大劣按蚊絲氨酸蛋白酶AdSP3的組織定位和定量研究
2011-08-06 11:42:16王英黃復生張錫林徐文岳段建華
成都醫學院學報 2011年2期
關鍵詞:水平
王英,黃復生,張錫林,徐文岳,段建華
(第三軍醫大學基礎部病原生物學教研室,重慶 400038)
明確某一分子在機體中不同組織的表達、分布及轉錄水平,對該蛋白的功能研究具有重要提示意義。昆蟲能夠通過其天然免疫機制防御病原體的入侵感染,其免疫系統包括物理屏障作用、細胞免疫反應和體液免疫反應等。其中,絲氨酸蛋白酶級聯反應能夠調節昆蟲的各種免疫反應,尤其在昆蟲抗入侵病原體的黑化包被反應中起著關鍵作用[1]。絲氨酸蛋白酶能夠催化無活性的PPO蛋白水解成有活性的PO,進而啟動黑色素的產生和沉積,同時促進傷口愈合和包被反應的發生[2,3]。絲氨酸蛋白酶還參與了細胞吞噬作用,因為利用PPO抗體或PAPs抗體能夠明顯阻斷對細菌的細胞吞噬作用;而分裂素活化蛋白激酶能夠通過調節PAPs的分泌控制黑化包被反應和細胞吞噬作用[4]。本研究利用 RTPCR技術和Real-time PCR技術,從mRNA水平研究了絲氨酸蛋白酶3(AdSP3)在大劣按蚊不同組織中的分布及表達水平與瘧原蟲感染的相關性,推測該酶在大劣按蚊抗約氏瘧原蟲感染中的生物學作用。
1 材料與方法
1.1 材料
大劣按蚊(Anopheles dirus)為我國海南株,約氏瘧原蟲(Plasmodium yoelii)為BY265株,感染所用小鼠為昆明株小鼠。
1.2 主要試劑
DEPC購自Sigma公司,瓊脂糖購自OXOID公司,溴化乙錠和T ripure RNA提取液購自 Roch公司,RT-PCR試劑盒、DNA Marker及DNA膠純化回收試劑盒購自 Takara公司,Taq酶購自Promega公司,SYBR Green I購自天澤基因公司。
2 方法
2.1 PCR方法對蚊體內絲氨酸蛋白酶的表達進行組織定位
2.1.1 標本采集及總RNA的提取 取正常4日齡大劣按蚊雌性成蚊200只,采用擠壓法(方法同前)將血淋巴細胞收集入1ml Tripure RNA提取液中,用微量加樣器槍頭吸打數次。取正常4日齡大劣按蚊雌成蚊20只,解剖獲得蚊胃。取正常4日齡大劣按蚊雌成蚊30只,解剖獲得蚊唾液腺。將采集的蚊胃和唾液腺分別在1 ml的Tripure RNA提取液中進行勻漿。常規方法提取總RNA,并進行核酸定量及純度檢測。
2.1.2 反轉錄(RT)合成cDNA 加樣Total RNA(6 μ l)、5 ×Buffer(4 μ l)、MMLV 反轉錄酶(1 μ l)、dNTP(4 μ l)、Oligo(dT)15(1 μ l)、RNAi(1 μ l)、DEPC 處理水(3 μ l)。混勻后,42℃孵育2 h,99℃5 min滅活 MMLV,備用。
2.1.3 聚合酶鏈式反應(PCR)引物設計與合成:根據本課題組已合成的AdSP3的cDNA序列,利用Primer軟件設計引物序列,委托上海生工生物工程技術服務有限公司合成引物。上游引物序列(5′to3′)CAA CAA GAG GCT GGA TGA AGG,下游引物序列(5′to3′)TGC TGC TGG T TC TCG 。反應體系為 cDNA(3.0 μ l)、10×Buffer(2.5 μ l)、5 u/μ l的 Taq 酶(0.3 μ l)、25mM 的 MgCl2(1.5 μ l)、2.5 mM 的 dNTP(2.0 μ l)AdSP3 上游引物 1.0 μ l、Ad-SP3下游引物(1.0 μ l)、ddH2O(13.7 μ l)。反應條件為94℃預變性5 min;94℃變性1 min,55℃退火1 min,72℃延伸1 min,共 35個循環,最后 72℃再延伸5 min。1%瓊脂糖凝膠電泳檢測PCR產物。
2.2 大劣按蚊感染約氏瘧原蟲前后及不同時相點絲氨酸蛋白酶的轉錄水平研究
2.2.1 動物感染與實驗分組 設感染組和正常組,分別吸食感染約氏瘧原蟲的鼠血和正常鼠血。大劣按蚊感染約氏瘧原蟲的方法同前。分別于吸血后1 d、3 d、5 d、7 d和 9 d采集大劣按蚊雌蚊的血淋巴(吸感染血組依次為 I1d 、I3d、I5d、I7d、I9d 和吸正常血組N1d),直接虹吸入 Tripure RNA提取液中。總RNA的提取及反轉錄方法同上。
2.2.2 Real-time PCR 引物同上述 RT-PCR,利用SYBR Green染色法。對以上分組的每組均分別進行AdSP3和S7(內參照)的擴增,另設AdSP3和S7擴增的空白對照管(只加引物,不加模板)。此外,同時以AdSP1的質粒為模板做一標準曲線。反應體系 :cDNA 2 μ l、10 ×Buffer 2.5 μ l、5 u/μ l的Taq 酶 0.3 μ l、25 mM 的 MgCl21.5 μ l、2.5 mM 的dNTP 2 μ l、AdSP3上游引物 1 μ l、AdSP3下游引物1 μ l、SYBR Green(1 ∶10000 稀釋液)1 μ l、ddH2O 13.7 μ l。反應條件:94℃預變性5 min后,94℃變性1 min、55℃退火1 min、72℃延伸1 min,讀板,共35個循環。
3 結果
3.1 絲氨酸蛋白酶在正常大劣按蚊血淋巴細胞、唾液腺和蚊胃中的分布情況
經過RT-PCR及瓊脂糖凝膠電泳后,在紫外儀內可見血淋巴細胞組(L)和唾液腺組(S)均有一清晰而銳利的420 bp條帶,這與根據AdSP3序列預測大小一致。蚊胃組(G)則見多個條帶(見圖1)。
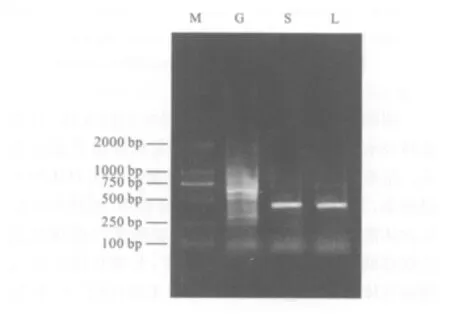
圖1 1%瓊脂糖凝膠電泳檢測RT-PCR產物Fig.1 Result of 1%agarose electrophoresis to test product of RT-PCR
3.2 大劣按蚊感染約氏瘧原蟲前后及不同時相點絲氨酸蛋白酶的轉錄水平
Real-time PCR結果中的熔解曲線圖(見圖2)顯示,除主峰外未見雜峰,擴增AdSP3的 Tm值(熔解溫度)為80℃,擴增S7的 Tm 值為79℃,說明擴增效果好,特異性強(無非特異擴增)。因此,用于定量研究結果可信。在對AdSP3和S7的擴增中,熒光檢測顯示起跳好,說明擴增成功(見圖3)。
根據檢測結果和標準曲線計算出各試驗組的AdSP3和S7的模板含量(拷貝數),然后求得同組的Ad*SP3與S7模板含量比值(見表1),此比值即可反映各試驗組間AdSP3的相對轉錄水平(即mRNA水平)。結果顯示,I7d與I9d兩組間無顯著性差異(P>0.05),其余各試驗組間的AdSP3轉錄水平比較均具有顯著性差異(P<0.01)。I1d組AdSP3的轉錄水平明顯高于同時相點的N1d組;I3d組則水平下降,低于I1d組;I5d組轉錄水平又明顯上升;I7d組AdSP3的轉錄水平又明顯回降,I9d組轉錄水平與I7d組相當,趨于穩定表達水平(見圖4)。
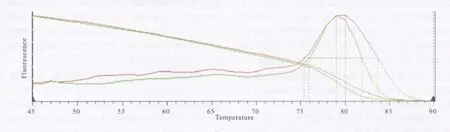
圖2 AdSP3和S7的熔解曲線Fig.2 Melting curve of AdSP3 and S7

圖3 AdSP3的Real-time PCR擴增結果Fig.3 Results of Real-time PCR to amplify AdSP3
表1 Real-time PCR結果中各試驗組的轉錄水平(n=6,±s)Tab.1 Transcription level of groups shown in the results of Real-time PCR(n=6,±s)

表1 Real-time PCR結果中各試驗組的轉錄水平(n=6,±s)Tab.1 Transcription level of groups shown in the results of Real-time PCR(n=6,±s)
?
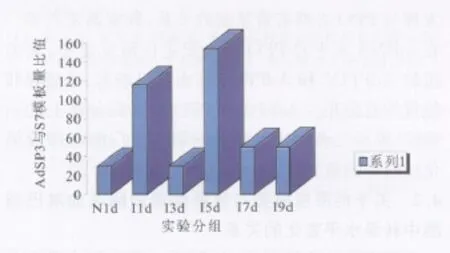
圖4 吸血后不同時相點大劣按蚊各組AdSP3的轉錄水平Fig.4 Transcriptions of AdSP3 in An.dirus at different time points after blood feeding
4 討論
4.1 關于絲氨酸蛋白酶在蚊體內的組織定位
本研究首先利用RT-PCR方法,從mRNA水平檢測了AdSP3在大劣按蚊不同組織的分布情況。由結果看,從血淋巴細胞和唾液腺中均能成功擴增出特異條帶,大小420 bp,與根據Genbank中的AdSP3序列及引物序列預測的目的片段大小完全一致。本研究結果提示,在血淋巴細胞和唾液腺中存在絲氨酸蛋白酶AdSP3的mRNA,即蚊血淋巴細胞和唾液腺能夠成功表達絲氨酸蛋白酶。而在蚊胃中對AdSP3擴增,出現多個條帶,根據這一結果我們暫無法確定在蚊胃中有無AdSP3的轉錄。
對煙草天蛾組織定位研究顯示,不同的絲氨酸蛋白酶表達部位并不完全一致:HP13和 HP18只在血淋巴細胞內表達,而HP12和HP20-HP22則僅在脂肪體細胞表達,其它的HPs則在這兩種細胞內均有表達[5]。對岡比亞按蚊的研究發現,其絲氨酸蛋白酶Sp22D主要表達于血淋巴細胞、脂肪體細胞和蚊胃上皮細胞,且蚊胃組織的陽性信號最強[6]。提示大劣按蚊的各種絲氨酸蛋白酶表達部位可能也不完全一樣。本研究結果僅提示AdSP3在血淋巴細胞和唾液腺內有表達,在蚊胃內則不確定。
絲氨酸蛋白酶是黑化包被反應中的一個關鍵酶,它能夠催化無活性的PPO蛋白水解成有活性的PO,進而啟動黑化反應的發生。而黑化包被反應是昆蟲天然免疫的重要機制之一,也是蚊抗瘧原蟲感染的重要方式。絲氨酸蛋白酶在黑化反應中作用的發揮與PPO必然有著緊密的關系,我室郝宏興等[7]在mRNA水平對PPO的組織定位研究顯示,大劣按蚊AdPPO2和AdPPO3在血淋巴細胞、唾液腺和蚊胃均有分布。AdSP3與PPO在組織分布上的一致性,提示二者的相關性及絲氨酸蛋白酶在卵囊黑化反應中的重要作用。
4.2 關于瘧原蟲感染與絲氨酸蛋白酶在血淋巴細胞中轉錄水平變化的關系
根據熔解曲線可見,在PCR過程中基本沒有非特異擴增和引物二聚體的產生,使得試驗結果能較好地反映擴增的目的片段含量,從而準確計算出模板中的cDNA含量。本研究結果顯示,吸食感染血后1 d的按蚊血淋巴AdSP3的轉錄水平明顯高于同時相點的吸食正常血組。這與Volz等[8]的研究結果一致,即感染后1 d時血淋巴中絲氨酸蛋白酶的含量較吸正常血組增加。瘧原蟲感染按蚊后24 h,正是動合子穿過蚊胃壁的過程,這是瘧原蟲感染按蚊蚊胃并形成卵囊的時期,會啟動一系列的蚊天然免疫[9]。事實上,昆蟲的各種絲氨酸蛋白酶在感染后24 h的轉錄水平變化并不完全一樣。例如煙草天蛾在細菌攻擊感染后 24 h時血淋巴細胞內PAP2和血淋巴蛋白 2(HP2)、HP7、HP9、HP10、HP12-HP22的 mRNA水平均增高,血漿中HP12、HP14-HP19、HP21和 HP22等蛋白的濃度在細菌攻擊感染后24 h也增加;而HP24感染后24 h卻幾乎未見表達。岡比亞按蚊在進行細菌或瘧原蟲攻擊感染后,其絲氨酸蛋白酶Sp14A、Sp14D1、Sp14D2和Sp22D的轉錄均上調,但Sp18D在攻擊感染后mRNA水平則未見增高[6],因此,不同的絲氨酸蛋白酶受到免疫信號的不同調節。總之,在昆蟲體內絲氨酸蛋白酶組成了一個復雜的蛋白酶網絡,介導著對創傷和病原體感染的快速防御反應[5,10]。本結果說明,瘧原蟲的感染誘導了按蚊AdSP3的表達增強,進一步支持了絲氨酸蛋白酶在蚊抗瘧原蟲感染中的作用。
根據吸血后不同時相點的定量檢測,得知吸血后3 d大劣按蚊的AdSP3 mRNA水平明顯下降,這可能由兩個原因造成:一是感染后24 h時大量mRNA參與了AdSP3的翻譯、表達,對mRNA的消耗過大;二是感染后3 d時瘧原蟲已穿過蚊胃壁,在胃基底膜處定位后,卵囊逐漸發育(這一階段相對較為穩定),對按蚊的刺激因子較少,以使得按蚊對Ad-SP3的轉錄降低。而蚊感染瘧原蟲后5 d時,Ad-SP3的表達又明顯增加,提示感染后5 d時又大量啟動了絲氨酸蛋白酶的轉錄、表達,以增強黑化反應的發生。我室徐文岳等[11]的研究發現,大劣按蚊對約氏瘧原蟲的卵囊黑化在光鏡下最早于感染后7 d時發現,因此黑化反應的啟動則在7 d之前。我室黃復生等[12]的研究則提示,約氏瘧原蟲感染大劣按蚊后5 d在電鏡下其周圍即可見少數血細胞附著,即包被反應已經發生。在蚊對瘧原蟲的黑化包被反應中,需要絲氨酸蛋白酶級聯反應的啟動和PPO的活化。因此,絲氨酸蛋白酶轉錄水平的變化必然與PPO和PO有著密切關系。我室徐文岳等[13,14]對PPO的研究發現,在吸血后4~5 d,PPO的蛋白合成明顯增強;而對PO的研究則顯示,在血餐后5 d時PO活性最高,之后逐漸下降,這與本結果中吸血后5 d絲氨酸蛋白酶轉錄水平增高一致,提示SP與PPO及PO之間轉錄水平的變化存在相關性及它們在黑化反應中具有重要作用。由此推斷,大劣按蚊絲氨酸蛋白酶及PPO級聯反應在血餐后5 d時達到高峰。吸血后7 d和9 d較之5 d時的AdSP3轉錄水平發生下降,并趨于穩定水平,這可能是由于免疫刺激因子減少,導致轉錄水平下降并趨于穩定。
總之,本試驗在mRNA水平研究了AdSP3在大劣按蚊不同組織的表達分布及吸血和感染瘧原蟲后不同時相點的表達情況。結果提示,AdSP3在不同組織的分布和其轉錄水平的變化與PPO和瘧原蟲的感染相關,說明絲氨酸蛋白酶AdSP3在大劣按蚊抗約氏瘧原蟲感染中可能作為前酚氧化酶活化酶發揮了重要作用。絲氨酸蛋白酶在蚊抗瘧原蟲感染天然免疫及黑化包被反應中的作用是通過復雜的信號通路完成的[15],AdSP3在大劣按蚊對約氏瘧原蟲的黑化包被反應中的具體作用機制還有待進一步研究。
[1]Sakamoto M,Ohta M,Suzuki A,et al.Localization of the serine protease homolog BmSPH-1 in nodules of E.coli-injected Bomby x mori larvae and functional analysis of its role in nodule melanization[J].Dev Comp Immunol,2011,35(5):611-619.
[2]Cerenius L,Soderhall K.The prophenoloxidase-activating system in invertebrates[J].Immunol Rev,2004,198(1):116-126.
[3]Nappi AJ,Vass E.Cy totoxic reactions associated with insect immunity[J].Adv Exp Med Biol,2001,484:329-348.
[4]Mav rouli M D,Tsakas S,T heodorou GL,et al.M AP kinase mediate phagocy tosis and melanization via prophenoloxidase activation in medfly hemocytes[J].Biochimica et Biophysica Acta,2005,1744(2):145-156.
[5]Wang Y,Zou Zhen,Jiang HB.An ex pansion of the dual clipdomain serine proteinase family in Manduca sexta:Gene organization,expression,and evolution of prophenoloxidase-activating proteinase-2,hemolymph proteinase 12,and other related proteinases[J].Genomics,2006,87(3):399-409.
[6]Gorman MJ,Andreeva OV,Paskewitz SM.Sp22D:a multidomain serine protease with a putative role in insect immunity[J].Gene,2000,251(1):9-17.
[7]郝宏興,邱宗文,段建華,等.大劣按蚊AdPPO2、AdPPO3基因cDNA克隆與組織定位[J].第三軍醫大學學報,2004,26(6):470-472.
[8]Volz J,Osta M A,Kafatos FC,et al.The roles of two clip domain serine proteases in innate immune responses of the malaria vector Anopheles gambiae[J].J Biol Chem,2005,280(48):40161-40168.
[9]Vlachou D,Schlegelmilch T,Christophides GK,et al.Functional genomic analysis of midgut epithelial responses in A-nopheles during Plasmodium invasion[J].Curr Biol,2005,15(13):1185-1195.
[10]Jiang HB,Wang Y,Gu Yongli,et al.Molecular identification of a bevy of serine proteinase in Manduca Sexta hemolymph[J].Insect Biochemistry and Molecular Biology,2005,35(8):931-943.
[11]Xu WY,Huang FS,Hao HX,et al.Two serine protease from Anopheles dirus haemocytes exhibit changes in transcript abundance after infection of an incompatible rodent malaria parasite,Plasmodium yoelii[J].Veterinary Parasitology,2006,139(1):93-101.
[12]黃復生,王興相.大劣按蚊血細胞對約氏瘧原蟲卵囊黑化的作用[J].中國寄生蟲病防治雜志,1995,8(4):284-285.
[13]徐文岳,黃復生,況明書,等.約氏瘧原蟲卵囊黑化期間大劣按蚊血淋巴蛋白的分析[J].第三軍醫大學學報,2001,23(9):1098-1100.
[14]徐文岳,黃復生,張錫林,等.大劣按蚊血淋巴酚氧化酶與約氏瘧原蟲卵囊黑化關系的研究[J].第三軍醫大學學報,2001,23(4):440-442.
[15]Zou Z,Shin SW,Alvarez KS,et al.Distinct melanization pathways in the mosquito Aedes aegypti[J].Immunity,2010,32(1):41-53.
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30