植物青枯菌Ⅵ型分泌系統核心基因vasK突變株的構建及其致病性的測定
2011-08-27 00:40:22張麗勍許景升
植物保護 2011年4期
張麗勍, 許景升, 徐 進, 馮 潔
(中國農業科學院植物保護研究所,植物病蟲害生物學國家重點實驗室,北京 100193)
植物青枯菌Ⅵ型分泌系統核心基因vasK突變株的構建及其致病性的測定
張麗勍, 許景升, 徐 進, 馮 潔*
(中國農業科學院植物保護研究所,植物病蟲害生物學國家重點實驗室,北京 100193)
[目的]通過測試vasK基因突變株對番茄的致病力變化,評價該基因在青枯菌致病過程中的作用。[方法]根據青枯菌(Ralstonia solanacearum)中存在的Ⅵ型分泌系統基因簇中的核心基因vasK序列設計PCR引物,擴增并克隆vasK基因,將慶大霉素抗性基因(Gm)插入vasK基因內部,克隆至自殺質粒pK18mobsacB中,獲得重組自殺質粒pK18-vasK-Gm。將自殺質粒電轉化至青枯菌GMI1000感受態細胞中,采用同源重組雙交換法,將野生型vasK基因置換。對vasK基因突變菌株進行三步篩選和PCR擴增鑒定。[結果]篩選獲得了具有慶大霉素抗性的目標基因被抗性基因替換的青枯菌突變株(GMI1000-m)。土壤接種番茄青枯菌結果顯示,突變株GMI1000-m的致病性較野生型GMI1000明顯下降。[結論]vasK基因在青枯菌致病過程中具有重要作用。
青枯菌;Ⅵ型分泌系統;vasK基因
病原細菌通常利用蛋白的分泌來介導與寄主植物的相互作用[1]。革蘭氏陰性病原細菌能夠通過微小的裝置將蛋白運送到胞外,或直接運輸到寄主細胞中。這些微小的裝置稱為分泌系統。目前為止,發現在革蘭氏陰性病原菌中至少存在著6種不同類型的分泌系統(Ⅰ~Ⅵ型分泌系統),這些分泌系統通過分泌或注射的方式釋放胞外蛋白或效應子,以刺激或干擾寄主細胞的進程,從而能夠調控一系列的病原細菌—寄主細胞的相互作用。Ⅵ型分泌系統是新近發現的一種細菌蛋白分泌系統。具有Ⅵ型分泌系統的一些革蘭氏陰性病原菌對人類的健康構成了威脅:Vibrio cholerae,Yersinia pestis,Francisella tularensis,Burkholderia mallei,Salmonella ty phimurium,致病的Escherichia coli及條件性病原微生物如Pseudomonas aeruginosa,Burkholderia cenocepacia,Aeromonas hydrophila[2-5]。Ⅵ型分泌系統在細菌基因組中以基因簇形式存在,該基因簇通常編碼12~25個蛋白。核心組分包括IcmF類蛋白、IcmH 類蛋白(DotU)、AAA+ATP酶ClpV、未知脂蛋白以及泌出蛋白VgrG和Hcp。根據T6SS蛋白位于細菌細胞的不同部位可以將T6SS蛋白分為穿刺裝置、外膜蛋白、跨膜蛋白和內膜蛋白。
其中,VasF和VasK含有跨膜結構域和螺旋-螺旋(coiled-coil)結構,屬于IcmH和IcmF類蛋白。IcmH和IcmF可以有助于細菌在寄主細胞膜形成蛋白轉運孔,幫助細菌泌出毒性因子。因此作為細菌細胞周質蛋白的VasF和VasK在蛋白泌出中具有重要作用。VasK除了參與蛋白泌出外,還可能具有識別效應子蛋白和信號轉導的功能,因為VasK還擁有G-蛋白家族的P-loop結構域,該結構域具有核酸磷酸化水解酶的功能[6]。
經分析發現在青枯菌GMI 1000菌株大質粒中也存在Ⅵ型分泌系統基因簇,同源性達到80%。本試驗采用自殺性質粒載體與染色體進行基因同源交換技術,構建了青枯菌Ⅵ型分泌系統核心基因vasK的定位突變,通過測試vasK基因突變株對番茄的致病力變化,評價了該基因在青枯菌致病過程中的作用。
1 材料和方法
1.1 菌株和質粒
試驗中使用的菌株和質粒見表1。青枯菌GMI1000生長在NA平板或液體培養基中,28℃條件下培養。大腸桿菌生長在LB平板或液體培養基中,37℃條件下培養。10%蔗糖NA培養基為NA培養基加入相應體積的蔗糖儲存液(50%蔗糖,115℃滅菌20 min)至終濃度為10%。抗生素使用濃度分別為:氨芐青霉素(Ap)100 μ g/mL,卡那霉素(Kan)20 μ g/mL,慶大霉素(Gm)10 μ g/mL 。
1.2 主要試劑
普通Taq酶、dNTP 、SalI、PstI、T4 DNA Ligase購自大連寶生物(TaKaRa)公司,2×TaqPlatinum PCR MasterMix 、DL2000 、500 bp DNA ladder購自天根公司,瓊脂糖凝膠回收試劑盒購自北京百泰克公司,氨芐青霉素(Amp)、卡那霉素(Kan)、慶大霉素(Gm)、IPTG 、X-gal購自Sigma公司,蔗糖為上海生工公司產品,dATP、pMD19-T載體購自大連寶生物(TaKaRa)公司,引物為上海生工公司合成。
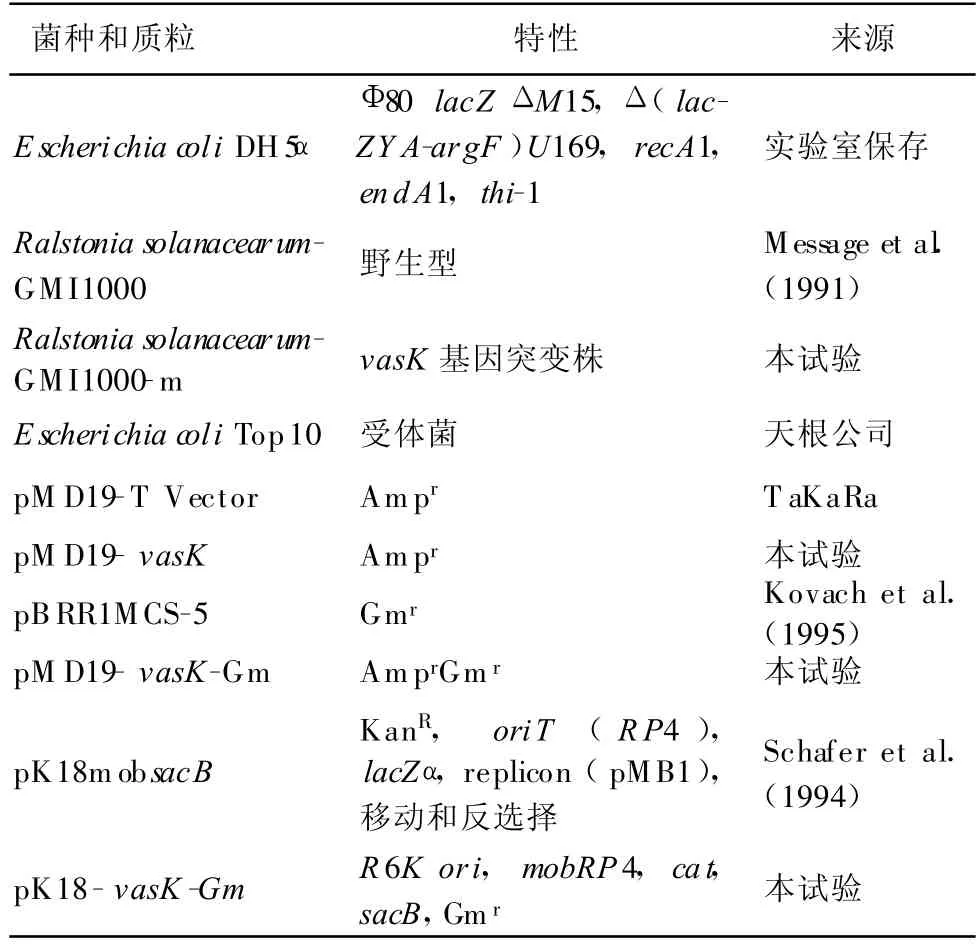
表1 菌種和質粒
1.3 vasK1-1455基因片段PCR擴增
根據vasK核苷酸序列設計引物,擴增vasK部分DNA片段,用于構建重組自殺質粒。上游引物從vasK起始密碼子的第1 bp開始,下游引物至第1 455 bp,共1 455 bp,其中在第429 bp和1 112 bp處存在PstI酶切位點,在1 444 bp處存在SalI酶切位點。部分vasK基因上下游引物序列分別為,vasK1:5′-T TCATCACCT TCTCTACACTT-3′;vasK2:5′-TACATATCACATCAAATTCCTCAACTTCC-3′。采用PCR擴增,擴增條件:94℃,3 min;35個循環包括:94℃,30 s;60℃,30 s;72℃,1.5 min;1個循環:72℃,10 min。
1.4 慶大霉素基因的PCR擴增
根據質粒載體pBBR1MCS-5(GenBank登錄號:U25061)中慶大霉素抗性基因的序列,設計引物擴增慶大霉素基因,作為重組突變體篩選標記基因。上下游引物序列分別為:5′-ATAATGCAT GGACGCACAC-CGTGGAAA-3′、5′-ATAATGCAT GGCGGCGTTGTGACAATTT-3′,下劃線部分為NsiI的酶切位點。采用降落PCR擴增條件:95℃5 min;30個循環:95℃1 min;70℃1 min(每個循環降低0.7℃);72℃1 min;10個循環:95℃1 min;55℃1 min;72℃1 min;72℃10 min。
1.5 vasK基因重組自殺質粒pK18-vasK-Gm的構建及鑒定
PCR產物的回收、連接、轉化、質粒提取及酶切參照文獻指南[9]或按照試劑盒說明書推薦的條件進行,構建過程見圖1。
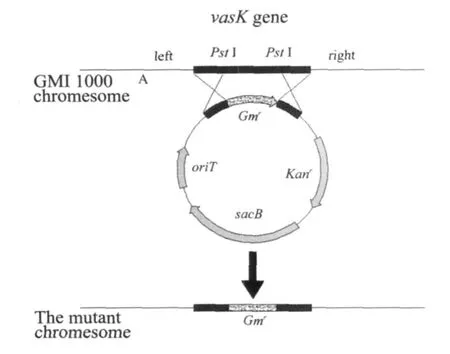
圖1 重組質粒pMD19-vasK-Gm的構建
1.6 青枯菌GMI 1000感受態細胞的制備及電轉化
青枯菌感受態細胞的制備及電轉化參照Ausubel等[11]和Lavie等[12]的方法并略有改動。于28℃,用NA培養液搖瓶培養青枯菌40 h,將菌液冰浴冷卻10~15 min,然后轉移到預冷的1.5 mL的Eppendorf管中。于4℃,5 000 r/min離心5 min,棄上清,沉淀用1 mL的ddH2O懸浮,離心,收集菌體,再重復一次。用1 mL的10%的甘油懸浮菌體,于4℃,5 000 r/min離心5 min,棄上清,沉淀用100 μ L 10%的甘油懸浮,加入1 μ g pK18-vasK-Gm質粒DNA混合物全部轉移至電極杯(0.2 cm gap electroporation cuvette,BioRad)中,將電極杯放在Gene Pulser(BioRad)電轉,參數為:2.5 kV,25 μ F,200 Ω[13]。
1.7 vasK基因突變菌株的篩選與鑒定
電轉結束后立即加入1 mL NA液體培養基,轉移至新的離心管中,28℃振蕩復蘇24 h,然后依次進行如下3次篩選。(1)初篩:將轉化后的菌液涂布于含卡那霉素和慶大霉素的NA固體培養基平板上,28℃培養24~48 h,至單個菌落形成,提取基因組DNA,用vasK1/vasK2引物擴增鑒定。(2)復篩:挑取上步的單菌落接種于NA液體培養基中,培養過夜后梯度稀釋并涂布于含有10%蔗糖和慶大霉素的NA固體平板上,28℃培養24~48 h,至單個菌落形成,篩選雙重組子。(3)第3次篩選:挑取上步的單菌落分別轉移至含有慶大霉素、卡那霉素和10%蔗糖平板上進行篩選。最終,篩選出對卡那霉素(Kans)和蔗糖(Sucs)敏感對慶大霉素具有抗性(Gmr)的菌落提取基因組,用vasK1/vasK2引物擴增鑒定[13]。
1.8 突變菌株和野生型菌株致病性的測定
參照He等[14]傷根澆注菌液接種法(土壤接種法)進行青枯菌接種試驗。選用番茄感病品種種子播種于滅菌蛭石中育苗,待幼苗生長至3~4片真葉時選取植株大小和根系發育均勻一致的幼苗,移栽于直徑15 cm裝有混合土(1∶1的普通田園壤土和草炭土)的營養缽中,每缽3株。待番茄植株幼苗生到7~8葉期,采用傷根澆注菌液接種法進行青枯菌接種試驗。在植株距莖1 cm處用手術刀向下劃深3~4 mm,長4 cm的口子,每株番茄苗澆30 mL濃度為3×106cfu/mL的菌懸液,每處理10株,3個重復。青枯病的病情調查參照Denny[15]等的方法。
2 結果與分析
2.1 vasK基因片段的克隆
vasK基因全長為3 942 bp,本試驗選擇克隆了結構基因的760 bp作為靶序列用于同源重組。將純化后的部分vasK基因的PCR擴增產物克隆入
pMD19-T載體,命名為pMD19-vasK。經SalI酶切和測序鑒定,與預期完全一致(圖2)。
2.2 慶大霉素基因的克隆及重組質粒pMD19-vasK-Gm的構建及鑒定
慶大霉素基因Gm,長度為855 bp。將純化后的慶大霉素基因PCR擴增產物經過NsiI酶切克隆至pMD19-vasK載體的vasK基因片段的
PstI酶切位點。將Gm基因插入到vasK基因的內部,一則將vasK基因突變使其失活,再則為以后篩選突變株提供抗生素標記。經含氨芐青霉素和慶大霉素的 LB平板進行篩選,獲得重組質粒pMD19-vasK-Gm,經酶切鑒定與預期片段大小一致。
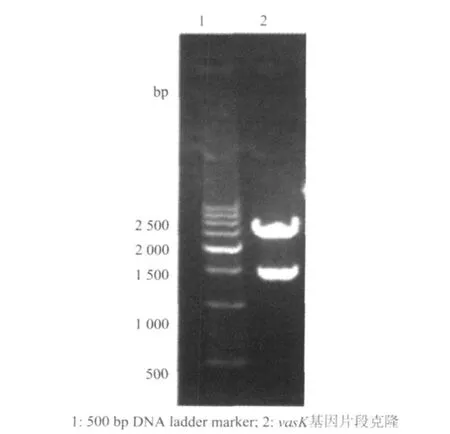
圖2 重組質粒pMD19-vasK的酶切鑒定圖
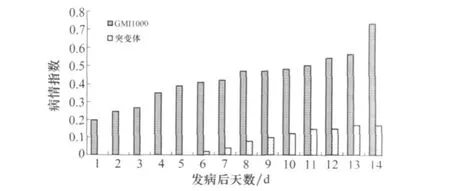
圖3 vasK基因突變株致病力的測定
2.3 自殺質粒pK18-vasK-Gm的構建及鑒定
重組質粒pMD19-vasK-Gm經SalI酶切后,瓊脂糖凝膠電泳分離1 396 bp的vasK-Gm片段,并亞克隆至經同樣酶切的pK18mobsacB質粒載體中,構建pK18-vasK-Gm自殺性質粒載體,轉化至E.coliDH5α菌株。在含卡那霉素和慶大霉素的LB平板上篩選克隆菌,獲得了重組質粒pK18-vasK-Gm,經酶切鑒定與預期片段大小一致,序列測定結果表明,該重組子為內部含有慶大霉素基因的vasK基因的重組質粒。
2.4 vasK基因敲除菌株的篩選與鑒定
采用電擊轉化方法將質粒pK18-vasK-Gm轉化至野生菌株中,通過三步篩選法獲得了突變株。最后對Gmr、Sucs、Kans的菌株進行PCR擴增,篩選獲得了突變株GMI1000-m,野生型菌株GMI1000的vasK基因已被突變了的vasK基因所置換。本試驗最初選擇了vasK基因的760 bp作為靶序列用于同源重組,基因本身含有PstI酶切位點,在與慶大霉素基因Gm連接的過程中,會損失219 bp的片段,慶大霉素基因Gm長855 bp,將純化后的慶大霉素基因 PCR擴增產物經過NsiI酶切克隆至pMD19-vasK載體的vasK基因片段的PstI酶切位點,vasK-Gm片段長1 396 bp。
2.5 敲除菌株和野生型菌株致病性的測定
野生型菌株接種7 d開始發病,突變株在野生型發病后第6天才出現癥狀。野生型發病后12 d,當野生型菌株病情指數達到0.56時,突變株的僅為0.17,表明vasK基因與青枯菌的致病力密切相關(圖3)。
3 結論與討論
2006年,科學家報道了在Vibrio cholerae和Pseudomonasaeruginosa中存在一種新型的分泌系統,并將其命名為Ⅵ型分泌系統[1,17]。在Ⅵ型分泌系統中通常存在一個基因簇。該基因簇通常編碼12~25個蛋白,只有DotU和IcmF樣的蛋白在其他分泌系統中也存在(T4SS)[18]。植物細菌性青枯病是由茄勞爾氏菌(Ralstonia solanacearum)引起的一種世界性分布的重大病害。2002年法國科學家Salanoubat等[19]在 Nature上報道了青枯細菌(GMI 1 000菌株,1號小種)的全基因組序列。
Ⅵ型分泌系統的作用機制表明,輔助蛋白VasK(IcmF蛋白家族)引起細菌細胞表面的重組,導致了對寄主細胞黏附度的增加。VasK同時幫助運輸和信號識別作用,因此可以增加運輸的效率。
本試驗圍繞青枯菌的Ⅵ型分泌系統與致病性的關系開展了研究,通過基因敲除的方法破壞了青枯菌T6SS基因簇中的vasK基因,發現突變菌株的致病力明顯降低,初步證實了vasK基因在青枯菌的致病過程中可能起到了重要作用。研究初步證實了vasK基因與青枯菌致病力可能相關。在致病力測定的基礎上,需要進一步分析vasK基因突變對青枯菌泌出蛋白的影響,以期揭示Ⅵ型分泌系統在青枯菌致病過程中的作用。
由于Ⅵ型分泌系統的研究主要集中在人類和動物病原細菌,而對植物病原細菌T6SS的研究僅有兩例報道。因此對植物青枯菌Ⅵ型分泌系統的研究,對探究T6SS致病機理具有非常重要的意義。
[1]Mougous J D,Cuff M E,Raunser S,et al.A virulence locus ofPseudomonasaeruginosaencodes a protein secretion apparatus[J].Science,2006,312:1526-1530.
[2]Das S,Chaudhuri K.Identification of a unique IA HP(IcmF associated homologous proteins)cluster inVibrio choleraand other proteobacteria throughin silicoanalysis[J].In Silico Biology,2003,3:287-300.
[3]Schell M A,Ulrich R L,Ribot W J,et al.Type VI secretion is a major virulence determinant inBurkholderia mallei[J].Molecular Microbiology,2007,64:1466-1485.
[4]Shalom G,Shaw J,Thomas M.Invivoexpression technology identifies a type VI secretion system locus inBurkholderia pseudomalleithat is induced upon invasion of macrophages[J].Microbiology,2007,153:2689-2699.
[5]Yen Y T,Bhattacharya M,Stathopoulos C.Genome-widein silicomapping of the secretome in pathogenicYersinia pestisKIM[J].FEMS Microbiol Lett,2008,279:56-63.
[6]Hamilton H L,Dominguez N M,Schwartz K J,et al.Neisseria gonorrhoeaesecretes chromosomal DNA via a novel type IV secretion sy stem[J].MolecularMicrobiology,2005,55:1704-1721.
[7]Message B,Boistard P,Pitrat M,et al.A new class of fluidal avirulent mutants ofPseudomonas solanacearumunable to induce a hypersensitive reaction[C].Proceeding s of the 4th International Conference on Plant Pathogenic Bacteria,1N RA,Angers,France,1978:823-833.
[8]Kovach M E,Elzer P H,Hill D S,et al.Four new derivatives of the broad host range cloning vector pBBR1M CS,carrying different antibiotic—resistance cassettes[J].Gene,1995,166(1):175-176.
[9]Schafer A A,Tauch W J,Kalinowsski G,et al.Small mobilizable multi-purpose cloning vectors derived fromEscherichia coliplasmids pK18 and pK19:selection of defined deletions in the chromosome ofCorynrbacterium glutamicum[J].Gene,1994,145:69-73.
[10]Sambrook J,Russell D W.Molecular cloning:A laboratory manual[M].3rd ed.Cold Spring Harbor Laborato ry Press,2002.
[11]Ausubel F M,Brent R,Kingston R E,et al.Sho rt protocols in molecular biology[M].3rd ed.John Wiley and Sons,Chichester,1998.
[12]Lavie M,Shillington E,Eguiluz C,et al.PopP 1,a new member of the YopJ/Av rRxv family of typeⅢeffector proteins,acts as a host specificity factor and modulates aggressiveness ofRalstonia solanacearum[J].M olecular Plant Microbe Interactions,2002,15(10):1058-1068.
[13]張爭,何禮遠,馮潔,等.植物青枯菌aac基因突變株的構建及其致病性的測定[J].農業生物技術學報,2009,17(3):522-528.
[14]He L Y,Sequeira L,Kelman A.Characteristics of strains ofPseudomonas solanacearumfrom China[J].Plant Disease,1983,67(12):1357-1361.
[15]Denny T P,Carney B F,Schell M A.Inactivation of multiple virulence genes reduces the ability ofPseudomonas solanacearumto cause wilt symptoms[J].Molecular Plant Microbiology Interaction,1990,3(5):293-300.
[16]吳乃虎.基因工程原理[M].北京:科學出版社,2001.
[17]Pukatzki S,Ma A T,Sturtevant D,et al.Identification of a conserved bacterial protein secretion system inVibrio choleraeusing theDictyosteliumhost model sy stem[J].Proceeding s of the National Academy of Sciences of the United States of America,2006,103:1528-33.
[18]Alain Filloux,Abderrahman Hachani,Sophie Bleves.The bacterial type VI secretion machine:yet another player fo r protein transport across membranes[J].Microbiology,2008,154:1570-1583.
[19]Salanoubat M,Genin S,Artiguenave F,et al.Genome sequence of the plant pathogenRalstonia solanacearum[J].Nature,2002,415(6871):497-502.
Construction and pathogenicity tests of the mutated strain of the core gene vasK of typeⅥsecretion system in Ralstonia solanacearum
Zhang Liqing, Xu Jingsheng, Xu Jin, Feng Jie
(State Key Laboratory for Biology of Plant Disease and Insect Pests,Institute of Plant Protection,Chinese Academy of Agricultural Sciences,Beijing100193,China)
[Objective]The change in pathogenicity of thevasKmutant to the tomato was investigated to estimate the function of thevasKgene in the pathogenicity process ofRalstonia solanacearum.[Method]According to the sequence ofvasKgene,a key component of typeⅥ secretion system inRalstonia solanacearum,PCR primers were designed to amplifyvasKgene.vasKgene was mutated by inserting a gentamicin?3-acetyltransferase gene(Gmgene),and the mutatedvasKwas inserted into suicide vector pK18mobsacB,resulting in pK18-vasK-Gm.The vector pK18-vasK-Gm was then introduced intoR.solanacearumGMI1000.ThevasKmutant,named GMI1000-m,was generated by homologous recombination and selected by a three-step method.GMI1000-m was identified by PCR amplification.[Result]The pathogenicity test of the mutant GMI1000-m was decreased compared with the wild type GMI1000.[Conclusion]ThevasKgene was an important factor in the pathogenesis ofR.solanacearum.
Ralstonia solanacearum; typeⅥsecretion system;vasKgene
Q 753
A
10.3969/j.issn.0529-1542.2011.04.007
2010-04-26
2010-10-15
國家重點基礎研究發展計劃(“973”)項目(2009CB119200);國家科技支撐計劃項目(2006BAD08A14);國家高技術研究與發展計劃(“863”)項目(2006AA10Z432)
*通信作者E-mail:jfeng@ippcaas.cn
