紅谷霉素對金黃色葡萄球菌抑菌機制的研究
2011-10-09 02:28:02薛秀園涂國全
天然產物研究與開發 2011年1期
涂 璇,薛秀園,涂國全
江西農業大學生物科學與工程學院,南昌 330045
紅谷霉素對金黃色葡萄球菌抑菌機制的研究
涂 璇,薛秀園,涂國全*
江西農業大學生物科學與工程學院,南昌 330045
紅谷霉素是一種抗細菌的新型抗生素。為探討紅谷霉素對金黃色葡萄球菌的抑菌機制,本文采用濃度為 EC50和 EC90的紅谷霉素對試驗菌株進行處理,并設空白對照,測定供試菌株的胞外多糖、細胞膜滲透性和生物大分子 (DNA、RNA、蛋白質和胞外酶活性)。結果顯示:紅谷霉素對金黃色葡萄球菌的抑制作用首先表現在對核酸合成的抑制,由于核酸的合成受阻,引起菌體內的蛋白質和其它的生物大分子的合成受阻,細菌細胞膜通透性改變。透視電鏡照片顯示:紅谷霉素處理后金黃色葡萄球菌菌體內部的原生質體明顯變稀。
紅谷霉素;金黃色葡萄球菌;抑菌機制
Abstract:Honggumycin is a kind of new antibiotic.UsingStaphalococcus aureusas the experimental bacterial,the antibacterialmechanis m of Honggumycin was investigated.The experimental bacterialwas treated with Honggumycin at the concentration of EC50and EC90,with no Honggumycin as blank control.And the EPS,plas malemma transparence,and biomolecular(DNA,RNA,and protein)were determined.The results showed that the antibacterial activity of Honggumycin toS.aureuswas presented in inhibiting the synthesis of nuclei acid.The synthesis of protein and other biomolecular were restrained due to the inhibition of nuclei acid synthesis,and the plas malemma permeability was changed.Electron microscope photograph showed the protoplast ofS.aureushad become thin after treatingwith Honggumycin.
Key words:Honggumycin;Staphalococcus aureus;antibacterialmechanis m
江西農業大學生物科學與工程學院應用微生物研究室在以棉花枯萎病菌為靶目標開展農抗生產菌的分離篩選研究中,從土壤中分離篩選到一株鏈霉菌,命名為鏈霉菌 702。其產生的生物活性物質對棉花枯萎病菌有較強的抑菌活性,進一步測定它的抗菌譜,發現其所產生生物活性物質既能抑制革蘭氏陽性細菌,又能抑制革蘭氏陰性細菌,同時還能抑制霉菌和酵母菌[1]。毒理實驗表明其為無毒物質,而且通過對鏈霉菌 702生物活性物質的穩定性研究測定表明其對熱和紫外線穩定,在 pH3~12條件下穩定[2]。這些初步研究結果已顯示了其作為防腐劑的廣闊前景。鏈霉菌 702所產抗細菌組分經紫外、紅外、質譜、核磁共振和 X衍射等測定,結果表明該活性組分與放線菌素 X2同質,為國內首次發現,命名為紅谷霉素。本論文在總結前人工作的基礎上,以金黃色葡萄球菌為試驗菌株,探索紅谷霉素對 G+金黃色葡萄球菌的營養體的抗菌作用性質。通過紅谷霉素對金黃色葡萄球菌的抗菌活性和毒力測定,探索紅谷霉素的抑菌能力,為紅谷霉素的應用提供基礎數據資料。
1 材料和方法
1.1 材料
1.1.1 紅谷霉素純品 (含量 99.47%)
由江西農業大學生物工程院提供。
1.1.2 抑菌測定指示菌
金黃色葡萄球菌(Staphalococcus aureus)
1.1.3 培養基[3]
①指示菌斜面培養基:牛肉膏蛋白胨培養基;②指示平板培養基:GPP培養基。
1.2 方法
1.2.1 紅谷霉素對金黃色葡萄球菌DNA作用影響的測定
1.2.1.1 DNA標準曲線的測定
取 6支試管,按表 1添加試劑。混勻后于 60℃恒溫水浴保溫 60 min,冷卻后以 0號管作對照,于595 nm處測定A595,以DNA濃度為橫坐標,光吸收值為縱坐標,繪制標準曲線,見表 1。
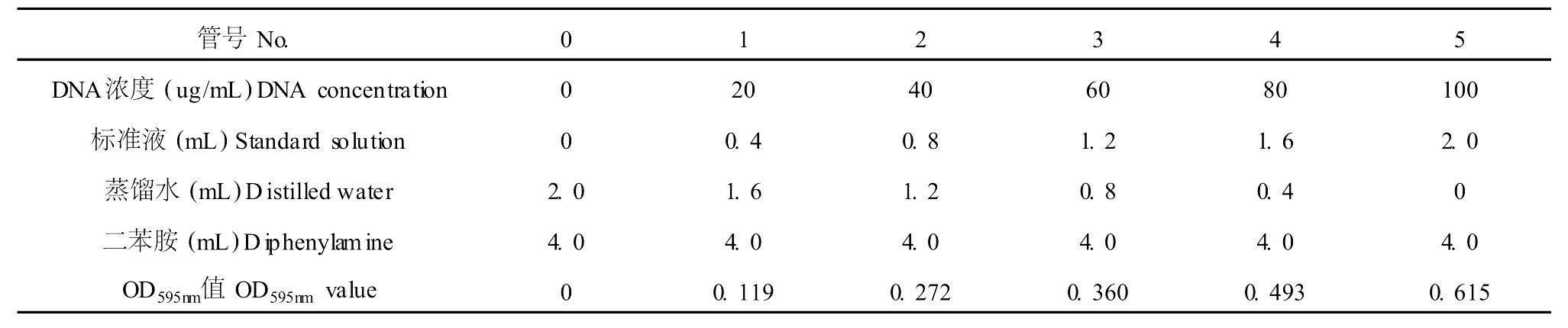
表1 DNA標準曲線的測定Table 1 The determination ofDNA specification curve
1.2.1.2 供試樣品中的DNA含量測定
DNA的含量測定采用改良的二苯胺法。將供試菌在 GPP培養液中振蕩培養至對數生長期,再分為 3瓶,處理紅谷霉素,使菌液紅谷霉素的最終濃度為 EC90、EC50、對照加無菌水,30℃ 120 r/min繼續培養;分別于加藥前、加藥后 1、3、6 h取樣。然后采用有機溶劑抽提法分離提取樣品中 DNA,將上層的水相和下層的有機溶劑層分開保存。取上層水相按DNA標準曲線測定方法進行DNA含量的測定。
1.2.2 紅谷霉素對金黃色葡萄球菌 RNA作用影響的測定
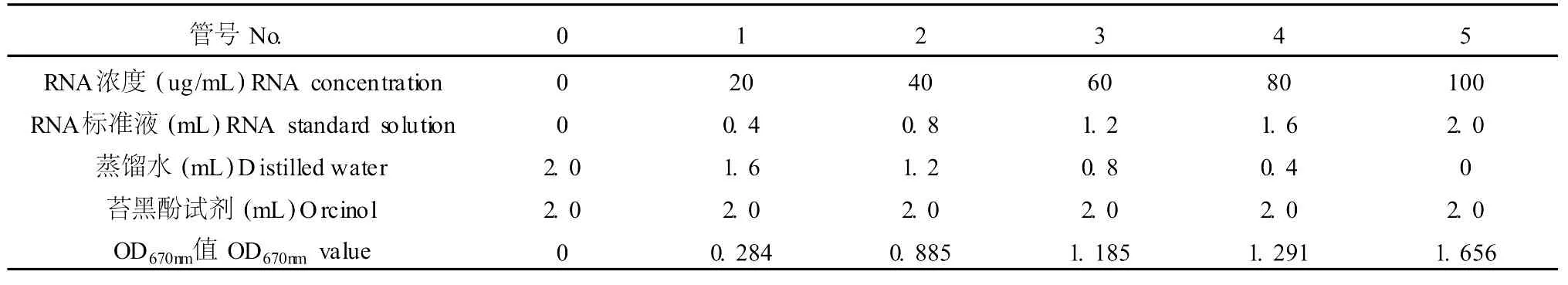
表2 RNA標準曲線的測定Table 2 The determination of RNA specification curve
1.2.2.2 供試樣品中 RNA含量的測定
RNA含量測定采用改良苔黑酚法。將供試菌在 GPP培養液中振蕩培養至對數生長期,再分為 3瓶,處理紅谷霉素,使菌液紅谷霉素的最終濃度為EC90、EC50、對照加無菌水,30 ℃,120 r/min繼續培養;分別于加藥前、加藥后 1、3、6 h取樣。然后采用熱酚法提取樣品中 RNA,再按 RNA標準曲線測定方法進行測定。
1.2.3 紅谷霉素對金黃色葡萄球菌蛋白質作用影響的測定
1.2.3.1 蛋白質標準曲線的測定
取 6支試管,按表 3添加試劑。混勻后放置 2 min后,在 595 nm下比色,繪制標準曲線,見表 3。
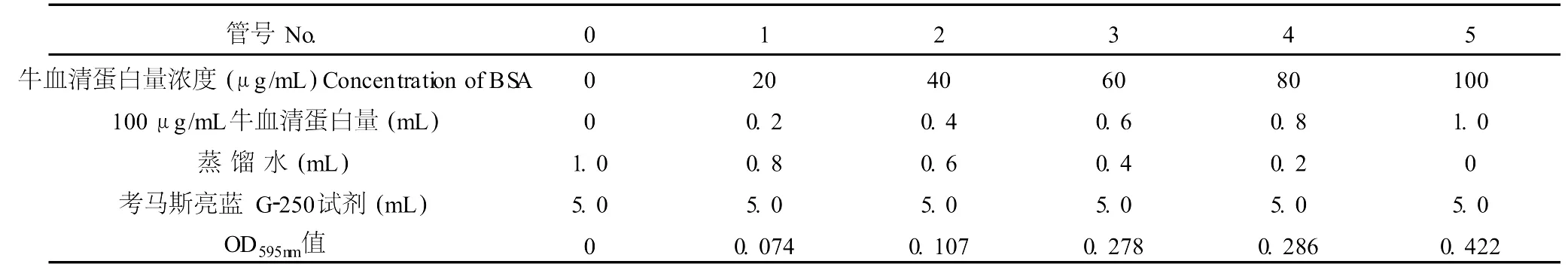
表3 蛋白質標準曲線的測定Table 3 The deter mination of protein from zero to a hundredμg/mL specification curve
1.2.2.1 RNA標準曲線的測定
取試管 6支,按表 2添加試劑。混勻后置 100℃恒溫水浴保溫 45 min,以 0號管為對照,于 670 nm處測定光吸收值 A670,以 RNA濃度為橫坐標,光吸收值為縱坐標作圖,繪制標準曲線,見表 2。
1.2.3.2 供試樣品中蛋白質含量的測定
蛋白質測定采用考馬斯亮藍法。將供試菌在GPP培養液中振蕩培養至對數生長期,再分為 3瓶,處理紅谷霉素,使菌液紅谷霉素的最終濃度為EC90、EC50、對照加無菌水,30 ℃,120 r/min繼續培養;分別于加藥前、加藥后 1、3、6 h取樣。然后采用有機溶劑抽提法分離提取樣品中蛋白質,再按蛋白質標準曲線測定方法進行測定。
1.2.4 紅谷霉素對供試菌體胞外多糖 (EPS)作用影響的測定[4]
將 0.1 mL菌體懸浮液分別均勻涂布于含紅谷霉素低于有效抑制中濃度 EC50系列濃度的 GPP平板上,28~30℃培養 2 d,收集菌落,用定量的無菌水洗下 EPS,離心上清液用粘度計算其粘度,菌體烘干后稱量,計算相對粘度值 (每毫升上清液粘度 /每毫克菌體干重),并比較紅谷霉素對菌體 EPS生物合成和菌體生長速度速率的影響。
1.2.5 紅谷霉素對菌體細胞膜滲透性作用影響的測定
將供試菌培養至對數生長中期,離心收集菌體,用 1%NaCl溶液洗滌離心,重復 2次,再將菌體懸浮于 1%NaCl溶液中,加紅谷霉素,使最終濃度為供試細菌的 EC50處理,置于 30℃條件下,分別于 0、3、6、12 h,取樣離心,收集上清液,用紫外分光光度計測 220~380 nm間的吸收峰型及 260 nm吸收物含量變化。
1.2.6 紅谷霉素對菌體作用前后菌體形態的變化的測定
挑取在平板上分離較好的單個菌落接種于 GPP液體培養基中,于 37℃過夜培養,進行擴大培養。再轉接于新鮮的液體培養基中培養至對數生長期,加入稀釋好的紅谷霉素,使培養液中紅谷霉素的濃度為 10倍殺菌濃度,分別取加藥前和加藥后的 3 h的菌液,離心收集菌體。
收集處理前后的菌體,用 0.1 mol/L的磷酸緩沖液洗滌菌體兩次,棄緩沖液,加入 2.5%戊二醛固定液,混勻菌體,于 4℃固定過夜;離心收集菌體,加入溶解后冷卻至 50℃左右的 1.5%的瓊脂糖,用牙簽輕輕將沉淀挑起來,確保菌體全部包埋進瓊脂糖中。待凝固后,用刀片切去多余的部分,把有樣品的留下,切成小長條,重新放入固定液中。
用 0.1 mol/L的磷酸緩沖液沖洗樣品 6次,每次 20 min;然后用 1%鋨酸 4℃冰箱中固定 2 h,再用 0.1 mol/L的磷酸緩沖液沖洗樣品 6次,每次 10 min;再經 50%、70%、80%、90%、100%(含 CuSO4)的酒精依次脫水,每次 10 min;再用環氧丙烷、環氧丙烷 +硫酸銅各處理 10 min;環氧丙烷:環氧樹脂 3∶1,處理 1 h;環氧丙烷 ∶環氧樹脂 1∶1,處理 2 h;環氧丙烷∶環氧樹脂 1∶3,處理 30 min;環氧樹脂保存過夜。
最后用 Epon818樹脂包埋、超薄切片、染色。制好的樣品在透射電鏡下觀察并記錄結果、拍照。
1.2.7 紅谷霉素對細菌產胞外酶作用影響的測定
胞外酶活性測定采用酶液瓊脂擴散法進行[5]。基本方法是:在 GPP培養基中加入相應底物,滅菌后待培養基冷卻至 50℃左右后加入不同濃度的紅谷霉素 1 mL,搖勻倒平板,然后點滴稀釋好的菌體懸液 0.1 mL,涂布均勻,30℃培養 3 d,向培養皿中加入相應酶的顯色劑顯現酶水解圈。據水解圈與菌落直徑的比值,了解各紅谷霉素濃度條件下菌株胞外酶的活性。
淀粉酶:底物為 0.25%可溶性淀粉,顯色劑為Lugol’s碘液 (I21g,KI2g,H2O 300 mL)。
蛋白酶:底物為 1%明膠,顯色劑為飽和硫酸銨。
脂酶:底物為 1%的吐溫 20,不用顯色劑。
2 結果和分析
2.1 紅谷霉素對金黃色葡萄球菌DNA的作用影響的測定結果
2.1.1 DNA標準曲線
根據表 1測定 DNA標準曲線結果,如圖 1所示。
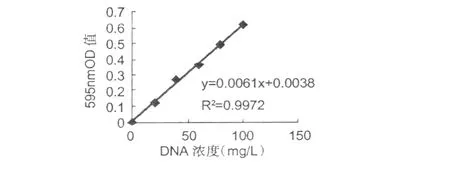
圖1 DNA標曲Fig.1 Standard curve ofDNA
2.1.2 紅谷霉素對金黃色葡萄球菌DNA的作用影響的測定結果
紅谷霉素對金黃色葡萄球菌 DNA的作用影響的測定結果,如圖 2所示。
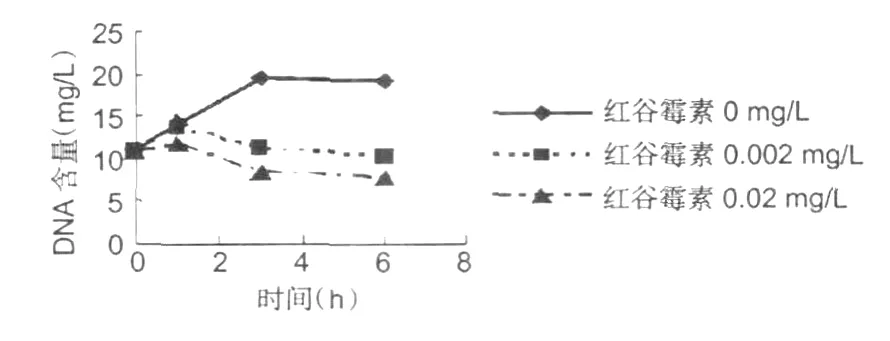
圖2 金黃色葡萄球菌 DNA含量變化Fig.2 The change ofS.aureuDNA
從圖 2可以看出,在紅谷霉素對供試菌體的DNA有強烈地抑制作用。隨著紅谷霉素濃度的增加和作用時間的延長,細菌的 DNA含量在迅速地減少。
2.2 紅谷霉素對金黃色葡萄球菌DNA的作用影響的測定結果
2.2.1 RNA標準曲線測定結果
根據表 2測定 RNA標準曲線結果,如圖 3所示。

圖3 RNA標曲Fig.3 Standard curve of RNA
2.2.2 紅谷霉素對金黃色葡萄球菌DNA的作用影響的測定結果
紅谷霉素對金黃色葡萄球菌 DNA的作用影響的測定結果,如圖 4所示。
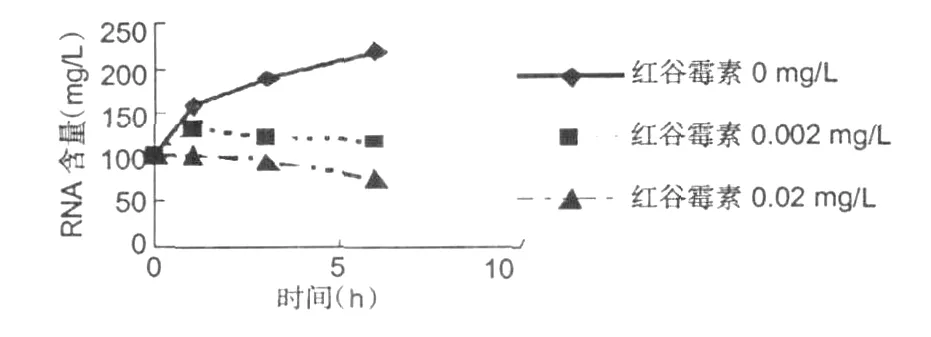
圖4 金黃色葡萄球菌 RNA含量變化Fig.4 The change ofS.aureuRNA
從圖 4可以看出在紅谷霉素對供試菌體的RNA有強烈地抑制作用。隨著紅谷霉素濃度的增加和作用時間的延長,細菌的 RNA含量在迅速的減少。
2.3 紅谷霉素對金黃色葡萄球菌蛋白質的作用影響的測定結果
2.3.1 蛋白質標準曲線的測定結果
根據表 3測定蛋白質標準曲線結果,如圖 5所示。
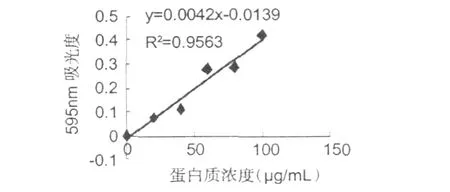
圖5 蛋白質標曲Fig.5 Standard curve of protein
2.3.2 紅谷霉素對金黃色葡萄球菌蛋白質的作用影響的測定結果
紅谷霉素對金黃色葡萄球菌蛋白質的作用影響的測定結果,見圖 6所示。
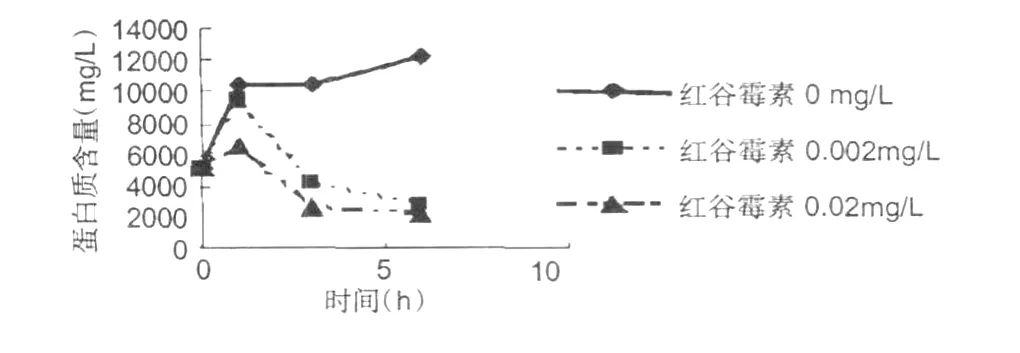
圖6 金黃色葡萄球菌蛋白質含量變化Fig.6 The change ofS.aureuprotein
從圖 6可以看出,紅谷霉素對供試菌體的蛋白質有強烈地抑制作用。隨著紅谷霉素濃度的增加和作用時間的延長,細菌的蛋白質含量在迅速的減少。
2.4 紅谷霉素對金黃色葡萄球菌胞外多糖 (EPS)作用的測定結果
根據紅谷霉素在 0、0.0025、0.005、0.01 mg/L和 0.02 mg/L五個濃度下對葡萄球菌作用 48 h后的相對粘度值 (每毫升上清液粘度/每毫克菌體干重),并將各相對粘度值與紅谷霉素濃度作圖,結果如圖 7所示。
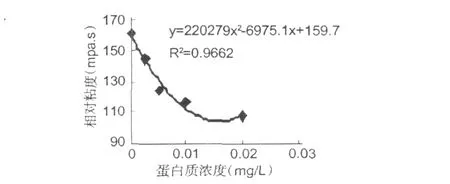
圖7 紅谷霉素濃度與金葡菌菌體粘度關系Fig.7 The relate of Honggumycin chroma andS.aureuviscosi
從圖 7的結果可以看出,隨著紅谷霉素濃度的增加,金黃色葡萄球菌的胞外多糖含量相應的減少,說明紅谷霉素對細菌胞外多糖的產生會有抑制作用。
2.5 紅谷霉素對菌體細胞膜滲透性的作用的測定結果
加入紅谷霉素 EC50濃度作用于金黃色葡萄球菌,置于 30℃條件下培養,分別在培養 0、3、6、12 h,取樣離心取上清液,用紫外分光光度計測吸收物含量變化,結果分別見圖 8~11。
從圖 8~11可以看出,經紅谷霉素處理過的金黃色葡萄球菌隨處理時間的增加,吸光度越大,說明胞外生物大分子含量增加。因此,可推斷紅谷霉素作用細胞膜的蛋白質合成,導致細菌細胞膜上膜蛋白含量的減少,從而造成細胞膜通透性增加。
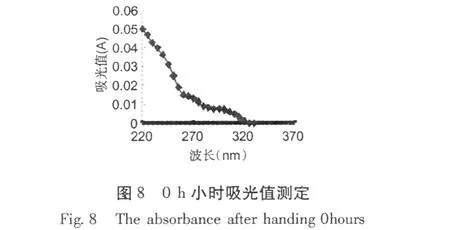
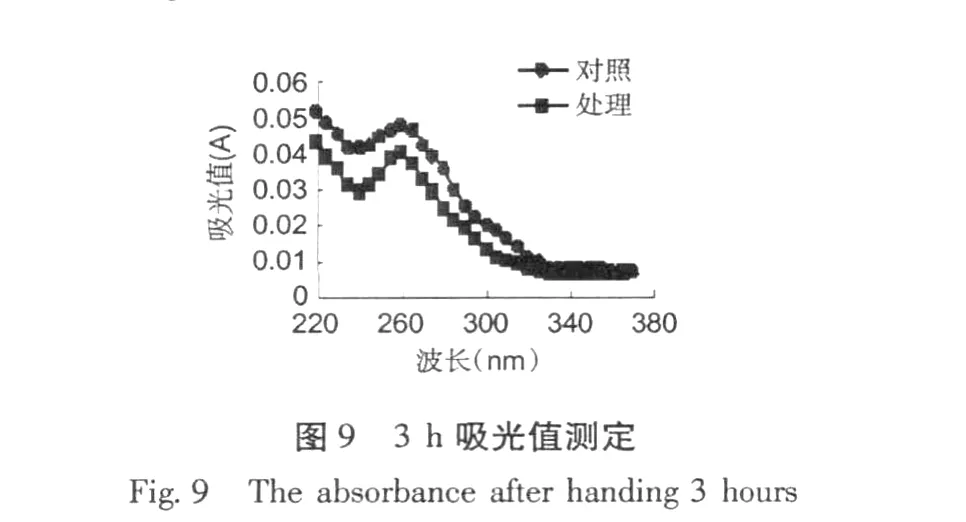
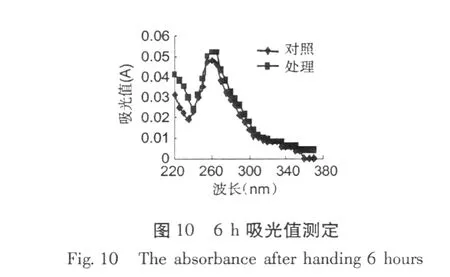
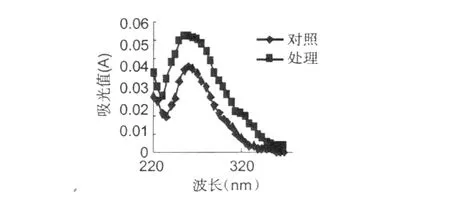
圖11 12 h吸光值測定Fig.11 The absorbance after handing 12 hours
2.6 紅谷霉素對金黃色葡萄球菌作用前后體形態變化的測定結果
紅谷霉素對金黃色葡萄球菌作用前后菌體形態變化的透射電鏡照片見圖 12、13。
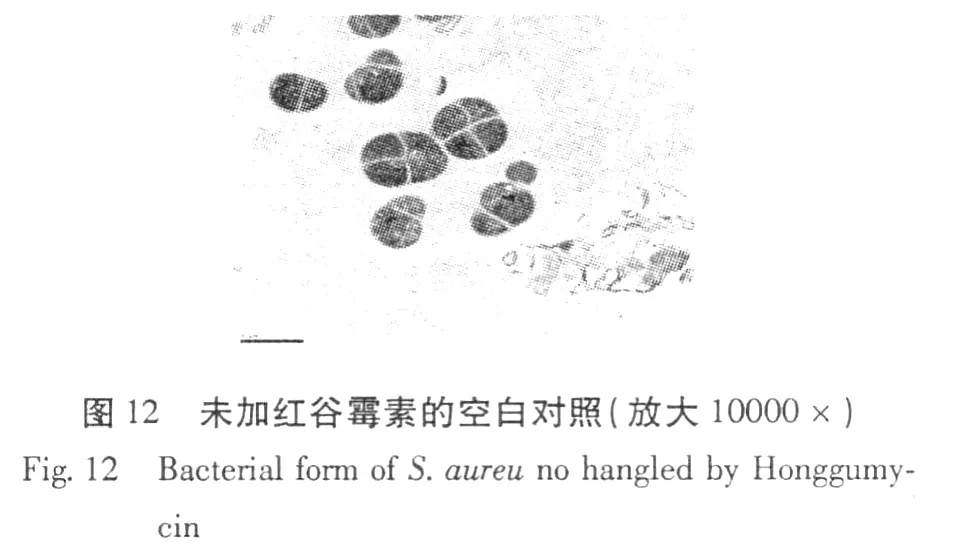
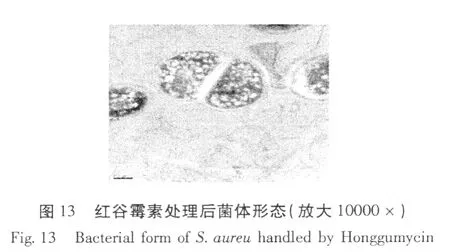
從圖 12、13的結果可以看出,用紅谷霉素處理后的細菌,其內部原生質體與對照相比明顯的變稀。紅谷霉素作用后菌體形態的變化進一步說明紅谷霉素對金黃葡萄球菌的抑菌機制是抑制菌體生物大分子 DNA、RNA和蛋白質的合成。
2.7 紅谷霉素對金黃色葡萄球菌胞外酶作用影響的測定結果
紅谷霉素處理前后金黃色葡萄球菌產胞外酶能力的比較結果如表 4所示:

表4 紅谷霉素對金黃色葡萄球菌的胞外酶產生的影響Table 4 The activity of extracellullar enzymes ofS.aureu
從表 4可以看出,在培養基中加入紅谷霉素后,細菌的胞外淀粉酶、蛋白酶和脂酶產量有明顯的降低,這進一步說明紅谷霉素對細菌蛋白質的合成有強烈的抑制作用。
3 討論
3.1 紅谷霉素對金黃色葡萄球菌的 DNA、RNA、蛋白質、胞外多糖和胞外淀粉酶、蛋白酶、脂酶的合成有強烈地抑制作用。隨著紅谷霉素濃度的增加和作用時間的延長,細菌的 DNA、RNA、蛋白質、胞外多糖和胞外淀粉酶、蛋白酶、脂酶含量在迅速地減少。說明紅谷霉素對金黃色葡萄球菌的作用機制是首先抑制細胞內 DNA、RNA的合成,從而導致細胞內外蛋白質的減少和細胞膜的滲透性增大。
3.2 紅谷霉素處理前后金黃色葡萄球菌菌體形態的透射電鏡照片表現出明顯的變化,這從菌體的形態學上進一步佐證了紅谷霉素對金黃色葡萄球菌的作用機制。
3.3 在以紅谷霉素為試驗材料,分別以 G+菌,臘質芽孢桿菌、巨大芽孢桿菌和 G-菌,大腸桿菌和小麥青枯病為供試菌進行抑制機制的研究,獲得了與金黃色葡萄球菌同樣的試驗結果,進一步說明紅谷霉素對細菌的作用機制是抑制細菌細胞內 DNA和RNA的合成,從而導致影響細胞內外蛋白質的合成。本試驗獲得的紅谷霉素對金黃色葡萄球菌的作用機制有待于分子生物學上進一步試驗和驗證。
1 Li KT,Li XH,Liu SH,et al.A prel iminary study on the effects of bio-antiseptic 702 on anti-bacteria,anti-moulds and anti-yeasts.Acta Agric Univ Jiangxiensis,2002,24:599-602.
2 Cai HJ,Wen Y,Tu GQ.Measure of the stability of the bioactive compound produced byStreptom yces702.Acta AgricUniv Jiangxiensis,2003,25:929-933.
3 Shen P,Fan XR,Li G W,et al.Microbiology Experments.Beijing:Higher Education Press,1999,18:214-223.
4 Huang YL,Hu ML. Improvement on cell membrane permeability experiment.J Zhanjiang No rm al Coll,2005,26:119-122.
5 Dingle J,RE ID WW,Solomons GL.The enzymic de gradation of pection and otherpolysaccharides.J Sci Food Agric,1953,4:149-155.
Antibacterial Mechanism of Honggumycin toStaphylococcus aureus
TU Xuan,XUE Xiu-yuan,TU Guo-quan*
B iological Science&Engineering College,Jiangxi Agricultural University,Nanchang 330045,China
Q939;R446.5
A
1001-6880(2011)01-0153-06
2009-09-04 接受日期:2009-11-30
江西省自然科學基金項目(0630013)
*通訊作者 E-mail:tuguoquan@263.net
