抗高效氟吡甲禾靈日本看麥娘生長(zhǎng)速率與生長(zhǎng)密度研究
2011-08-16 05:14:48董立堯
雜草學(xué)報(bào) 2011年2期
張 彬,李 俊,2,董立堯,2
(1.南京農(nóng)業(yè)大學(xué)植物保護(hù)學(xué)院/農(nóng)業(yè)部作物病蟲(chóng)害監(jiān)測(cè)與防控重點(diǎn)開(kāi)放實(shí)驗(yàn)室,江蘇南京 210095; 2.南京農(nóng)業(yè)大學(xué)理學(xué)院/江蘇省農(nóng)藥學(xué)重點(diǎn)實(shí)驗(yàn)室,江蘇南京 210095)
?
抗高效氟吡甲禾靈日本看麥娘生長(zhǎng)速率與生長(zhǎng)密度研究
張 彬1,李 俊1,2,董立堯1,2
(1.南京農(nóng)業(yè)大學(xué)植物保護(hù)學(xué)院/農(nóng)業(yè)部作物病蟲(chóng)害監(jiān)測(cè)與防控重點(diǎn)開(kāi)放實(shí)驗(yàn)室,江蘇南京 210095; 2.南京農(nóng)業(yè)大學(xué)理學(xué)院/江蘇省農(nóng)藥學(xué)重點(diǎn)實(shí)驗(yàn)室,江蘇南京 210095)
抗性日本看麥娘種群與敏感種群種子在低溫(4 ℃)清水浸泡處理超過(guò)6 d后萌發(fā)率均能達(dá)90%以上,但抗性種群休眠解除的時(shí)間要長(zhǎng)于敏感種群;敏感種群在播種后的前15 d內(nèi)生長(zhǎng)速率快于抗性種群,但此后抗性種群生長(zhǎng)速率逐漸超過(guò)敏感種群;抗性種群在播種后20 d開(kāi)始長(zhǎng)出第2片葉,比敏感種群晚2 d,播種后25 d兩種群出現(xiàn)第2片葉的比例均超過(guò)95%,進(jìn)一步說(shuō)明在生長(zhǎng)后期抗性種群生長(zhǎng)速率超過(guò)敏感種群;在相同的播種密度下,抗性種群長(zhǎng)勢(shì)優(yōu)于敏感種群,150~200株/0.11m2的密度范圍內(nèi)有利于日本看麥娘的生長(zhǎng),抗性種群的最佳生長(zhǎng)密度為208株/0.11m2,而敏感種群為186株/0.11m2。
日本看麥娘; 解除休眠; 生長(zhǎng)速率; 生長(zhǎng)密度
種子休眠是大多數(shù)雜草種子都具有的特性,是雜草在長(zhǎng)期自然選擇過(guò)程中形成的對(duì)不良環(huán)境的適應(yīng)[1]。韓建國(guó)將種子休眠定義為“具有生活力而停留在不能發(fā)芽的狀態(tài)”[2]。而國(guó)外學(xué)者則將種子休眠定義為“成熟而完好無(wú)損的種子在最適宜的條件下不能發(fā)芽或延遲發(fā)芽的現(xiàn)象”[3-4]。種子休眠對(duì)處于惡劣環(huán)境中的植物保持其自身繁衍發(fā)展具有重要的生態(tài)學(xué)意義[5]。大多雜草種子都具有休眠的特性,不同種類的植物及不同品種之間種子休眠時(shí)間長(zhǎng)短差異很大。楊彩宏等分別采用地下沉積和低溫清水浸泡兩種休眠解除方法對(duì)日本看麥娘種子休眠解除進(jìn)行了相關(guān)的研究,結(jié)果表明低溫清水浸泡法能有效的解除所有日本看麥娘種群種子的休眠[6]。適合度(fitness)簡(jiǎn)單的說(shuō)就是植物的存活及繁殖成效,即在某種環(huán)境條件下,某已知基因型的個(gè)體將其基因傳遞到后代基因庫(kù)中的相對(duì)能力,是衡量個(gè)體存活和繁殖機(jī)會(huì)的尺度[7-8]。適合度越大,存活和繁殖機(jī)會(huì)越高。國(guó)內(nèi)外對(duì)抗性種群適合度的研究已經(jīng)有很多報(bào)道,大多學(xué)者已經(jīng)證明,新的抗性種群的產(chǎn)生會(huì)伴隨著適合度的降低[9-11],但也有研究表明抗藥性能導(dǎo)致其適合度升高[12-13],因此對(duì)兩種雜草種群之間相對(duì)適合度的比較研究就顯得非常重要。
1 材料與方法
1.1 供試材料
日本看麥娘(AlopecurusjaponicusSteud.)種子采集于2009年6月份。通過(guò)種子生物測(cè)定法檢測(cè)不同地點(diǎn)日本看麥娘種群的抗性水平。經(jīng)測(cè)定,將采集于江蘇省句容市油菜田的日本看麥娘作為抗性種群,將江蘇省南京市紫金山草坪地中的日本看麥娘作為敏感種群(種子萌發(fā)率均為90%以上)。
供試除草劑為10.8%高效氟吡甲禾靈EC,美國(guó)陶氏益農(nóng)公司生產(chǎn)。
1.2 試驗(yàn)方法
1.2.1 低溫清水浸泡時(shí)間對(duì)解除抗高效氟吡甲禾靈日本看麥娘種子休眠的方法研究 取足量大小均勻、籽粒飽滿的成熟日本看麥娘種子置于潔凈的培養(yǎng)皿中,加適量的蒸餾水(以浸泡種子為準(zhǔn)),置于4 ℃冰箱中。分別在第0、1、2、3、4、5、6、7、8、9天取出40粒放于直徑為9 cm鋪有1張濾紙的培養(yǎng)皿中,加5 mL超純水,加蓋以保持水分,置于光照培養(yǎng)箱中培養(yǎng)觀察(溫度:白天18 ℃,夜間13 ℃;光周期:12 D/12 L),4次重復(fù)。每天統(tǒng)計(jì)種子的萌發(fā)數(shù)并計(jì)算萌發(fā)率(胚芽長(zhǎng)至2 mm時(shí)即為萌發(fā)),并比較兩個(gè)種群種子萌發(fā)率間的差異。
1.2.2 抗高效氟吡甲禾靈日本看麥娘幼苗生長(zhǎng)速率的研究 試驗(yàn)采用盆缽法。將細(xì)土均勻的裝入直徑為9 cm的塑料盆缽內(nèi),每盆播30粒已解除休眠的日本看麥娘種子,放于室內(nèi)培養(yǎng)箱中培養(yǎng),培養(yǎng)條件如1.2.1。各盆缽?fù)寥朗冀K保持濕潤(rùn),4次重復(fù)。出苗后每盆定苗20株,每隔2 d記錄日本看麥娘的株高,觀察記錄雜草長(zhǎng)第2片葉的時(shí)間,并比較兩個(gè)種群幼苗的生長(zhǎng)速率。試驗(yàn)期間人工拔除其他雜草。
1.2.3 抗高效氟吡甲禾靈日本看麥娘最佳種群生長(zhǎng)密度的研究 試驗(yàn)采用盆缽法。將均勻的細(xì)土裝入直徑為11.5 cm的塑料盆缽內(nèi),每盆分別播50、40、30、20、10粒已解除休眠的日本看麥娘種子,放于室內(nèi)培養(yǎng)箱中培養(yǎng),培養(yǎng)條件如1.2.1。各盆缽?fù)寥朗冀K保持濕潤(rùn),4次重復(fù)。出苗后分別每盆定苗40、35、30、25、20、15、10、5株,出苗后第35天測(cè)量日本看麥娘的株高,并比較兩個(gè)種群間幼苗株高的差異。試驗(yàn)期間人工拔除其他雜草。
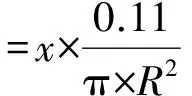
其中,x為每盆的雜草株數(shù),R為盆缽的半徑。
1.3 統(tǒng)計(jì)分析
試驗(yàn)采用Excel 2003計(jì)算平均值和標(biāo)準(zhǔn)差并作圖,用DPS v3.01統(tǒng)計(jì)軟件進(jìn)行統(tǒng)計(jì)分析。
雜草隨時(shí)間變化而變化規(guī)律應(yīng)用一元回歸模型進(jìn)行研究,即以雜草生長(zhǎng)時(shí)間為自變量(x),以雜草的株高為因變量(y),公式為y=f(x)。以相關(guān)系數(shù)最大、離回歸方差最小為原則進(jìn)行優(yōu)選[14-15]。
不同種植密度下日本看麥娘的長(zhǎng)勢(shì)應(yīng)用一元二次曲線方程模型進(jìn)行研究,即以雜草的種植密度為自變量(x),以雜草的株高為因變量(y),公式為y=ax2+bx+c。以相關(guān)系數(shù)最大、離回歸方差最小為原則進(jìn)行優(yōu)選[14-15]。
2 結(jié)果與分析
2.1 低溫清水浸泡時(shí)間對(duì)解除抗高效氟吡甲禾靈日本看麥娘種子休眠的影響
抗高效氟吡甲禾靈日本看麥娘種子休眠解除時(shí)間長(zhǎng)于敏感種群(圖1)。隨著低溫清水浸泡時(shí)間的延長(zhǎng),2個(gè)日本看麥娘種群種子萌發(fā)率均逐漸增加。未經(jīng)低溫清水浸泡處理的兩個(gè)日本看麥娘種群種子萌發(fā)率很低,分別只有5.56%、16.67%。在第6天時(shí)2個(gè)日本看麥娘種群種子萌發(fā)率分別達(dá)到84.44%、92.22%,但抗性種群萌發(fā)率仍低于敏感種群。因此,抗性日本看麥娘種群種子比敏感種群難于解除休眠。
2.2 抗高效氟吡甲禾靈日本看麥娘生長(zhǎng)速率
抗高效氟吡甲禾靈日本看麥娘生長(zhǎng)速率不同于敏感種群(圖2)。在生長(zhǎng)早期,抗性種群生長(zhǎng)速度稍微慢于敏感種群,但生長(zhǎng)18 d以后,抗性種群生長(zhǎng)速率逐步加快,并超過(guò)敏感種群。通過(guò)兩個(gè)日本看麥娘種群生長(zhǎng)速率的線性比較也驗(yàn)證了這點(diǎn)(R為0.617 2;S為0.511 4)。


抗高效氟吡甲禾靈日本看麥娘第二片葉片的出現(xiàn)時(shí)間晚于敏感種群(圖3),在種植第18天時(shí),敏感種群生出第二片葉,而第19天時(shí)抗性種群開(kāi)始生第二片葉,第25天時(shí),兩種日本看麥娘種群幾乎都長(zhǎng)出第二片葉,出現(xiàn)比率達(dá)到95%以上。通過(guò)曲線回歸方程也可以看出抗性種群的二次項(xiàng)系數(shù)為0.952 4,敏感種群的二次項(xiàng)系數(shù)為-0.937 5,說(shuō)明抗性日本看麥娘種群在生長(zhǎng)后期快于敏感種群。

2.3 抗高效氟吡甲禾靈日本看麥娘最佳種群生長(zhǎng)密度
在同等密度下抗性日本看麥娘種群長(zhǎng)勢(shì)優(yōu)于敏感種群(圖4)。150 ~200株/0.11m2種植密度有利于日本看麥娘的生長(zhǎng),低密度和高密度都不利于日本看麥娘的生長(zhǎng)。初步建立了日本看麥娘種群密度與株高的二次曲線方程,通過(guò)曲線方程可以推算出抗性日本看麥娘種群的最佳生長(zhǎng)密度為208株/0.11m2,大于敏感種群的186株/0.11m2。

3 討論
種子休眠是植物在長(zhǎng)期系統(tǒng)發(fā)育過(guò)程中形成的抵抗不良環(huán)境條件的適應(yīng)性,是調(diào)節(jié)種子萌發(fā)的最佳時(shí)間和空間分布的有效方法,具有普遍的生態(tài)意義。種子休眠時(shí)間的長(zhǎng)短因不同種類和不同品種間差異很大,而且不同種子破除休眠的方法不盡相同。對(duì)于室內(nèi)雜草的研究,低溫清水浸泡破除日本看麥娘種子休眠是最有效、最快捷的方法[6]。試驗(yàn)中通過(guò)對(duì)抗高效氟吡甲禾靈日本看麥娘種群種子低溫清水浸泡解除休眠研究發(fā)現(xiàn),與敏感種群相比,抗性種群的休眠解除明顯困難。但到目前為止,出現(xiàn)這種結(jié)果的原因尚未有相關(guān)的報(bào)道和說(shuō)明,因此需要進(jìn)一步研究。
大多抗性種群的產(chǎn)生都會(huì)伴隨其適合度的降低,但本試驗(yàn)研究發(fā)現(xiàn)抗性日本看麥娘種群在某些特性上優(yōu)于敏感種群。在生長(zhǎng)早期,抗高效氟吡甲禾靈日本看麥娘種群生長(zhǎng)速率慢于敏感種群,但生長(zhǎng)后期抗性種群生長(zhǎng)速率及長(zhǎng)勢(shì)要優(yōu)于敏感種群,與Sibony and Rubin(2002)和Holt and Thill(1994)報(bào)道的致使產(chǎn)生抗性的變異并沒(méi)有降低適合度相吻合。Shrestha等在2010報(bào)道了對(duì)草甘膦產(chǎn)生抗性的加拿大乍蓬(Conyzacanadensis)在田間試驗(yàn)研究中表現(xiàn)出強(qiáng)于敏感種群的競(jìng)爭(zhēng)優(yōu)勢(shì)[16],但對(duì)于抗性種群適合度增高的原因尚未見(jiàn)報(bào)道,尤其是本試驗(yàn)中發(fā)現(xiàn)的抗性日本看麥娘種群其表現(xiàn)出的適合度增高的原因,需要進(jìn)一步的試驗(yàn)證明。
日本看麥娘屬群集性生長(zhǎng)的一種雜草,田間出苗率很高[17]。過(guò)高或過(guò)低的種群密度都不利于其正常的生長(zhǎng)。試驗(yàn)證明,150~200株/0.11m2范圍內(nèi)有利于日本看麥娘的生長(zhǎng),抗性日本看麥娘種群的最佳生長(zhǎng)密度208株/0.11m2,而敏感型為186株/0.11m2,這是否與抗性和敏感兩種種群的生長(zhǎng)環(huán)境有關(guān),需要進(jìn)一步的試驗(yàn)研究。但其最佳種群密度結(jié)果的發(fā)現(xiàn),對(duì)以后田間日本看麥娘的控制和防除有一定的幫助和支持。
[1]蘇少泉. 雜草科學(xué)[M]. 北京:農(nóng)業(yè)出版社,1993:24-32.
[2 ]韓建國(guó),李青豐,陳寶書,等. 牧草種子學(xué)[M]. 北京:中國(guó)農(nóng)業(yè)大學(xué)出版社,2000:68-82.
[3]Prada D,Romagosa I,Ullrich S E,et al. A centromeric region on chromosome 6(6H) affects dormancy in an induced mutant in barley[J]. Journal of Experimental Botany,2005,56(409):47-54.
[4]Romagosa I,Prada D,Moralejo M A,et al. Dormancy,ABA content and sensitivity of a barley mutant to ABA application during seed development and after ripening[J]. Jourmal of Experimental Botany,2001,52(360):1499-1506.
[5]張 鵬,沈海龍. 白蠟樹(shù)屬樹(shù)種種子休眠及其萌發(fā)的調(diào)控[J]. 植物生理學(xué)通訊,2006,42(2):354-358.
[6]楊彩宏,董立堯,李 俊,等. 油菜田日本看麥娘對(duì)高效氟吡甲禾靈抗藥性的研究[J]. 中國(guó)農(nóng)業(yè)科學(xué),2007,40(12):2759-2765.
[7]劉鳳沂,須志平,薄仙萍,等. 昆蟲(chóng)抗藥性與適合度[J]. 昆蟲(chóng)知識(shí),2008,45(3):374-378.
[8]孟香清,芮昌輝,趙建周,等. 抗三氟氯氰菊酯棉鈴蟲(chóng)種群相對(duì)適合度研究[J]. 植物保護(hù),1998,24(6):12-14.
[9]Park K W,Smith C A,Ball D A,et al. Ecological fitness of acetolactate synthaseinhibitor-resistant and -susceptible downy brome (Bromustectorum) biotypes[J]. Weed Science,2004,52:768-773.
[10]Hartzler R G,Battles B A. Reduced fitness of velvetleaf (Abutilontheophrasti) surviving glyphosate[J]. Weed Technology,2001,15:492-496.
[11]Massinga R A,Al-Khatib K,Amand P St,et al. Relative fitness of imazamox-resistant common sunflower and prairie sunflower[J]. Weed Science,2005,53:166-174.
[12]Steckel L E,Gwathmey C O. Glyphosate-resistant horseweed (Conyzacanadensis) growth,seed production,and interference in cotton[J]. Weed Science,2009,57:346-350.
[13]Poston D H,Wilson H P. Growth and development of imidazolinone-resistant and -susceptible smooth pigweed biotypes[J]. Weed Science,2002,50:485-493.
[14]王順建,張 玉,株良備. 安徽淮北地區(qū)麥田蚤綴生物學(xué)特性及生態(tài)經(jīng)濟(jì)閾值研究[J]. 雜草科學(xué),2002(3):23-26.
[15]于金鳳,王金信,陳茂學(xué),等. 麥田混生雜草生態(tài)經(jīng)濟(jì)閾值的研究[J]. 植物保護(hù),2002,28(5):13-15.
[16]Shrestha A,Hanson B D,Fidelibus M W,et al. Growth,phenology,and intraspecific competition between glyphosate-resistant and glyphosate-susceptible horseweeds (Conyzacanadensis) in the San Joaquin Valley of California[J]. Weed Science,2010:58:147-153.
[17]曹春田,李國(guó)宇,劉新峰,等. 麥田惡性雜草日本看麥娘的發(fā)生及防治[J]. 河南農(nóng)業(yè)科學(xué),2003(10):73.
Research on Growth Rate and Planting Density of Japanese Foxtail (Alopecurusjaponicus) Resistant to Haloxyfop-R-methyl
ZHANG Bin1, LI Jun1,2, DONG Li-yao1,2
(1.College of Plant Protection,Nanjing Agricultural University/Key Laboratory of Monitoring and Management of Crop Diseases and Pest Insects,Ministry of Agriculture,Nanjing 210095,China;2.College of Science,Nanjing Agricultural University/Jiangsu Key Laboratory of Pesticide Science,Nanjing 210095,China)
The germination rate of Japanese foxtail(Alopecurusjaponicus)in the resistant and sensceptible population to haloxyfp-R-methyl could reach to more than 90% after 6 days for water soaking at 4 ℃,but dormancy breaking in the resistant population needed more time than that of sensceptible population.The resistant population presented a lower growths rate than that of sensceptible population after 15 days of sowing,but after that date the growth rate in resistant poputation was getting higher than that of sensceptible poputation. The resistant population began to appear the second leaf after 20 days for sowing that was later than sensceptible population by 2days,and the rate for the second leaf appearance in both populations could reach to 95% affer 25 days of sowing which proved that the resistant population grew more rapidly than that of sensceptible population during the later period of growth time after sowing. The resistant population performed a better growth than that of sensceptible population under the same planting density. Japanese foxtail could grow well under the planting density of 150~200 plants/0.11m2. The suitable growth density for resistant population cowld he designed as 208 plants/0.11m2,but 186 plants/0.11m2for sensceptible population.
Japanese foxtail(Alopecurusjaponicus);dormancy breaking; growth rate; growth densitity
2011-04-11
國(guó)家自然科學(xué)基金(編號(hào):30971928);國(guó)家博士點(diǎn)基金新教師類項(xiàng)目(編號(hào):20090097120046)。
張 彬(1985—),男,碩士研究生,研究方向?yàn)槌輨┒纠砑翱顾幮浴-mail:2008102117@njau.edu.cn。
董立堯,博士,教授,從事除草劑毒理及抗藥性研究。Tel:(025)84395672; E-mail:dly@njau.edu.cn。
S451
A
1003-935X(2011)02-0043-04
猜你喜歡
科教新報(bào)(2022年22期)2022-07-02 12:34:28
今日農(nóng)業(yè)(2021年5期)2021-11-27 17:22:19
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
高中生學(xué)習(xí)·閱讀與寫作(2019年2期)2019-09-10 07:22:44
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
現(xiàn)代農(nóng)業(yè)(2015年5期)2015-02-28 18:40:49
