M,Tsukamoto Y,Berglund K,et al.Different roles of ribbon-associated and ribbon-free active zones in retinal bipolar cells[J].Nat Neurosci,2007,10(10):1268-1276.
[24]Platzer J,Engel J,Schrott-Fischer A,et al.Congenital deafness and sinoartrial node dysfunction in mice lacking class D L-type Ca2+channels[J].Cell,2000,102(1):89-97.
[25]Zanazzi G,Matthews G.The molecular architecture of ribbon presynaptic terminals[J].Mol Neurobiol,2009,39(2):130-148.
[26]Safieddine S,Wenthold RJ.SNARE complex at the ribbon synapse of cochlear hair cells:analysis of synaptic vesicle and synaptic membrane-associated proteins[J].Eur J Neurosci,1999,11(3):803-812.
[27]Wu LG,Ryan TA,Lagnado L.Modes of vesicle retrieval at ribbon synapses,calyx-type synapses,and small central synapses[J].J Neurosci,2007,27(44):11793-11802.
[28]Khimich D,Nouvian R,Pujol R,et al.Hair cell synaptic ribbons are essential for synchronous auditory signaling[J].Nature,2005,434(7035):889-894.
[29]Lenzi D,Crum J,Ellisman MH,et al.Depolarization redistributes synaptic membrane and creates a gradient of vesicles on the synaptic body at a ribbon synapse[J].Neuron,2002,36(4):649-659.
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
中國中醫(yī)藥現(xiàn)代遠程教育(2014年22期)2014-03-01 04:32:55
