黃土高原中部沙棘與秋胡頹子光合參數之異同
2011-08-04 03:26:20何正祥劉廣全1王鴻喆
水資源開發與管理 2011年2期
關鍵詞:植物
何正祥,劉廣全1,,3,王鴻喆
(1.中國水利水電科學研究院,北京 100048;2.西北農林科技大學,陜西楊凌 712100;3.國際泥沙研究培訓中心,北京 100048)
沙棘 (Hippophae rhamnoides L.)和秋胡頹子 (Elaeagnus umbellata Thunb.)同為胡頹子科典型植物種,是生態環境脆弱地帶水土保持植被恢復的主要生態經濟型樹種,具有良好的保持水土、防風固沙、改良土壤功能,抗干旱瘠薄、耐鹽堿、抗嚴寒、耐酷暑和適應性強等特點,而且具有較高的經濟價值,為了改善生態環境、應對全球氣候變化、振興區域經濟和實現經濟社會可持續發展,大力發展、開發、利用和保護沙棘及秋胡頹子資源勢在必行。有關沙棘的研究文獻很多,光合作用研究的文獻也較多[1~7],但是有關秋胡頹子的研究文獻較少,所見文獻主要集中在分類、栽培和利用上[8~10],全面系統開展沙棘葉片和秋胡頹子葉片光強度響應曲線、CO2濃度響應曲線、光合速率日變化曲線、熒光動力學曲線、淬滅分析、熒光響應曲線、熒光CO2響應曲線及其參數等比較研究未見報道,深入開展此方面研究意義重大。
1 自然概況
研究地點位于陜西省延安市西北農林科技大學延安樹木園內,該地屬暖溫帶大陸性季風氣候森林草原過渡地帶黃土丘陵溝壑區,年平均氣溫9.4℃,極端最高氣溫39.9℃,極端最低氣溫-28.5℃,≥10℃有效積溫3268.4℃,1月平均氣溫-6.7℃,7月平均氣溫22.9℃。年均降水量550mm,年蒸發量1585mm,年均相對濕度62%,無霜期180d,年日照時數2445h,最大凍土深76cm。土壤以黃綿土為主,母質為黃土,pH值為8.4,土層達100cm以上。天然植被已經不覆存在,多數人工植被發育良好。
2 研究方法
2004年和2005年6~10月在黃土高原中部延安市寶塔區西北農林科技大學延安樹木園,選擇具有代表性20年生的沙棘和秋胡頹子林等為測定對象,用LICOR-6400光合儀對其光合響應曲線、ACI曲線、光合速率、蒸騰速率、氣孔導度和葉綠素熒光動力學曲線、淬滅分析、熒光光曲線以及熒光ACI曲線等光合生理指標進行測定,測定光源分別為紅藍光源和葉綠素熒光葉室光源,每個指標作3個重復,每個重復分別讀數5次。
測定光合響應曲線的光照強度取值分別為:0、20、50、100、200、500、1000、1500 和2000μ.molm-2.s-1;測定ACI曲線時光合有效輻射控制在 1500μ mol?m-2?s-1,CO2濃度取值為:50、100、200、300、400和600μ mol?mol-1;光合作用日進程測定時間為:8:00、10:00、12:00、14:00、16:00和18:00;葉綠素熒光生理指標測定前要確定每個葉片的Fo、Fm、Fv/Fm值,多選在上午6:00~7:00之間測定,測定前采用暗適應,用遮光夾進行20min左右的遮光處理,以保證葉片完全的暗適應。
3 結果與分析
3.1 光強度響應曲線
光強度響應曲線反映植物光合速率隨光照強度增減的變化規律。從沙棘和秋胡頹子光合速率與光輻射強度的關系曲線可以看出,開始光合速率為負值,即光合產物用于植物本身呼吸消耗,隨著光照強度的增加,光合積累與呼吸消耗速率相當,光照強度達到補償點;之后光照強度增加,所測定樹種的光合速率明顯增大,當光照強度達到一定值以后,光合速率基本上穩定在一定水平,即達到光飽和 (圖1和圖2)。
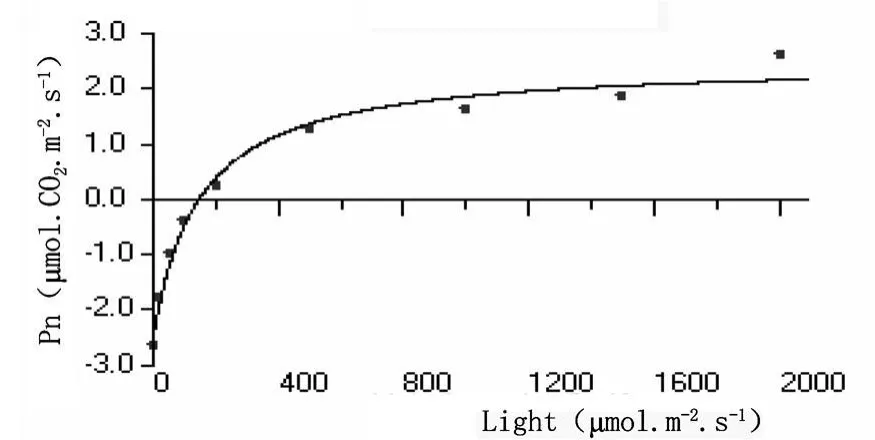
圖1 沙棘葉片光照強度響應曲線
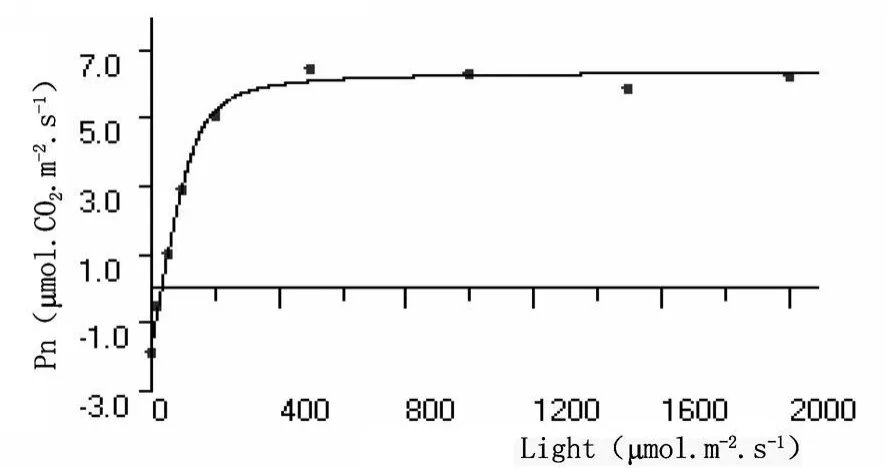
圖2 秋胡頹子葉片光照強度響應曲線
黃土高原中部沙棘葉片和秋胡頹子葉片的暗呼吸速率、表觀量子效率、最大光合速率、光補償點和光飽和點等參數平均值分別為:-3.6770、-1.7150μ mol?CO2?m-2?s-1,0.05169、0.05487mol?mol-1,7.367、8.084μ mol?CO2?m-2?s-1、71.15、31.26μ mol? m-2? s-1和 213.7、178.6μ mol?m-2?s-1。可見不同樹種光強度響應曲線參數值差別較大,沙棘葉片的暗呼吸速率、表觀量子效率和最大光合速率皆小于秋胡頹子葉片的,光補償點和光飽和點相反。
3.2 CO2濃度響應曲線
從圖3和圖4可以看出,沙棘葉片和秋胡頹子葉片光合作用速率均隨CO2濃度的增加而逐漸增大,當植物光合作用吸收的CO2量與呼吸作用和光呼吸釋放的CO2量達到動態平衡時,環境中CO2的濃度為CO2補償點;之后隨著CO2進一步提高,植物光合作用速率迅速上升,幾乎呈線形變化,直線的斜率被稱為羧化效率,其值大小反映相同CO2濃度的光合作用效率的高低。當CO2的濃度升高達到一定值時,光合速率值達到最大,此時的CO2濃度為CO2飽和點,之后隨CO2濃度增加,其光合速率值相對穩定或略有下降,原因可能是CO2濃度過高時引起細胞中pH值的改變,葉片保衛細胞膨壓下降,氣孔開度受到影響,進一步影響凈光合速率。
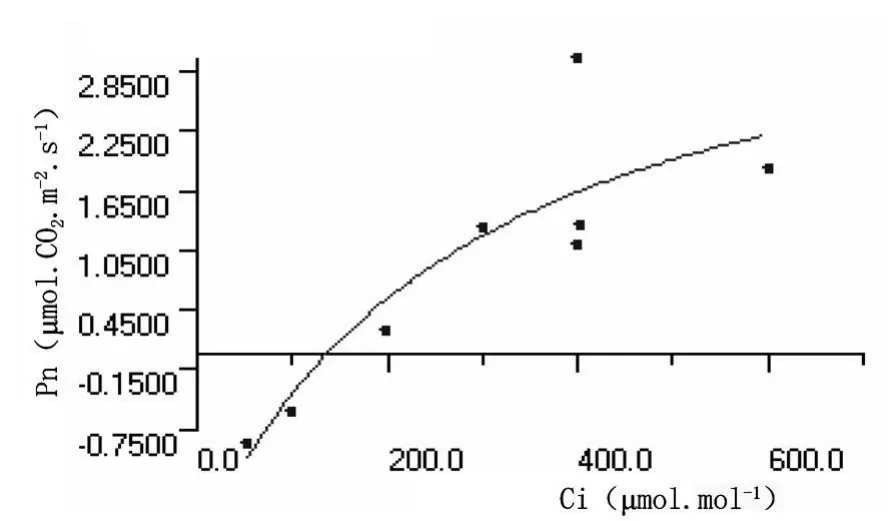
圖3 沙棘葉片CO2濃度響應曲線
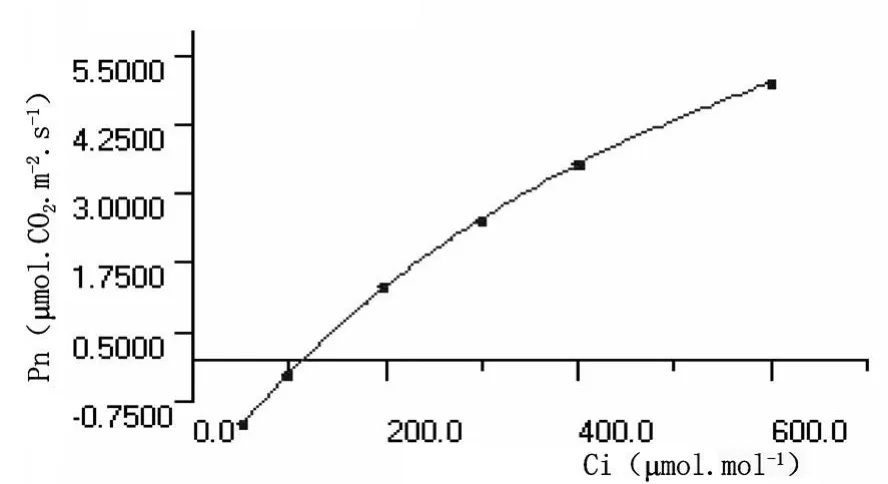
圖4 秋胡頹子葉片CO2濃度響應曲線
測定結果表明,沙棘葉片和秋胡頹子葉片CO2濃度響應曲線參數值分別為:CO2補償點150.00μ mol?mol-1、99 ?00μ mol?mol-1,CO2飽和點506.83370μ mol?mol-1、490.0662μ mol?mol-1,暗呼吸速率 -1.100μ mol?CO2?m-2?s-1、-2.349μ mol? CO2.m-2? s-1, 羧化效率0.00630mol?mol-1、0.02446mol?mol-1,最大光合速率 3.01μ mol? CO2?m-2.s-1、14.80μ mol?CO2?m-2?s-1。秋胡頹子葉片的羧化效率和最大光合速率遠高于沙棘葉片的,暗呼吸速率和CO2補償點卻低于沙棘葉片的,CO2飽和點二者差異不大,可見秋胡頹子對CO2補償反應敏感,且利用率更高。
3.3 光合速率日進程
沙棘葉片和秋胡頹子葉片光合速率 (Pn)日變化都呈現為單峰曲線,峰值出現在10:00~12:00時左右,光合速率日變化從早晨日出起逐漸升高,出現峰值后光合速率逐漸降低。沙棘葉片和秋胡頹子葉片最大凈光合速率平均值分別為16.3547 ±0.7128 、 17.7742 ±0.7245μ mol?CO2?m-2?s-1,二者差別不大;光合速率逐漸降低,之后隨著夜間的來臨其光合速率逐漸降低,沙棘葉片和秋胡頹子葉片最低凈光合速率平均值分別為 3.4806 ±0.7377、9.0465 ±0.5145μ mol?CO2?m-2?s-1,后者是前者的2.6倍 (表1)。
植物的光合速率除了主要受葉片氣孔的調節外,還受葉片所吸收的凈輻射、風速、大氣溫度、空氣濕度、土壤水分等多種因子的影響。葉片氣孔導度 (Gs)的日變化與光合速率的日變化呈現出很好的相關性。在中午 12:00~14:00時,大氣溫度高達最高,從而限制了葉片的光合速率。光合有效輻射 (PAR)是影響植物光合速率大小的主要因素之一,光合有效輻射日變化曲線走勢呈明顯的單峰曲線,即早晚低、中午高,而且峰值出現在中午10:00~12:00之間。植物葉片氣孔限制值大小與其光合速率呈現明顯的負相關關系,但是在中午氣孔限制值達最低時,部分植物光合速率也到達最低值,按照Farquhar和Sharke的觀點,部分植物發生 “午休”現象主要原因是非氣孔因素造成,即葉肉因素對光合速率的限制起主要作用。

表1 沙棘葉片和秋胡頹子葉片光合速率日變化特征
3.4 熒光動力學曲線
植物熒光動力學曲線是研究植物葉片從黑暗條件下轉入光下的活化過程中各個參數的變化。一般而言,暗適應的植物葉片突然照光后,熒光上升,但是光合速率等指標并沒有明顯變化,沒有隨著高光照而表現出高的光合速率,這是由于兩方面的條件限制的:酶活性的激活和氣孔的誘導開放,這時往往氣孔導度很低。電子傳遞速率(ET R-Electron Transport Rate)與植物光合速率有很強的線性關系,是又一個表征植物光合能力高低的變量。從圖5和圖6中可以看出,隨著光照時間的加大,所測定的樹種的ETR逐漸上升并在一定時間后達到穩定,與光合作用速率同步,只是不同樹種達到穩定所需要的時間不同,變化的斜率也有較大的差異。PhiPSⅡ指PSⅡ所吸收的光量子中用于光化學反應的比例,在低光照條件下,PSⅡ的量子產量通常較高,因為葉片所吸收的光能中有較大的比例被用于光化學反應中;而在高光照強度條件下,因為葉片所吸收能量中的很大比例通過非化學過程而散失,所以經過高光照強度光適應的葉片的PhiPSⅡ較低。結果表明,沙棘葉片和秋胡頹子葉片的PhiPSⅡ隨著光照時間逐漸上升,穩定時間與光合同步。
當葉片從黑暗條件轉入光下,PSⅡ反應中心逐漸關閉,這造成葉綠素熒光產量 (1秒鐘之內)的上升,在此之后,熒光產量開始下降,持續大約幾分鐘或幾十分鐘,這就是熒光淬滅。首先,電子被從PSⅡ傳遞走的速率開始上升,這是由于光誘導對C代謝酶的活化和氣孔開放的活化,這種淬滅稱為光化學淬滅 (qP-Proportion of Open PSII);同時,能量轉化為熱能的效率也提高了,這種過程稱為非光化學淬滅(NPQ-Non Photochemical Quenching)。圖 5和圖6的結果表明,在黃土高原中部測定的沙棘葉片和秋胡頹子葉片從黑暗條件轉入光下,qP和NPQ隨著光照時間逐漸上升,達到一定的時間后突然減低,然后逐漸上升,在經過一段時間之后完成淬滅并達到相對穩定的狀態。
3.5 淬滅分析 (三相分離)
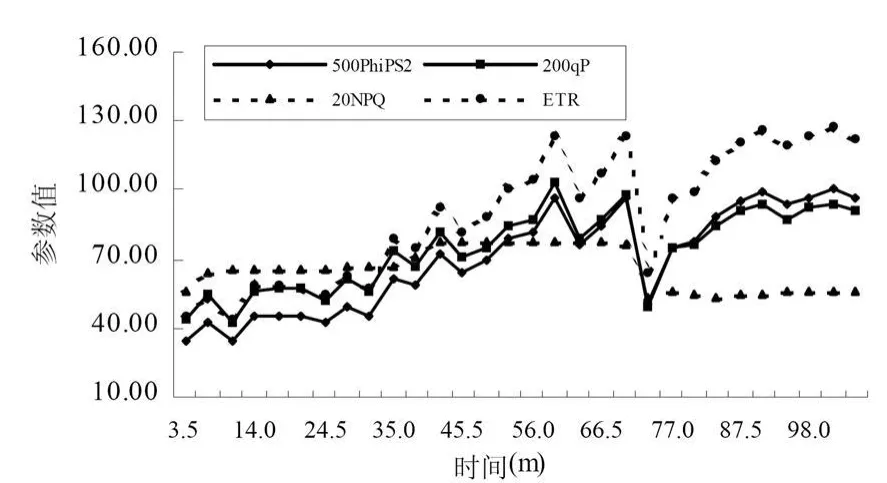
圖5 沙棘葉片熒光動力學曲線參數變化
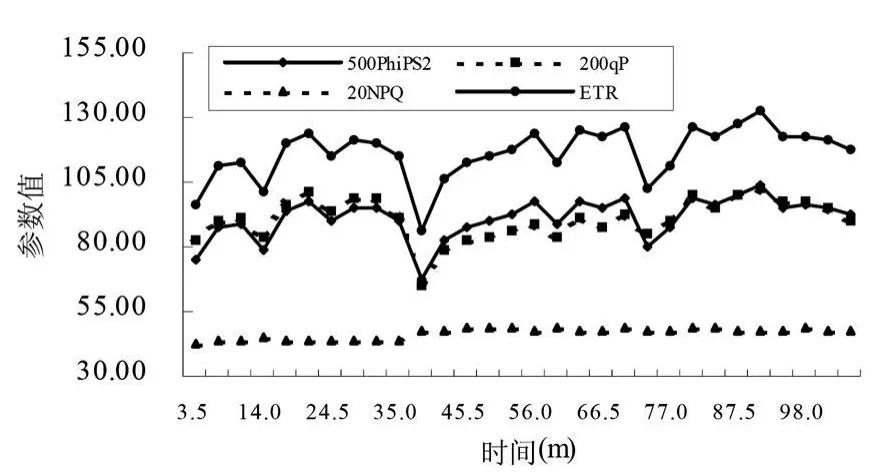
圖6 秋胡頹子葉片熒光動力學曲線參數變化
非光化學淬滅NPQ和熱耗散線性相關,在一個典型的植物中,在飽和光強下大致在0.5~3.5之間。這取決于物種和植物以前的經歷過程。qN是非光化學淬滅的比較傳統的老的術語,有的時候也會被用到。它的范圍在0~1.0之間,因此在淬滅較高時不很敏感。同樣的淬滅在參考點淬滅較高時可能會表現為很小的上升,因此直接的比較不很明確。通常,如果暗適應的Fv/Fm明顯不同,那么NPQ也不能直接進行比較。一般而言,非光化學淬滅的增長可能是由于葉片為免受光破壞的保護機制。研究此過程的一種方法是照光后弛豫的速率。不同的過程有不同的弛豫速率。其時間范圍從幾分鐘到幾小時。
圖7和圖8顯示了沙棘葉片和秋胡頹子葉片在照光后弛豫的速率。就是對光適應的葉片轉入黑暗下之后一個小時之內各種熒光參數的變化動態。從圖中可看出最大熒光產量Fm'(maximal fluorescence during a saturating light flash)在葉片轉入黑暗后逐漸上升,在17min左右趨于穩定。qP(Photochemical quenching)變化不明顯,原因可能是關閉了活化光。qN變化曲線不很明顯,原因可能是在淬滅較高時不很敏感。當光適應的葉片轉入黑暗下之后,NPQ開始下降,在25min左右達到穩定。正常條件下,在利用瞬時光能時,低光照強度適應葉片的光能捕獲效率更高。在低光照強度條件下,沒有多余的光合有效輻射,因此允許大多數植物具有更高的光能捕獲效率。
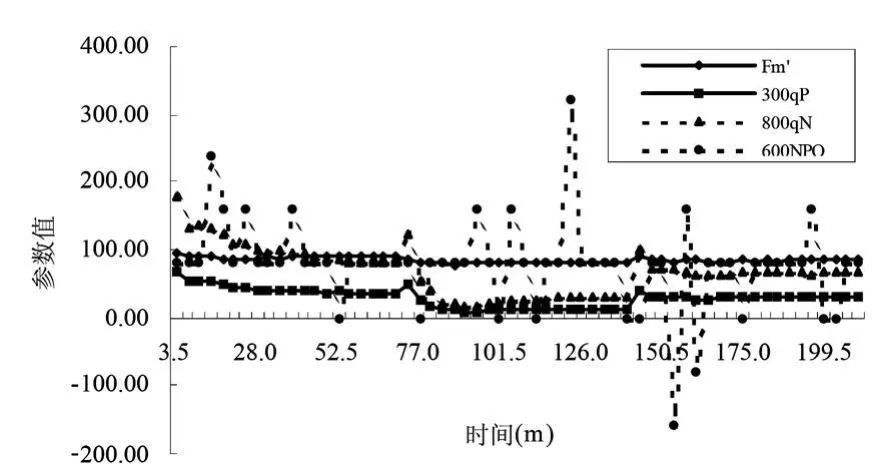
圖7 沙棘葉片熒光淬滅動力學曲線參數變化
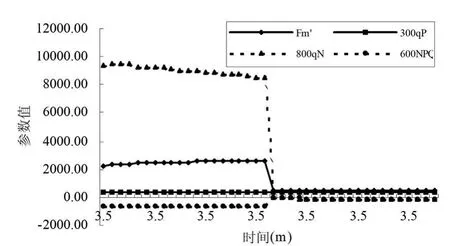
圖8 秋胡頹子葉片熒光淬滅動力學曲線參數變化
3.6 熒光光強響應曲線
植物熒光光照強度響應曲線與植物普通光強度響應曲線的變化規律類似(圖9和圖10),熒光光照強度為零時,植物光合速率為負值,光合產物用于植物本身呼吸消耗,隨著光照強度的增加,光合積累與呼吸消耗速率相當,光照強度達到補償點 (LCP);之后光照強度增加,植物光合速率明顯增大,當光照強度達到一定值以后,光合速率基本上穩定在一定水平,即達到光飽和(LSP)。
沙棘葉片和秋胡頹子葉片熒光響應曲線參數暗呼吸速率、光的補償點、光的飽和點、最大光合速率和表觀量子效率其值分別為-0.5106μ mol?CO2?m-2?s-1、-1.3670μ mol?CO2?m-2?s-1,14.72μ mol?m-2?s-1、 28.94μ mol ? m-2? s-1,259.6μ mol? m-2? s-1、321.3μ mol ? m-2? s-1,8.494μ mol?CO2?m-2?s-1、 13.810μ mol?CO2?m-2?s-1和 0.03469mol?mol-1、0.04723mol?mol-1,植物葉的熒光光響應參數與光合作用參數一樣,主要是植物的遺傳特性所決定,不同植物種類存在一定的差異。
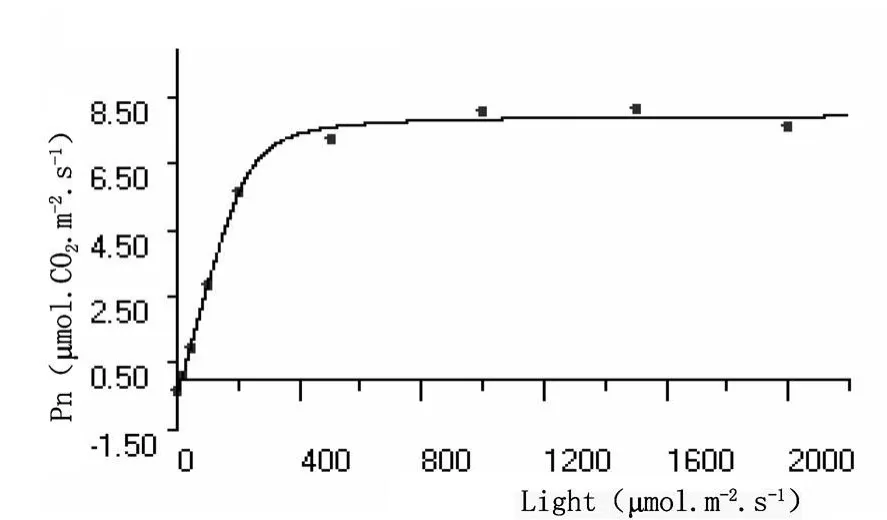
圖9 沙棘葉片熒光光強響應曲線
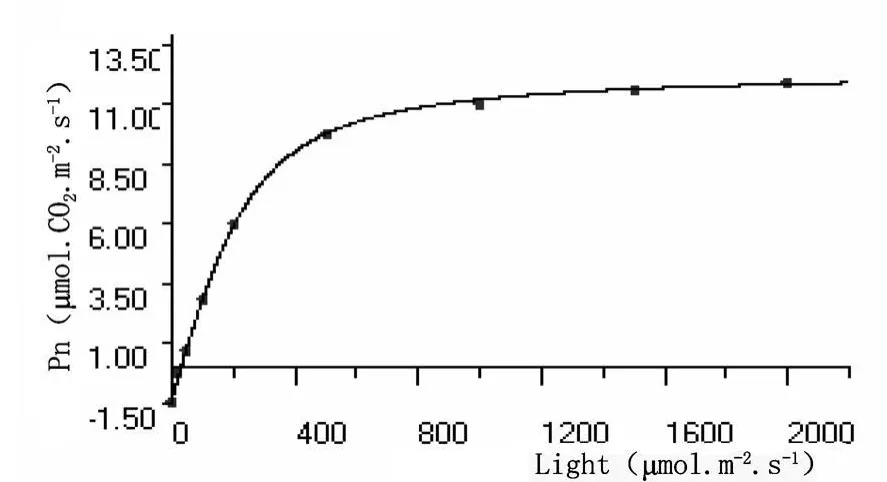
圖10 秋胡頹子葉片熒光光強響應曲線
在植物熒光光照強度響應曲線的實驗中,將從一個光適應的植物開始,通過減少瞬時光照強度,目的是為了逐漸得到更高的量子效率和量子產量。PhiPsⅡ指通過熒光計算的PSⅡ量子產量,從圖中看出,隨著光照強度的減弱PhiPsⅡ逐漸增高,說明葉片吸收的光能中用于光化學反應的比例逐漸增大。在一定的程度上,隨著光強度的逐漸降低,PhiPsⅡ值幾乎呈直線上升,光強度拐點值一般在 200~500μ mol?m-2?s-1之間,不同樹種存在一定的差異;達到該值之后,光強度逐漸降低,PhiPsⅡ緩慢上升。PhiCO2是通過CO2同化作用計算的光量子產量,光強度在2000~25μ mol?m-2?s-1之間,樹木葉的PhiCO2呈緩慢上升趨勢,當光強度降到25μ mol?m-2?s-1左右時,PhiCO2值突然下降或上升。樹種葉子光強度在 2000~25μ mol?m-2?s-1之間降低時,qP 值與光強呈顯著的線形負相關關系;而qN、NPQ和ET R值與光強呈的線形正相關關系 (圖11和圖12)。
3.7 熒光C O2響應 (ACI)曲線
木本植物熒光CO2強度響應曲線與普通光源CO2強度響應曲線變化規律相似,即植物光合作用速率均隨CO2濃度的增加而逐漸增大,當植物光合作用吸收的CO2量與呼吸作用和光呼吸釋放的CO2量達到動態平衡時;之后隨著CO2進一步提高,植物光合作用速率迅速上升,幾乎呈線形變化,其值大小反映相同CO2濃度的光合作用效率的高低。當CO2的濃度升高達到一定值時,光合速率值達到最大,之后隨CO2濃度增加,其光合速率值相對穩定或略有下降(圖13和圖14)。
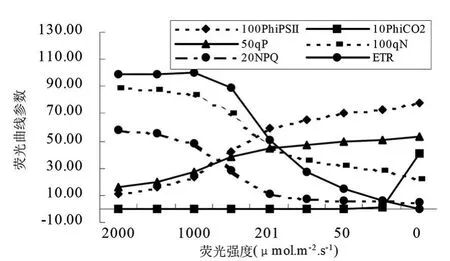
圖11 沙棘葉片熒光光強響應曲線參數變化
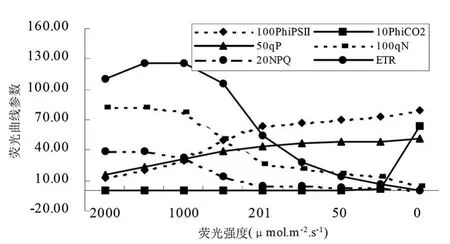
圖12 秋胡頹子葉片熒光光強響應曲線參數變化
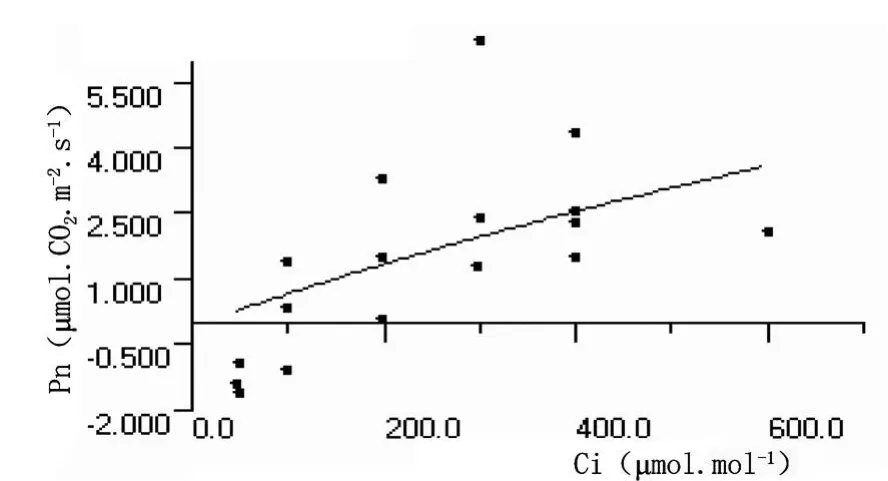
圖13 沙棘葉片熒光CO2響應曲線
沙棘葉片和秋胡頹子葉片的熒光CO2響應曲線參數分別為:暗呼吸速率、CO2補償點、CO2飽和點、最大光合速率以及羧化效率數值分別為 -0.0914、-1.0180μ mol?CO2?m-2?s-1,87.12、 117.80μ mol ? mol-1, 745.2123、832.7138μ mol ? mol-1, 17.90、22.40μ mol?CO2?m-2?s-1和 0.00769、0.01292mol?mol-1。
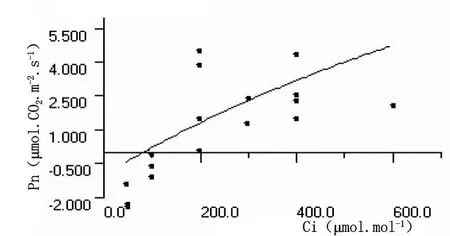
圖14 秋胡頹子葉片熒光CO2響應曲線
沙棘葉片和秋胡頹子葉片熒光ACI曲線參數PhiPsⅡ、PhiCO2、qP、qN、NPQ和 ET R變化中,隨著CO2濃度的降低qN的值保持一個相對穩定的數值,NPQ呈現逐漸上升的趨勢;其它參數PhiPsⅡ、PhiCO2、qP和ETR值隨著CO2濃度的降低而降低,并且彼此呈顯較好線形正相關關系 (圖15和圖16)。

圖15 沙棘葉熒光ACI曲線參數關系
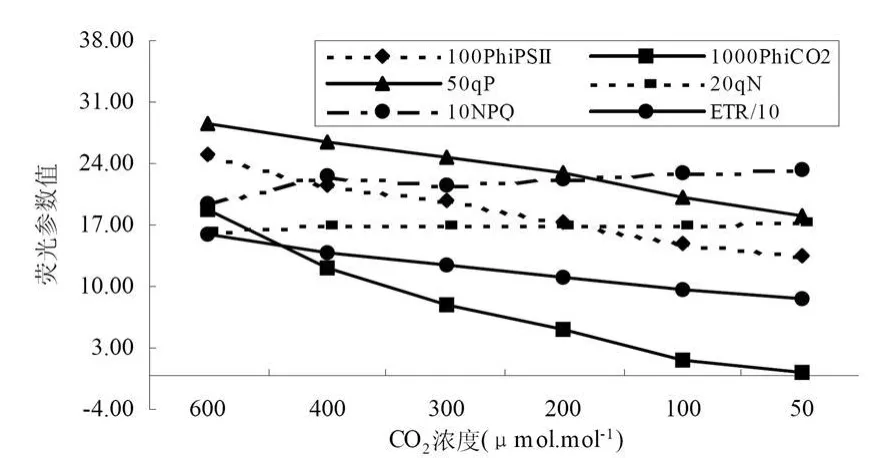
圖16 秋胡頹子葉熒光ACI曲線參數關系
4 小結
(1)沙棘葉片和秋胡頹子葉片開始光合速率為負值,即光合產物用于植物本身的呼吸,隨著光強的增加,光合速率與呼吸速率相當,之后光強增加光合速率明顯增大,當光照強度達到一定值以后,光合速率基本上穩定在一定水平;二者光強響應曲線參數暗呼吸速率、表觀量子效率、最大光合速率、光補償點和光飽和點平均值分別為:-3.6770μ mol ? CO2? m-2? s-1、-1.7150μ mol?CO2?m-2?s-1,0.05169mol? mol-1、0.05487mol?mol-1,7.367μ mol?CO2?m-2?s-1、8.084μ mol?CO2?m-2?s-1、71.15μ mol?m-2?s-1、31.26μ mol?m-2? s-1和 213.7μ mol?m-2? s-1、178.6μ mol?m-2?s-1。
(2)沙棘葉片和秋胡頹子葉片光合速率隨CO2濃度的增加而逐漸增大,當植物光合吸收的CO2量與呼吸和光呼吸釋放的CO2量達到動態平衡時,之后隨著CO2濃度提高,植物光合速率迅速上升,幾乎呈線形變化,延續到光合速率值達到最大,再后光合速率相對穩定或略有下降。測定二者葉片CO2濃度響應曲線參數CO2補償點,CO2飽和點,暗呼吸速率,羧化效率及最大光合速率分別為 CO2補償點150.00μ mol?mol-1、 99.00μ mol ? mol-1,506.83370μ mol?mol-1、 490.0662μ mol?mol-1,-1.100μ mol ? CO2? m-2? s-1、-2.349μ mol?CO2?m-2?s-1,0.00630mol ? mol-1、0.02446mol?mol-1,3.01μ mol?CO2?m-2?s-1、14.80μ mol?CO2?m-2?s-1。
(3)沙棘葉片和秋胡頹子葉片光合速率日變化為單峰曲線,其峰值出現在10:00~12:00時左右,最大值分別為 16.3547±0.7128、17.7742±0.7245 μ mol?CO2?m-2?s-1, 最低值分別為 3.4806±0.7377、9.0465±0.5145μ mol?CO2?m-2?s-1;實際上,植物的光合速率除了主要受葉片氣孔的調節外,還受葉片所吸收的凈輻射、風速、大氣溫度、空氣濕度、土壤水分等多種因子的影響。
(4)隨著光照時間的加大,沙棘葉片和秋胡頹子葉片的ET R逐漸上升并在一定時間后達到穩定,與光合作用速率同步。沙棘葉片從黑暗條件轉入光時,qP和NPQ隨著光照時間逐漸上升,達到一定的時間后突然減低,然后逐漸上升,在經過一段時間之后完成淬滅并達到相對穩定的狀態。
(5)在照光后弛豫的速率,就是對光適應的葉片轉入黑暗下之后一個小時之內各種熒光參數的變化動態。沙棘葉片和秋胡頹子葉片最大熒光產量Fm'在葉片轉入黑暗后逐漸上升,在17min左右趨于穩定;qP和qN變化不明顯;當光適應的葉片轉入黑暗下之后,NPQ開始下降,25min左右達到穩定。
(6)沙棘葉片和秋胡頹子葉片熒光強度響應曲線與其普通光強響應曲線的變化規律類似,熒光響應曲線參數暗呼吸速率、光的補償點、光的飽和點、最大光合速率和表觀量子效率其值分別為 -0.5106μ mol? CO2? m-2? s-1、-1.3670μ mol?CO2?m-2?s-1,14.72μ mol?m-2?s-1、28.94μ mol?m-2?s-1,259.6μ mol?m-2?s-1、321.3μ mol?m-2?s-1,8.494μ mol?CO2? m-2?s-1、13.810μ mol?CO2?m-2?s-1和 0.03469mol? mol-1、0.04723mol?mol-1。
(7)沙棘葉片和秋胡頹子葉片熒光CO2響應曲線與普通光源CO2響應曲線變化規律相似,熒光CO2響應曲線參數暗呼吸速率、CO2補償點、CO2飽和點、最大光合速率以及羧化效率分別為 -0.0914μ mol?CO2?m-2? s-1、-1.0180μ mol?CO2?m-2?s-1,87.12μ mol ? mol-1、117.80μ mol ? mol-1,745.2123μ mol ? mol-1、832.7138μ mol?mol-1,17.90μ mol?CO2?m-2?s-1、22.40μ mol?CO2?m-2?s-1和0.00769mol?mol-1、0.01292mol?mol-1。
[1] 阮成江,李代瓊.半干旱黃土丘陵區沙棘的光合特性及其影響因子[J].植物資源與環境學報,2000,9(1):16-21.
[2] 劉瑞香,楊吉力,高麗.不同土壤水分條件對中國沙棘和俄羅斯沙棘的光合和蒸騰作用的影響[J].內蒙古大學學報(自然科學版),2006,37(2):163-180.
[3] 蔣高明,林光輝.幾種荒漠植物與熱帶雨林植物在不同CO2濃度下光合作用對光照強度的反應[J].植物學報,1996,39(12):972-991.
[4] 孟函寧,劉明國,劉青柏,等.阜新地區不同沙棘品種光合及蒸騰特性的研究 [J].沈陽農業大學學報,2007,38(3):345-348.
[5] 李文華,朱清科,賴亞飛,等.陜北黃土高原丘陵溝壑區沙棘的光合特性研究 [J].干旱地區農業研究,2007,25(1):135-139.
[6] 張瑋,李曉燕,李連國,等.內蒙古西部地區沙棘光合與蒸騰特性研究[J].華北農學報,2007,22(3):97-100.
[7] 唐道鋒,賀康寧,朱艷艷,等.白榆沙棘光合生理參數與土壤含水量關系研究 [J].水土保持研究,2007,14(1):230-233.
[8] 柴發熹.牛奶子營養成分及開發利用途徑[J].林業科技開發.1995(1):50.
[9] 徐曉丹,鄭偉,鐘曉紅.胡頹子屬種質資源開發利用研究進展 [J].湖南農業科學,2007,(5):44-46.
[10] 朱篤,徐曲.胡頹子果實營養成分的測定[J].江西師范大學學報 (自然科學版),2000,24(1):90-91.
[11] 武麗君,畢潤成.不同生境傘花胡頹子光合生理特征的比較[J].山西師范大學學報 (自然科學版),2009,23(2):78-82.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
