鷹嘴豆芽素A通過PPARα/γ抑制脂多糖誘導豬外周血單個核細胞炎性細胞因子的分泌①
2011-07-30 13:32:56邱龍新林一平林臻楨林麗欽戴愛玲楊小燕龍巖學院生命科學學院龍巖364000
中國免疫學雜志 2011年10期
邱龍新 林一平 林臻楨 林麗欽 戴愛玲 楊小燕 (龍巖學院生命科學學院,龍巖364000)
植物異黃酮是一類與人類生活息息相關的植物雌激素,長期以來其雌激素樣作用、抗腫瘤作用、心血管保護作用和抗氧化作用受到廣泛重視和深入研究。最近,植物異黃酮在抗炎癥方面的作用引起了越來越多的關注[1]。紅車軸草異黃酮提取物能夠顯著抑制LPS誘導小鼠巨噬細胞RAW264.7分泌TNF-α、IL-6等炎性細胞因子,說明該異黃酮提取物具有抗炎癥的作用[2]。大豆異黃酮、鷹嘴豆異黃酮提取物等植物異黃酮也都具有類似的抗炎作用。大豆異黃酮主要含有染料木黃酮、大豆素等異黃酮成分,而紅車軸草異黃酮主要含有鷹嘴豆芽素A、刺芒柄花素、染料木黃酮、大豆素等4種異黃酮成分。目前,大豆異黃酮的抗炎作用被認為主要是染料木黃酮的作用。染料木黃酮的抗炎作用和機制已經得到較為深入的研究[1],但是,對于鷹嘴豆芽素A的抗炎作用卻少有報道,對于其機制的研究更是尚未見報道。
過氧化物酶體增殖物激活受體(Peroxisome proliferator-activated receptors,PPARs)是配體激活的轉錄因子核受體超家族成員之一,目前已知有三種亞型:PPARα、β/δ和γ。PPARs在免疫細胞上廣泛表達,近年來的研究發現,PPARs在抵抗炎性疾病和免疫調節方面具有重要作用[3]。有趣的是,鷹嘴豆芽素A、刺芒柄花素、染料木黃酮等具有PPARα/γ雙激動劑活性,一些研究據此推測這些異黃酮的抗炎作用與其PPARα/γ雙激動劑活性有關[2,4]。但是,目前并沒有直接的證據證明鷹嘴豆芽素A、刺芒柄花素、染料木黃酮等通過PPARα/γ途徑調控炎癥反應。另外,在單核細胞和淋巴細胞中PPARα與PPARγ均有較多表達而在巨噬細胞中PPARα幾乎不表達[5,6]。為了研究鷹嘴豆芽素A對免疫細胞分泌炎性細胞因子的影響及其與PPARα/γ的關系,以外周血單個核細胞(Peripheral blood mononuclear cells,PBMCs)為研究材料將較為合適。綜上,本研究將因此利用PBMCs探討鷹嘴豆芽素A的抗炎作用及其是否通過PPARα/γ途徑調控炎癥反應。
1 材料與方法
1.1 材料 豬血采自福建龍巖龍馬種豬場;Ficollpague淋巴細胞分離液購自GE Healthcare公司;RPMI1640細胞培養基、Trizol購自Invitrogen公司;LPS、鷹嘴豆芽素 A、GW9662、MK886、羊抗兔 IgG-HRP 等購自 Sigma公司;NF-κB p65抗體購自 Cell Signaling公司;PCRmix購自廣州東盛生物科技有限公司;細胞核蛋白與細胞漿蛋白抽提試劑盒、BCA蛋白濃度測定試劑盒購自碧云天生物技術研究所;其它試劑為國產分析純試劑。
1.2 方法
1.2.1 豬PBMCs的分離與藥物處理 采集豬血自150日齡杜×長×大母豬,利用Ficoll-paque淋巴細胞分離液(密度1.077 g/ml)按廠家說明分離豬PBMCs。細胞懸浮于RPMI1640培養基(含10%胎牛血清、100 U/ml青霉素和0.1 mg/ml鏈霉素)并以1×107個/ml的密度將細胞接入24孔板,在 5%CO2,37℃條件下培養。加入待研究藥物孵育3小時,然后加入LPS(終濃度1μg/ml)進行刺激。
1.2.2 PBMCs分泌 TNF-α、IL-6的測定 LPS處理細胞6小時后收集細胞培養液,2 000 r/min離心15分鐘后收集上清液,利用ELISA試劑盒(R&D systems)按廠家說明測定上清液中TNF-α、IL-6的含量。
1.2.3 RT-PCR分析TNF-α、IL-6 mRNA 的表達LPS處理細胞6小時后,用Trizol試劑提取細胞總RNA,利用 RevertAidTMFirst Strand cDNA Synthesis試劑盒(Fermentas)按廠家說明將RNA反轉錄成cDNA,PCR 擴增 TNF-α、IL-6,以 β-Actin作為內參。引物序列如下,Sense:5′-CGTTGTAGCCAATGTCAAAGCC-3′,Antisense:5′-TGCCCAGATTCAGCAAAGTCCA-3′(TNF-α);Sense:5′-CCAGGAACCCAGCTATGAAC-3′,Antisense:5′-CTGCAGAGCCTCGACATT-3′(IL-6);Sense:5′-CTGGCATTATCATAAACTCT-3′,Antisense:5′-GCGATGATCTTGATCTTCAT-3′(β-Actin)。PCR 產物經1.5%瓊脂糖凝膠電泳分離后用溴化乙錠顯色。
1.2.4 NF-κB活性的分析 NF-κB的激活可以通過它從胞漿移位至胞核來判定。LPS處理細胞30分鐘后,用細胞核蛋白與細胞漿蛋白抽提試劑盒按廠家說明進行操作分別獲得胞漿與胞核蛋白。用Western blot技術檢測NF-κB的進核情況。一抗(NF-κB p65)以1∶1 000濃度4℃孵育過夜,二抗(羊抗兔)以1∶2 000濃度孵育2小時后顯影。
1.3 數據處理與統計學分析 實驗數據采用GraphPad Prism 5.0軟件處理。組間用One-Way ANOVA with Newman-Keuls Multiple Comparison Test比較。當P值<0.05時差異被認為具有統計學意義。
2 結果
2.1 鷹嘴豆芽素A對LPS刺激豬PBMCs后TNF-α、IL-6分泌量和mRNA表達的影響 TNF-α是一種與炎癥反應密切相關的促炎癥細胞因子。ELISA結果顯示,豬PBMCs在經LPS刺激后,TNF-α的分泌較未經LPS刺激時大幅上調(P<0.001,圖1A)。而在加入LPS刺激前3小時用鷹嘴豆芽素A預處理細胞則能顯著抑制由LPS刺激引起的TNF-α的分泌,終濃度為100μmol/L的鷹嘴豆芽素A(P<0.001)比50 μmol/L的(P<0.01)抑制作用效果更明顯。利用RT-PCR分析mRNA表達的結果進一步證明了鷹嘴豆芽素A抑制TNF-α生成的作用(圖1B)。
IL-6為另一種促炎癥細胞因子。如圖2A所示,豬PBMCs在經LPS刺激后,IL-6的分泌較未經LPS刺激時大幅上調(P<0.001)。同樣的,在加入LPS刺激前3小時用鷹嘴豆芽素A預處理細胞則能顯著抑制由LPS刺激引起的IL-6分泌量的增加,終濃度為100μmol/L的鷹嘴豆芽素A(P<0.001)比50 μmol/L的(P<0.01)抑制作用效果更明顯。對IL-6 mRNA表達的研究進一步證實了上述鷹嘴豆芽素A抑制IL-6生成的結果(圖2B)。這些以TNF-α和 IL-6為代表的結果表明鷹嘴豆芽素A能顯著地抑制PBMCs分泌炎性細胞因子。
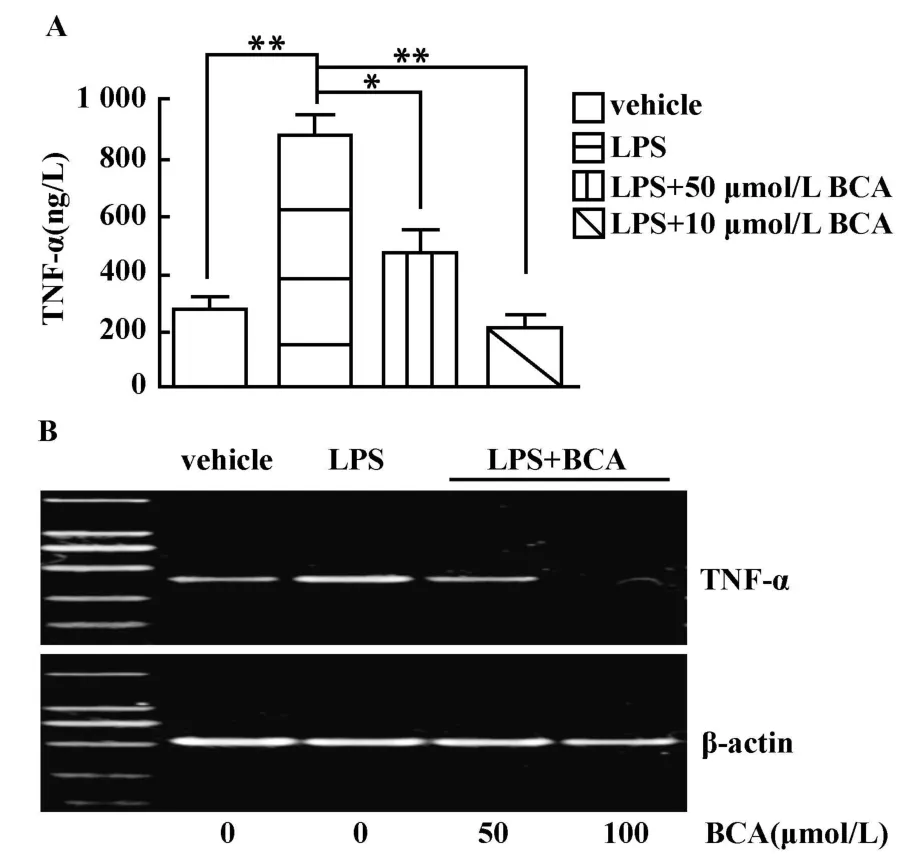
圖1 鷹嘴豆芽素A對LPS刺激豬PBMCs后TNF-α分泌(A)和mRNA表達(B)的影響Fig.1 Effect of biochanin A on the secretion(A)and mRNA expression(B)of TNF-αin porcine PBMCs

圖2 鷹嘴豆芽素A對LPS刺激豬 PBMCs后IL-6分泌(A)和mRNA表達(B)的影響Fig.2 Effect of biochanin A on the secretion(A)and mRNA expression(B)of IL-6 in porcine PBMCs
2.2 鷹嘴豆芽素A對LPS刺激豬PBMCs后NF-κB激活的影響 NF-κB的激活對LPS誘導炎癥因子的釋放具有重要作用。NF-κB在正常情況下以無活性的三聚體形式存在于胞漿中,受刺激活化進入細胞核引起下游多種基因表達,NF-κB通路是LPS誘導炎癥因子表達的關鍵信號通路。可以通過Western blot技術分析NF-κB蛋白在細胞質與細胞核之間的轉移來檢測NF-κB的激活。如圖3所示,在正常情況下豬PBMCs細胞核內僅有少量的NF-κB,在經30分鐘LPS處理后胞漿內NF-κB蛋白含量下降,核內NF-κB含量顯著增加,說明 LPS處理后NF-κB被激活。而在LPS刺激前用鷹嘴豆芽素A預處理3小時,可抑制NF-κB的進核,說明鷹嘴豆芽素A可以抑制LPS誘導的豬PBMCs NF-κB信號途徑的活化。
2.3 鷹嘴豆芽素A通過PPARα與PPARγ途徑抑制LPS刺激豬PBMCs后NF-κB的激活 LPS刺激前用終濃度為100μmol/L的鷹嘴豆芽素A和10μmol/L PPARγ拮抗劑GW9662共同預處理豬PBMCs,這時PPARγ的作用會被GW9662大幅消除。圖4顯示,在PPARγ無法發揮作用的情況下,鷹嘴豆芽素A對LPS誘導的豬PBMCs NF-κB信號途徑活化的抑制作用被顯著減弱;同樣,LPS刺激前用鷹嘴豆芽素A和20μmol/L PPARα拮抗劑 MK886共同預處理豬PBMCs以抑制 PPARα途徑,鷹嘴豆芽素A對LPS誘導的豬PBMCs NF-κB信號途徑活化的抑制作用也被顯著減弱。說明鷹嘴豆芽素A可以通過PPARα和PPARγ途徑抑制 LPS誘導的豬PBMCs NF-κB信號途徑的活化,鷹嘴豆芽素A對LPS刺激PBMCs后炎癥因子的釋放具有抑制作用的原因至少有一部分可歸因于鷹嘴豆芽素A通過PPARα和PPARγ途徑調控NF-κB信號途徑的活化。
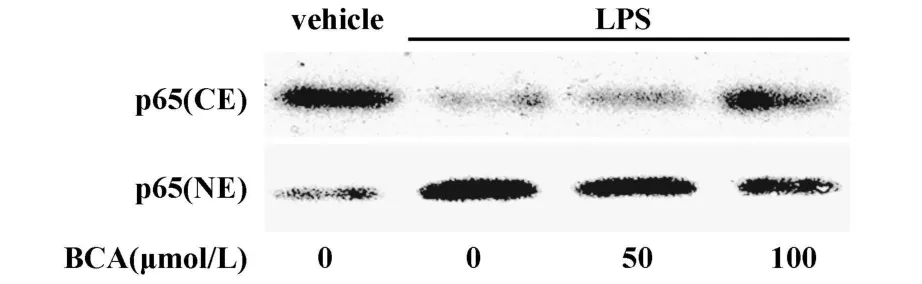
圖3 鷹嘴豆芽素 A對LPS刺激豬PBMCs后NF-κB p65進核激活的影響Fig.3 Effect of biochanin A on the nuclear translocation of NF-κB p65
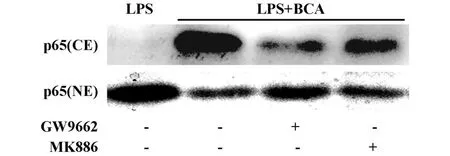
圖4 PPARα或 PPARγ拮抗劑減弱了鷹嘴豆芽素A對LPS誘導豬PBMCs NF-κB p65進核激活的抑制作用Fig.4 PPARαor PPARγantagonist attenuated the suppression effect of biochanin A on the nuclear translocation of NF-κBp65 in LPS-stimulated porcine PBMCs
3 討論
體內體外的一些研究已經證明一些異黃酮具有抗炎作用。在這些異黃酮化合物中,染料木黃酮是被最為廣泛研究的一種,它的抗炎作用在小鼠、人體內和在一些細胞株內被深入探討[1]。大豆素的抗炎癥作用也有過一些報道[1]。然而,對鷹嘴豆芽素A的抗炎癥作用卻很少有報道。本研究報道了鷹嘴豆芽素A具有抑制豬PBMCs經LPS刺激引起TNF-α和IL-6等炎癥因子分泌的作用,這與Muller等[2]在小鼠巨噬細胞RAW264.7內的研究結果一起證實了鷹嘴豆芽素A的抗炎癥作用。
染料木黃酮的抗炎作用機制已被深入地研究,它可以通過抗氧化和清除自由基、抑制COX-2的表達、調控MAPK的表達和調控NF-κB信號途徑來抵抗炎癥反應[7-12]。然而,鷹嘴豆芽素A的抗炎作用機制未見任何報道。本研究報道了鷹嘴豆芽素A可以通過抑制LPS誘導NF-κB信號途徑的活化來抑制炎癥反應,同時,本研究進一步發現了鷹嘴豆芽素A對NF-κB信號途徑的調控是依賴于PPARα和PPARγ這兩種核受體的。有趣的是,刺芒柄花素具有比鷹嘴豆芽素A更高效的PPARα/γ激動劑活性,但是其抑制RAW264.7細胞釋放TNF-α和IL-6的能力卻比鷹嘴豆芽素 A更低[2,4]。更為復雜的是,PPARγ激動劑匹格列酮具有抑制RAW264.7細胞釋放TNF-α的作用而PPARγ另一種激動劑羅格列酮卻沒有這種作用[5,13]。本研究小組在RAW264.7細胞中進行的研究也發現雖然染料木黃酮具有PPARα/γ激動劑活性,但其對炎癥因子釋放的抑制并不依賴于PPARα/γ途徑(數據未展示)。另外,本研究小組還發現在RAW264.7細胞中,鷹嘴豆芽素A的抗炎作用只依賴于PPARγ途徑而不依賴于PPARα途徑(數據未展示),這可能與在RAW264.7細胞中PPARα的表達量很低有關。總之,異黃酮的抗炎癥作用與其PPARα/γ激動劑活性的關系還有待更深入的研究。
1 Garcia-Lafuente A,Guillamon E,Villares A et al.Flavonoids as anti-inflammatory agents:implications in cancer and cardiovascular disease[J].Inflamm Res,2009;58(9):537-552.
2 Mueller M,Hobiger S,Jungbauer A.Red clover extract:a source for substances that activate peroxisome proliferator-activated receptor alpha and ameliorate the cytokine secretion profile of lipopolysaccharide-stimulated macrophages[J].Menopause,2010;17(2):379-387.
3 Kota B P,Huang T H,Roufogalis B D.An overview on biological mechanismsof PPARs[J].Pharmacol Res,2005;51(2):85-94.
4 Shen P,Liu M H,Ng T Y et al.Differential effects of isoflavones,from Astragalus membranaceus and Pueraria thomsonii,on the activation of PPARalpha,PPARgamma,and adipocyte differentiation in vitro[J].J Nutr,2006;136(4):899-905.
5 Cui T,Schopfer F J,Zhang J et al.Nitrated fatty acids:Endogenous antiinflammatory signaling mediators[J].JBiol Chem,2006;281(47):35686-35698.
6 Jones DC,Ding X,Daynes RA.Nuclear receptor peroxisome proliferatoractivated receptor alpha(PPARalpha)is expressed in restingmurine lymphocytes.The PPARalpha in T and B lymphocytes is both transactivation and transrepression competent[J].J Biol Chem,2002;277(9):6838-6845.
7 Khan N,Afaq F,Mukhtar H.Cancer chemoprevention through dietary antioxidants:progress and promise[J].Antioxid Redox Signal,2008;10(3):475-510.
8 Win W,Cao Z,Peng X et al.Different effects of genistein and resveratrol on oxidative DNA damage in vitro[J].Mutat Res,2002;513(1-2):113-120.
9 Horia E,Watkins B A.Complementary actions of docosahexaenoic acid and genistein on COX-2,PGE2 and invasiveness in MDA-MB-231 breast cancer cells[J].Carcinogenesis,2007;28(4):809-815.
10 Hwang J T,Lee Y K,Shin JI et al.Anti-inflammatory and anticarcinogenic effect of genistein alone or in combination with capsaicin in TPA-treated rat mammary glands or mammary cancer cell line[J].Ann N Y Acad Sci,2009;1171:415-420.
11 Davis JN,Kucuk O,Sarkar F H.Genistein inhibits NF-kappa B activation in prostate cancer cells[J].Nutr Cancer,1999;35(2):167-174.
12 Li Y,Sarkar F H.Inhibition of nuclear factor kappaB activation in PC3 cells by genistein is mediated via Akt signaling pathway[J].Clin Cancer Res,2002;8(7):2369-2377.
13 Ao C,Huo Y,Qi L et al.Pioglitazone suppresses the lipopolysaccharideinduced production of inflammatory factors in mousemacrophages by inactivating NF-kappaB[J].Cell Biol Int,2010;34(7):723-730.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
中成藥(2017年4期)2017-05-17 06:09:50
