烤煙品種幼苗低溫反應敏感度與其生化差異性研究
2011-07-31 03:23:16劉少云羅成剛李彥東蘇振剛宋志美
中國煙草科學 2011年5期
馬 倩,常 平,劉少云,熊 萍,羅成剛*,李彥東,劉 魁,蘇振剛,宋志美
(1.國家煙草專賣局煙草遺傳育種重點實驗室,中國農業(yè)科學院煙草研究所,青島 266101;2.中國農業(yè)科學院研究生院,北京 100081;3.山東臨沂煙草有限公司,山東 臨沂 276000;4.山東省煙草公司,濟南 250101;5.山東日照煙草有限公司,山東 日照 276800)
低溫冷害是北方煙區(qū)的主要氣象災害之一,是影響煙草產量和品質的主要限制因子,低溫脅迫使植物體內產生大量自由基,引起膜系統(tǒng)損傷,造成低溫傷害[1]。低溫對烤煙的影響一方面是抑制煙株的生長,另一方面是誘導早花,導致煙葉產量的下降。張燕等[2]研究了在5 ℃低溫脅迫下PEG對煙苗抗冷性生理生化特性的影響,發(fā)現(xiàn)20% PEG浸種處理對提高煙草幼苗膜系統(tǒng)穩(wěn)定性及抗冷性物質含量有一定作用。同時,也發(fā)現(xiàn)CaCl2浸種能夠提高煙草幼苗中結合鈣含量,增強對細胞膜結構的穩(wěn)固作用,減緩低溫對煙草幼苗細胞膜的傷害[3]。陳衛(wèi)國等[4]通過對低溫脅迫下5個烤煙品種一些生理指標的測定,對這5個品種的耐寒差異性進行了研究。本研究選用北方煙區(qū)種植的烤煙品種中煙100、NC82、K326為試驗材料,旨在通過研究低溫條件下烤煙體內脯氨酸含量、膜保護酶含量、丙二醛及相對電導率變化,探討不同烤煙品種的耐寒性生理生化指標的變化規(guī)律,研究不同品種的敏感溫度和時間,為研究防御煙草低溫冷害提供理論依據(jù),為煙草抗寒性育種提供參考。
1 材料與方法
1.1 供試材料
選用烤煙品種中煙 100、NC82、K326為供試材料,種子由中國農業(yè)科學院煙草研究所提供。
1.2 材料處理
試驗于 2009年在中國農業(yè)科學院煙草研究所青島溫室進行。選擇籽粒飽滿的 3個品種種子在28~32℃的溫室播種,自然光照,統(tǒng)一栽培管理。當植株長到五葉一心時進入人工氣候箱進行低溫處理,同時以常溫條件下的幼苗為對照(T)。低溫處理條件和處理時間為:2℃低溫條件下分別培養(yǎng)1 d、2 d、3 d(分別用T2-1、T2-2、T2-3表示);5℃低溫條件下分別培養(yǎng)3 d、5 d、7 d(T5-3、T5-5、T5-7);8℃低溫條件下分別培養(yǎng) 7 d、9 d、11 d(T8-7、T8-9、T8-11)。
1.3 測定方法
低溫處理完成后,各處理分別取生長一致的煙株,對第3片葉、第4片葉、第5片葉進行抗寒指標測定。用DDS-307型電導率儀測定外滲液的相對電導率[5];采用硫代巴比妥酸法測定丙二醛(MDA)含量[6];愈創(chuàng)木酚比色法測定過氧化物酶(POD)含量[7];酸性茚三酮法測定脯氨酸(Pro)含量[6]。所有指標測定均重復3次,取平均值(圖中數(shù)據(jù)為所有葉片各3次重復取樣后測定指標的平均)。
2 結 果
2.1 低溫脅迫對不同烤煙品種相對電導率的影響
電導率的變化能夠反應出低溫脅迫對所測材料的質膜的傷害程度。由圖1可知,在不同低溫處理條件下,3個品種的相對電導率變化趨勢一致,均明顯高于常溫對照。相同脅迫溫度不同脅迫時間,各品種的相對電導率均隨處理時間的增加逐步升高。相同脅迫時間不同脅迫溫度,各品種相對電導率隨溫度降低而增大。在2℃處理3 d時,相對電導率增加程度最明顯,中煙 100、NC82、K326分別較常溫對照增加了81.92%、86.11%、127.20%;在8℃處理7 d時,相對電導率增加程度最小,中煙 100、NC82 、K326分別較常溫對照增加了31.53%、34.79%、59.49%。品種間差異明顯,中煙100相對電導率的變化幅度最小,K326變化幅度最大,NC82居中。
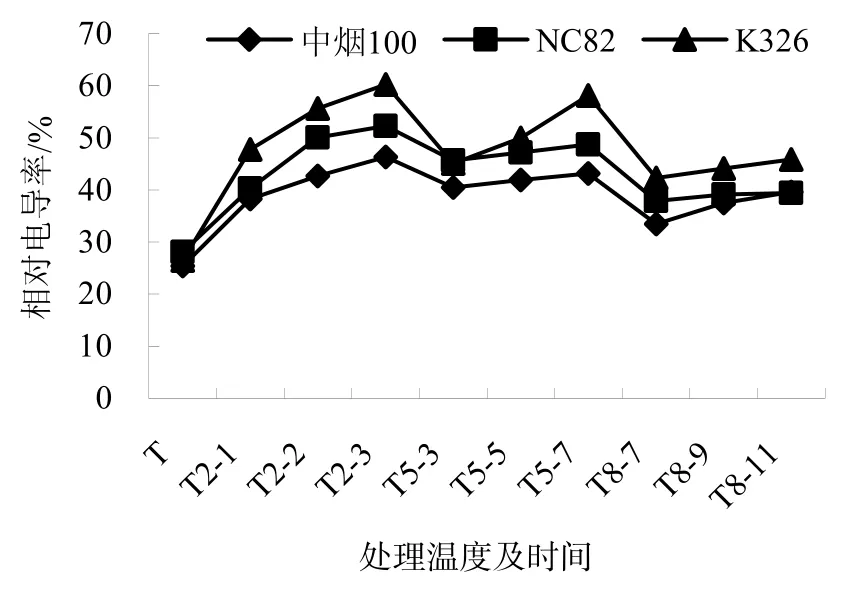
圖1 低溫脅迫下煙草幼苗相對電導率的變化Fig.1 Trends of the relative electrical conductivities of the tobacco seedlings under chilling stress
2.2 低溫脅迫對不同烤煙品種丙二醛(MDA)的影響
MDA是細胞膜脂過氧化的產物,其含量的高低反應了細胞膜脂過氧化水平,結果見圖2。煙草幼苗在低溫脅迫下MDA含量隨脅迫時間的延長逐漸增加,隨脅迫溫度的升高而減小,均高于對照。在對照及不同低溫脅迫下,K326的MDA含量一直高于中煙100和NC82,但3個品種的變化趨勢和幅度相同。3個品種在T2-3、T5-7時,MDA含量達到峰值,另外,中煙100在T5-5、T8-11變化幅度也較大,NC82在 T5-5、T2-2變化較大,K326在T2-2、T2-1變化較大。
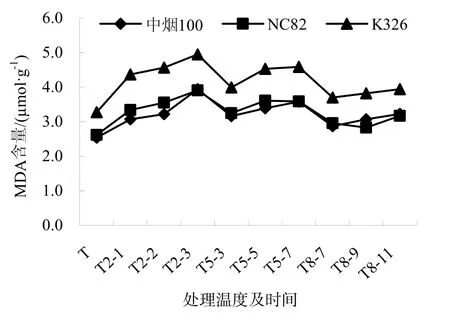
圖2 低溫脅迫下煙草幼苗MDA的變化Fig.2 Trends of MDA contents of the tobacco seedlings under chilling stress
2.3 低溫脅迫對不同烤煙品種過氧化物酶(POD)的影響
POD是植物細胞內清除自由基對細胞膜傷害的重要保護酶之一。在常溫條件下,3個品種的POD活性均保持較低水平。在2 ℃條件下,各品種POD活性均急速上升,中煙100上升幅度最大,且隨處理時間延長,上升幅度逐漸加大;NC82、K326的變化趨勢相同。較之于2 ℃,各品種在5 ℃的POD絕對值降低,變化趨勢相似。而在8 ℃條件下,POD變化趨勢平緩。這說明 POD活性隨低溫脅迫程度的加強而逐漸增大,耐寒性強的品種 POD活性上升幅度大于耐寒性弱的品種。3個品種在2 ℃各個處理和5 ℃培養(yǎng)7 d變化幅度較大,8 ℃變化較小。詳見圖3。
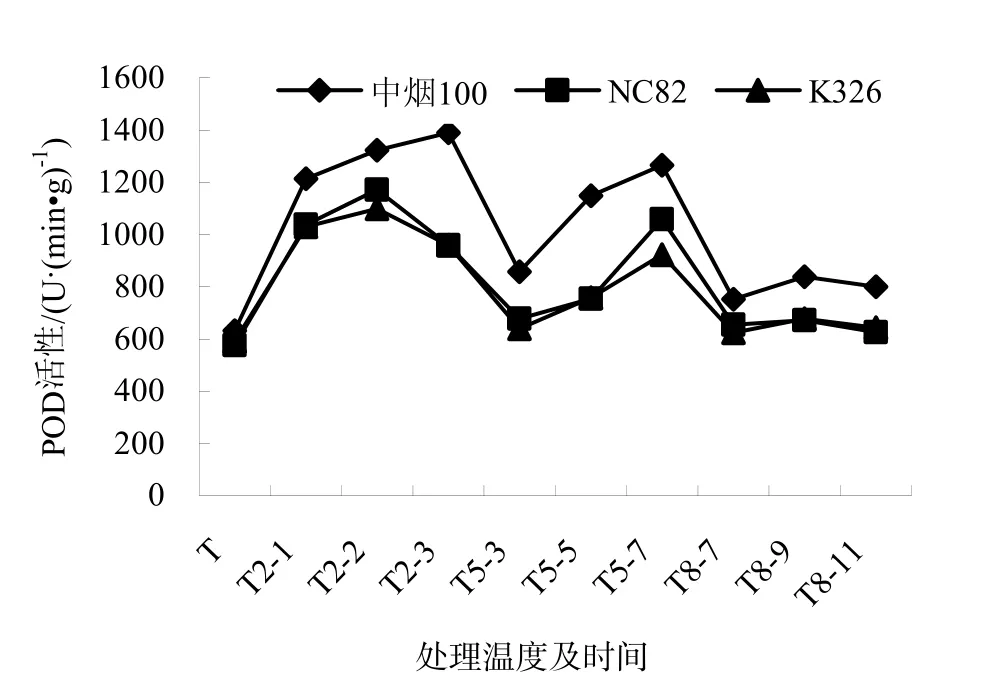
圖3 低溫脅迫下煙草幼苗POD活性的變化Fig.3 Changes of POD activity of the tobacco seedings under chiling stress
2.4 低溫脅迫對不同品種烤煙脯氨酸(Pro)的影響
脯氨酸在植物的低溫脅迫過程中,作為防脫水劑保護植物,同時,其作為一種重要的滲透調節(jié)物質,脯氨酸含量的增加可以有效防止低溫脅迫對植物細胞膜的傷害[8]。由圖4可知,Pro含量隨處理溫度降低而升高,隨脅迫時間的延長而呈增加的趨勢。脅迫溫度的高低和脅迫時間的長短直接影響品種的脯氨酸含量。Pro含量在2 ℃上升速度最快,3個品種均在T2-3處理下達到峰值,中煙100和K326在5 ℃有明顯增加的趨勢,在8 ℃表現(xiàn)較為平緩,而NC82在8 ℃增加明顯。其中,中煙100的Pro隨脅迫處理的變化幅度尤為明顯,處理 T2-3比對照增加了212.91%,T5-7比對照增加了185.28%。K326變化幅度相對平緩,T2-3增加了 147.57%。T5-7增加了129.37%。
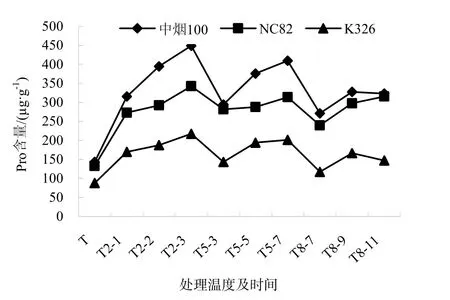
圖4 低溫脅迫下煙草幼苗脯氨酸含量的變化Fig.4 Changes of Pro content of the tobacco seedings under chiling stress
3 討 論
植物的抗寒性是植物體內一系列生理生化過程綜合作用的結果,由相關的多基因調控,并受外界環(huán)境因子的影響或制約。在生理方面,許多研究證明,膜透性、MDA、POD、Pro均可作為鑒定植物耐寒性的指標[9-10]。
生物膜是低溫傷害作用于植物細胞的原初部位,當植物受寒時,其生物膜首先發(fā)生生物相變,生物膜的透性因此增大,細胞內的電解質外滲[11]。因此,可以通過相對電導率的大小來反映植物的抗寒性;加之電導率不受休眠等因素的影響,許多研究者采用此法測定植物在低溫下的受害程度及植物的抗寒力。在本試驗中,根據(jù)電導率的變化,3個品種均在T2-3、T2-2、T5-7、T5-5有低溫敏感性較強的表現(xiàn);電導率增加程度與品種耐寒性強弱相關,耐寒性弱的K326增加幅度最大。
低溫脅迫下,植物產生過多活性氧,引起膜質過氧化作用加劇,MDA含量升高,同時,細胞膜系統(tǒng)因膜質過氧化作用而受損害,導致細胞膜透性增加。從本試驗中可看出,MDA含量變化趨勢與相對電導率相同,隨脅迫程度的增強而增加,3個品種在 2℃和 5℃有最強的低溫敏感性,中煙 100在T8-11也表現(xiàn)出較強的敏感性。
外界的脅迫會導致植物組織內的活性氧超量造成氧化脅迫,從而破壞細胞內的活性氧代謝平衡,活性氧容易引發(fā)膜脂過氧化,造成膜的損傷。植物細胞中存在清除活性氧的保護酶系統(tǒng),能有效地清除活性氧,阻止膜脂過氧化,從而使細胞膜免受傷害[12]。在本試驗中,POD隨低溫脅迫程度的加重呈遞增的趨勢,但在T2-3處理下,NC82和K326增加程度降低,可能隨著脅迫程度加強,植株因低溫造成生理機能的下降和過氧化作用的加劇,膜保護酶系統(tǒng)已不能抵抗逆境傷害,植株的抗逆性降低,損傷加重,葉片干枯。
在逆境條件下,脯氨酸不僅是細胞質中重要的滲透劑和防脫水劑,而且還是活性氧的清除劑,保護膜結構,防止膜脂過氧化。該試驗結果表明,在低溫情況下,根據(jù)3個不同低溫敏感性品種脯氨酸含量的變化,可以看出脯氨酸含量以及變化幅度耐寒性強的品種大于耐寒性弱的品種。3個品種在T2-3有最強低溫敏感性,其次是 T5-7,中煙 100和K326在T5-5、T2-2,NC82在T8-11、T8-9也有明顯變化。
4 結 論
綜合以上各生化指標的變化趨勢可以看出,隨低溫脅迫程度增加,各品種的相對電導率、MDA、脯氨酸、POD均呈增加的趨勢;3個品種均在2 ℃和5 ℃處理下有較強的低溫敏感性,尤其2 ℃條件下培養(yǎng)3 d最強,其次是2 ℃條件下培養(yǎng)2 d及5 ℃條件下培養(yǎng)5 d和7 d,NC82和K326在2 ℃培養(yǎng)3 d時,植株已出現(xiàn)嚴重損傷;3個品種在8 ℃條件下處理7 d時,低溫敏感性最低。3個品種相比較,K326對低溫最敏感,NC82次之,中煙100對低溫最不敏感。
[1]李美如,劉鴻先,王以柔.鈣對水稻幼苗抗冷性的影響[J].植物生理學報,1996,22(4):379-384.
[2]張燕,方立,姚照兵,等.PEG和低溫脅迫對煙草幼苗某些生理特性的影響[J].吉林農業(yè)大學學報,2002,24(6):14-19.
[3]張燕,方立,李天飛,等.鈣對低溫脅迫的煙草幼苗某些酶活性的影響[J].植物學通報,2002,19(3):342-347.
[4]陳衛(wèi)國,李永亮,周冀衡,等.烤煙品種耐寒性及相關生理指標研究[J].中國煙草科學,2008,29(3):39-42.
[5]李錦樹,王洪春,王文英,等.干旱對玉米葉片細胞膜透性及膜脂的影響[J].植物生理學報,1983,4(9):223-227.
[6]朱光廉,鐘誨文,張愛琴.植物生理學實驗[M].北京:北京大學出版社,1990:51-249.
[7]宋風斌,戴俊英.水分脅迫對玉米雌穗生長發(fā)育和產量的影響[J].吉林農業(yè)大學學報,2000,22(1):18-22.
[8]張媛媛,劉明國,趙偉杰.3種外來樹種抗寒性生理指標的比較[J].安徽農業(yè)科學,2007,35(5):1298-1299.
[9]王微.低溫脅迫下紫花苜蓿幾個抗寒生理指標的變化規(guī)律[D].長春:東北師范大學,2009.
[10]張素勤,耿廣東,程智慧.低溫脅迫程度對茄子幼苗生理特性的影響[J].山地農業(yè)生物學報,2008,27(2):119-112.
[11]JYONS J M.Chilling injury in plants[J].Ann Rev Plant Physiol,1973,2(4):445-446.
[12]李明,工根軒.干旱脅迫對甘草幼苗保護酶活性及脂質過氧化作用的影響[J].生態(tài)學報,2002,22(4):503-507.
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫(yī)結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
聲屏世界(2014年6期)2014-02-28 15:18:09
