品種混種方式對大豆胞囊線蟲控制作用
2012-01-11 10:29:30,,2,,,,,,,4
土壤與作物 2012年2期
關鍵詞:大豆
, ,2,,,,,, ,4
(1.中國科學院 東北地理與農業生態研究所 黑土區農業生態院重點實驗室,海倫農田生態系統國家野外觀測研究站, 黑龍江 哈爾濱 150081;2.大興安嶺地區農林科學院, 黑龍江 加格達奇 165000;3.黑龍江省農業科學院 大慶分院, 黑龍江 大慶 163316;4.黑龍江農墾九三管理局 植保植檢站,黑龍江 嫩江 161441)
大豆是我國北方主要作物,以品質好而著稱,但生產中大豆胞囊線蟲(Soybean cyst nematode,SCN)病嚴重影響其產量[1],該病也是全球大豆生產的重要病害[2]。多年來人們一直在不斷地尋找最佳防治措施,由于大豆胞囊線蟲以抗逆性較強的胞囊形式多年存活于土壤中,給防治帶來極大的困難,最經濟有效的控制方法為與禾本科作物輪作[3-4]以及應用抗病品種。由于大豆經濟價值較高,長期輪作難以實現;抗病品種可以控制土壤中SCN胞囊數量,減輕SCN病害的發生,我國近年也審定了一些抗性較好的品種[5],生產中應用面積在逐步擴大,但由于抗病品種在外觀上和適應地區有一定局限性,大面積應用在近期還不容易做到。生產上如何發揮這些抗線蟲品種的作用、有效地減少線蟲危害是目前大豆生產中急需解決的問題。
利用作物品種遺傳多樣性可有效地控制植物病害,還可同時提高作物產量。作物種內多樣性在病害控制上的應用包括多系品種和品種混合兩種形式[6-7]。在水稻和小麥等作物上有許多成功的例子[8-10]。水稻上應用主要是采用抗感品種混栽控治水稻稻瘟病[11]。也有研究發現小麥品種混種可減少小麥白粉病、葉銹病和條銹病發病害,但對病害控制效果也因品種等因素存在差別[12],如小麥條銹病防治效果的變化幅度在13 %~97 %[13]之間。以上研究都是利用品種多樣性來控制空氣傳播的病害,但是應用于土傳病害的研究鮮為報道。為了探討利用品種多樣性控制大豆胞囊線蟲病的可能性,在北方大豆產區開展了利用抗大豆胞囊線蟲品種與優質高產品種混合種植控制大豆胞囊線蟲病研究,旨在探討土傳病害控制新途徑。試驗在黑龍江省中部的黑土區進行,采用北方抗大豆胞囊線蟲病品種與當地生產中廣泛應用的優質豐產品種黑農進行混種,探討混種方式對大豆胞囊線蟲病的控制作用以及對大豆生長發育和產量的影響。
2 材料與方法
2.1 試驗地點及方法
試驗地點在黑龍江省中部大豆主產區,位于海倫市中國科學院海倫農業生態試驗站,中心地理位置為東經126°38′,北緯47°26′。茬口為15 a大豆連作和大豆迎茬(大豆-玉米-大豆)。大豆胞囊線蟲自然發病區,土壤中大豆胞囊線蟲經鑒定為3號生理小種,土壤類型為中厚黑土。
試驗區隨機排列,3次重復。小區長5 m,寬3.5 m,6壟。人工播種,3鏟3趟。大豆施肥量:磷酸二銨150 kg· hm-2;K2SO4(K2O含量50%)75 kg·hm-2。種植密度30萬株·hm-2。
2.2 大豆品種
2.2.1大豆胞囊線蟲抗性品種。抗線4號(KX4),由黑龍江省農業科學研究院大慶分院提供。
品種特性:亞有限結莢習性,株高70 cm左右,圓葉,白花,灰毛。圓粒,褐臍,百粒質量20 g~22 g,高抗大豆胞囊線蟲3號生理小種,稈強,蛋白質含量38.20 %,脂肪20.77 %,生育日數113 d左右,平均產量2 300 kg·hm-2左右[14],該品種適于黑龍江省和內蒙東部地區種植,耐鹽堿和干旱[15]。
2.2.2大豆胞囊線蟲非抗性品種。黑農35號(HN35),由黑龍江省農業科學研究院大豆研究所提供。
品種特性:高蛋白品種,蛋白質含量45.24 %,脂肪含量18.36 %,亞有限結莢習性,植株高度為80 cm~85 cm,亞有限結莢習性,主莖發達,少分枝,白花,長葉,生育日數115 d左右。因其蛋白質含量高,豐產性好,是黑龍江省中部大豆主產區主栽品種[16]。
2.3 試驗處理
抗性與非抗性品種混種方式。試驗共設置6個處理,見表1,分別設置15 a大豆連作和大豆迎茬兩不同茬口。
2.4 大豆植株根冠比測定
取樣方法:大豆出苗后開始取樣,每處理取大豆10株,3次重復,保持根系完整。每7 d取1次,共取6次。
大豆植株用清水沖洗掉根部殘余土壤,自然晾干、稱質量,計算大豆植株根冠比。
2.5 土壤中大豆胞囊線蟲二齡幼蟲測定
取樣方法:大豆出苗后開始取樣,去掉表土,拔出大豆根,取5 cm~20 cm耕層大豆根圍土。每小區取樣20個點,每點取約100 g,將所有土樣混合,用四分法取500 g,放入新塑料袋中封好以保濕運回實驗室,放在4 ℃冰箱中保存待用。每7 d取1次樣,共取6次。
2.5.1大豆胞囊線蟲二齡幼蟲分離。大豆胞囊線蟲二齡幼蟲分離采用蔗糖濃度梯度離心法。取出冰箱中保存的土樣,每份稱取100 g置于250 ml燒杯中,加水150 ml浸泡1 h并轉移到1 000 ml的塑料燒杯中,離心機1 750 rpm攪拌3 min,用強水流沖洗土樣混濁液,靜止30 s后過80目和500目套篩。收集500目篩上物于50 ml離心管中,2 000 rpm離心3 min,棄上清,再加入45 ml 615 g·L-1蔗糖溶液,充分混勻,500 rpm離心3 min,上清液過500目篩。用水輕輕沖洗篩上物,洗去殘留的蔗糖,沖洗收集篩上線蟲于50 ml離心管中定容到50 ml后放置于4 ℃冰箱中備用。

表1 抗性與非抗性品種混種方式Tab.1 Mixture treatments of resistant and non-resistant varieties
注:* 抗線4號和黑農35按種子數量比例1∶1混種種植;** 壟內不同單行分別種植抗線4號和黑農35。
2.5.2線蟲計數。取出在冰箱內保存的線蟲收集物,放置于振蕩器上振勻。用移液管吸取5 ml放置于直徑5 cm的培養皿內,在解剖鏡下觀察計數大豆胞囊線蟲2齡幼蟲數量,每處理3次重復。
2.6 土壤中線蟲胞囊及卵計數
取樣方法同2.5。
2.6.1土壤中線蟲胞囊收集。土壤中線蟲胞囊收集采用蔗糖濃度梯度離心法。取出冰箱中保存的土樣,每份稱取100 g置于250 ml燒杯中,加水150 ml浸泡1h并轉移到1 000 ml的塑料燒杯中,離心機1 750 rpm攪拌3 min,用強水流沖洗土樣混濁液,靜止30 s后過20目和80目套篩。收集80目篩上物于50 ml離心管中,2 500 rpm離心3 min,棄上清,再加入45 ml 70 %蔗糖溶液,充分混勻,2 500 rpm離心3 min。上清液過60目篩,用水輕輕沖洗篩上物,洗去殘留的蔗糖,沖洗到放有濾紙的漏斗中,過濾后取出濾紙,收集胞囊。
2.6.2胞囊計數。解剖鏡下觀察挑取的胞囊,并記錄數量。
2.6.3線蟲卵密度測定。取出冰箱中保存的土樣,每份稱取100 g置于250 ml燒杯中,加水150 ml浸泡1h,用濕篩法將土壤樣品中的胞囊篩出,經橡膠塞在200目篩上研磨后,收集200目和500目篩上物于50 ml離心管中,35 %蔗糖溶液梯度2 500 rpm離心3 min,含卵的上清液再過500目篩,清水沖洗后,收集于50 ml離心管中定容為50 ml,將卵懸液保存在4 ℃冰箱中,用以測定大豆胞囊線蟲的卵密度。測定時向塑料小皿(直徑為35 mm)加入1 ml卵懸液,放置于顯微鏡下觀察,記錄卵數量,3次重復,計算卵密度。
2.7 測產考種
各小區內取2 m2用于測產量,連續取10株用于考種。考種項目有:株數·m-2、株高、結莢高度、節數、分枝數、單株莢數、有效莢數、單株粒數、單株粒質量、蟲食率、病粒率等。
2.8 數據分析
試驗結果的方差分析均采用SAS 6.0版本。
3 結果與分析
3.1 品種混種方式對大豆胞囊線蟲控制作用
3.1.1不同混種方式土壤中大豆胞囊線蟲二齡幼蟲變化。在我國北方大豆主產區,胞囊線蟲第一代對生產威脅很大,因此,控制技術對大豆胞囊線蟲第一代的控制效果非常重要。測定土壤中的大豆胞囊線蟲二齡幼蟲(J2)數量,可以在一定程度上明確不同混種方式中病害侵染源的動態變化。
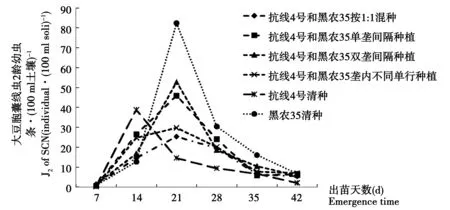
圖1 15年大豆連作田土壤中SCN 二齡幼蟲動態變化Fig.1 The fluctuation of SCN juvenile in 15-year soybean continuous cropping field
在15 a大豆長期連作區,見圖1,所有6個處理的土壤中胞囊線蟲2齡幼蟲數量在調查期內總體變化趨勢相近,即隨著大豆生育進程,在出苗后逐步增加,在大豆出苗后7 d始見二齡幼蟲(J2),14 d開始顯著增加,21 d時達到高峰后逐漸下降。6個處理的土壤中胞囊線蟲2齡幼蟲數量總體比較,在21 d后黑農35清種最高,達到82.4條·(100 ml土壤)-1;抗線4號最低,其它的2個品種間種和混種的4個處理居中,在這些間種和混種方式中以黑農35和抗線4號1∶1混合種植胞囊線蟲2齡幼蟲數量為低。在14 d時抗性品種抗線4號根際土壤內線蟲數量最高,達到39.6條·(100 ml土壤)-1,并且與黑農35凈種和1∶1混種處理間差異顯著。該處理的高峰早于其它處理,其原因可能由于抗線蟲品種根系分泌物對線蟲卵孵化的刺激要早于非抗線品種或抗線4號根系分泌物對大豆胞囊線蟲卵孵化較非抗線蟲品種有更強促進作用所致[17]。出苗28 d后所有處理土壤內J2數量顯著下降,并且持續減少。可能是土壤中有活性的卵已經基本完成孵化,孵化后的J2已經侵入大豆根部或者未能找到寄主而在土壤中死亡。所有處理的土壤中J2數量在出苗后35 d和42 d時達到最低點。在整個過程中,1∶1混種減少了土壤中J2數量,僅高于抗線4號。
大豆迎茬地塊土壤中大豆胞囊線蟲2齡幼蟲數量總趨勢與15 a大豆連作接近圖2,整體J2數量上要高于15 a大豆連作區。土壤內線蟲數量在出苗后14 d開始顯著增加,抗線4號根際土壤內J2數量最高,達到70.3條·(100 ml土壤)-1,并且與黑農35以及1∶1混種處理間差異顯著。其它處理則在出苗后21 d時土壤中J2數量達到最大值,以黑農35清種數量最高,顯著高于各混種處理,混種方式間差異不顯著。而抗線4號的根周圍土壤內的J2數量在21 d后開始下降,低于其它各處理水平。所有處理在出苗28 d后土壤內J2數量明顯下降,到出苗35 d和42 d時達到最低點。各混種處理對大豆胞囊線蟲2齡幼蟲的控制效果接近抗性品種清種。
3.1.2不同混種方式大豆根表雌蟲數量變化。大豆胞囊線蟲雌蟲在發育成熟后體壁加厚發育為胞囊,附著在大豆根表,而后落入土壤中,胞囊中的卵在周圍環境適合時孵化為幼蟲侵入寄主根內,形成再次侵染循環。因此,根表面的雌蟲數量可以在一定程度上表明線蟲繁殖能力的強弱,雌蟲的多少直接影響到下一個世代線蟲的再侵染能力。
15 a大豆連作田中6個處理均為出苗后28 d時在大豆根部始見雌蟲,見圖3。出苗后35 d雌蟲數量達到高峰,其中以黑農35清種數量最多,平均每株大豆根上達到了22.3個,顯著高于其它各混種處理,抗線4號最低,僅為1.5個每株。1∶1混種方式低于抗線4號清種外所有的其它混種方式,各混種方式之間存在一定差異,但差異不顯著。出苗后42 d,所有處理根表面雌蟲數量開始減少,此時胞囊已經成熟,脫落到土壤當中。仍是以黑農35號的根表面的雌蟲數量最高,相當于其它處理根表雌蟲數量的1倍以上。所有混種方式中以1∶1混種處理效果最好,大豆根表雌蟲數量要顯著低于黑農35。
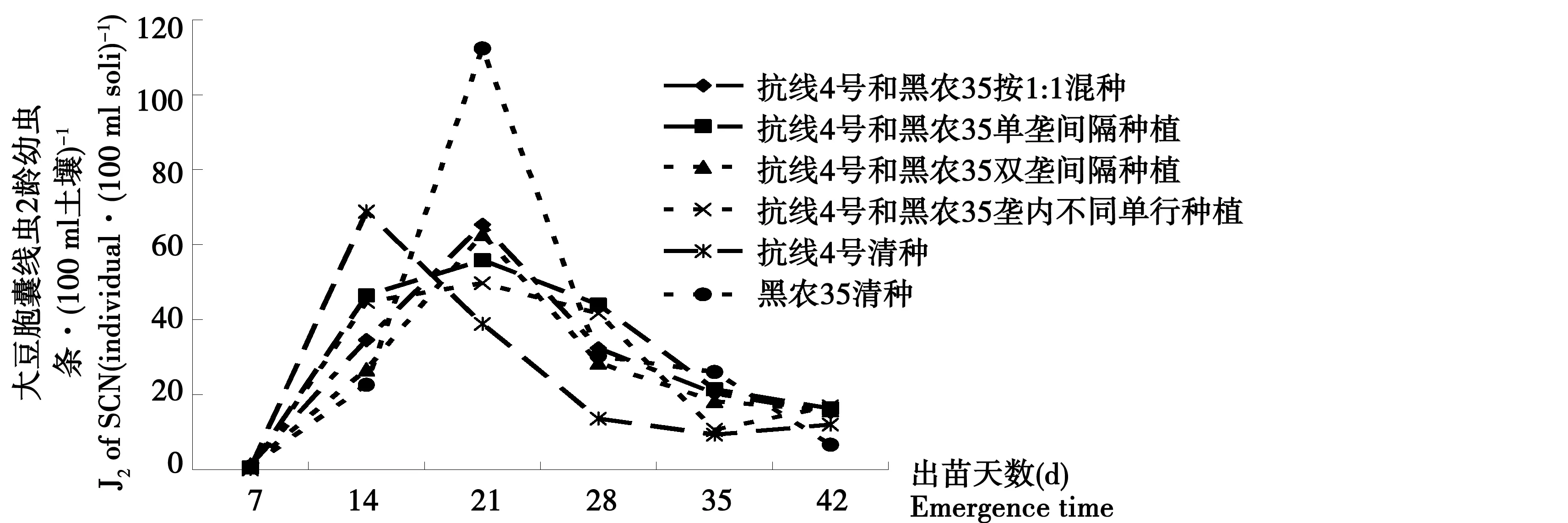
圖2 1年大豆連作田土壤中SCN 二齡幼蟲動態變化Fig.2 The fluctuation of SCN juvenile in corn-soybean continuous cropping field
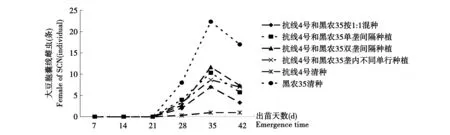
圖3 15年大豆連作田大豆根表SCN雌蟲動態變化Fig.3 The fluctuation of SCN Female on soybean root in 15-year soybean continuous cropping field
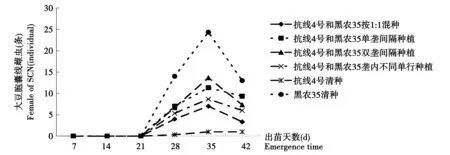
圖4 1年大豆田大豆根表SCN雌蟲動態變化Fig.4 The fluctuation of SCN Female on soybean root in corn-soybean continuous cropping field
1 a大豆迎茬田大豆根部雌蟲數量與15 a大豆連作總體趨勢相同,見圖4。所有處理在出苗后28 d時在大豆根部始見雌蟲,出苗后35 d雌蟲數量達到最高峰,其中以黑農35數量最高,相當于各混種方式的2~4倍,抗線4號最低,1∶1混種方式低于除抗線4號凈種外所有的混種方式,混種方式之間存在一定差異,但差異不顯著。出苗后42 d,所有處理根表面雌蟲數量減少。其中以黑農35號的根表面的雌蟲數量最高,為12.1個每株,1∶1混種處理的根表雌蟲數量接近抗線4號凈種,平均僅為2.6個每株,顯著低于其它混種處理以及黑農35清種。1∶1混種處理在所有混種方式中效果最好,接近抗線4號清種。
3.2 不同混種方式大豆根冠比變化
大豆胞囊線蟲侵染大豆后對大豆根部生物量的影響要大于對地上部的影響,因此也會引起大豆根冠比的變化,通過測定大豆的根冠比可以在一定程度上反映種植方式對大豆植株生長的影響。
15 a大豆連作田中所有處理大豆根冠比隨出苗天數增加而不斷降低,見圖5,在出苗28 d后趨于平穩。出苗7 d時黑農35的根冠比最高,顯著高于其它各處理,抗線4號的根冠比最低,可能是由于品種間差異引起。出苗14 d后由于大豆胞囊線蟲的侵入,非抗性品種的根冠比要低于抗性品種,而1∶1混合種植的根冠比最高,大于兩個清種處理,并存在顯著性差異。出苗21 d起,由于大豆植株的快速生長,使得各處理的根冠比相對于14 d時均發生大幅度降低,但是1∶1混合種植的根冠比仍然明顯高于所有清種處理。并且這一趨勢一直保持到出苗后42 d。因此,1∶1混合種植處理對大豆胞囊線蟲病害發生有較好的控制作用,減輕了線蟲對大豆根部的危害。
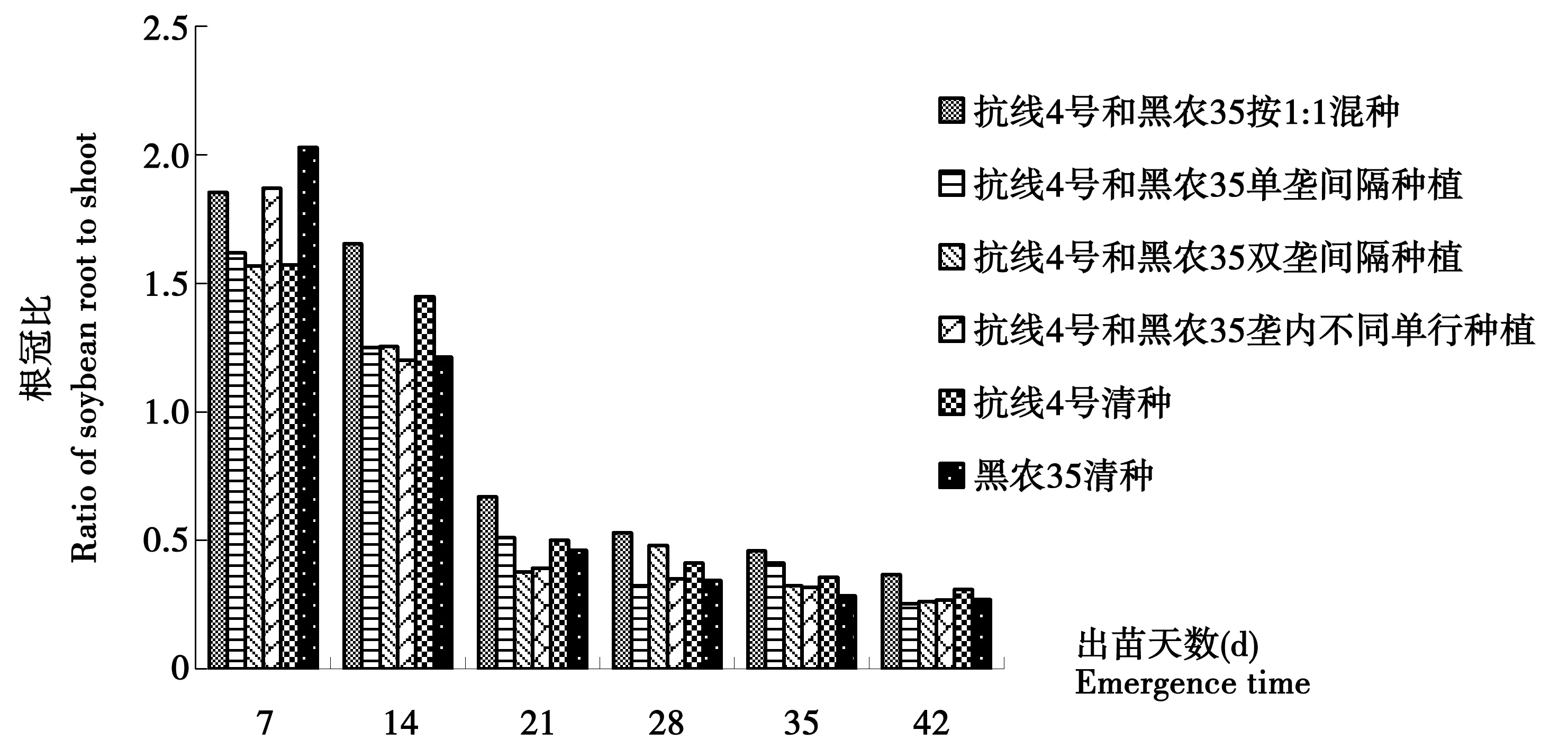
圖5 15年大豆連作田大豆根冠比動態變化Fig.5 The ratio of soybean root to shoot in 15-year soybean continuous cropping field
迎茬田大豆根冠比總體要高于15 a大豆連作,并且整體趨勢相同,見圖6,隨出苗天數增加而不斷降低,在出苗28 d后趨于平穩。出苗7 d時,黑農35清種的根冠比最高,并顯著高于抗線4號,與各混種處理之間存在差異但不顯著。出苗14 d后黑農35的根冠比要低于抗線4號,而1∶1混合種植的根冠比最高,顯著高于黑農35清種。出苗21 d開始,由于大豆植株的快速生長,使得各處理的根冠比相對于14 d時均發生大幅度降低,但是1∶1混合種植的根冠比仍然明顯高于清種處理。28 d后,各處理之間雖存在一定差異,但差異不顯著。

圖6 1年大豆連作田大豆根冠比動態變化Fig.6 The ratio of soybean root to shoot in corn-soybean continuous cropping field
3.3 不同混種方式對大豆產量影響
控制大豆胞囊線蟲病害發生的目的是減輕病害對大豆植株正常生長發育的影響,提高大豆產量,大豆的產量是評價不同混種方式之間是否存在差異的最重要因素之一。
在大豆成熟后對其測產,見表2,結果表明,大豆迎茬田中,以兩品種1∶1混合種植處理的產量最高,雙壟間隔種植產量最低。黑農35的產量要低于抗線4號,可能由于大豆胞囊線蟲病害的影響。1∶1混種較黑農35號清種增產8.8 %,差異顯著,但與抗線4號差異不顯著,較其增產2.8 %。15 a連作與迎茬相比總體產量較低,大豆產量以1∶1混合最高,單壟間隔種植最低, 1∶1混種較黑農35號清種增產8.2 %,差異顯著,較抗線4號清種增產7.4 %,但無顯著性差異。無論在15 a連作還是迎茬條件下單壟和雙壟間隔種植產量都不高,明顯不如混合種植。品種搭配以1∶1混合種植增產效果明顯。
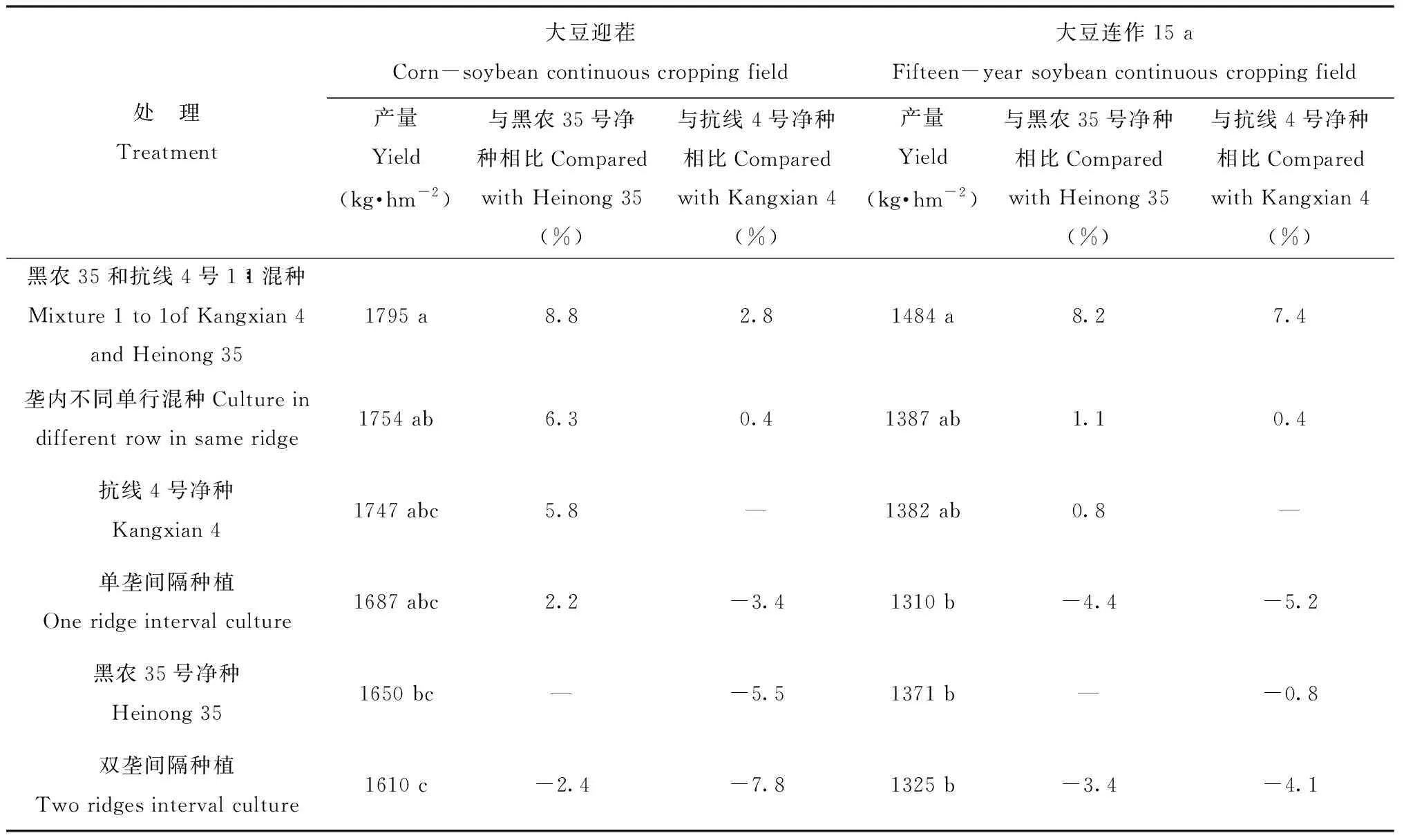
表2 大豆迎茬和15年連作田不同混種方式大豆產量Tab.2 Soybean yield under different mixture cultivar model in corn-soybean and 15-year soybean continuous cropping fields
3.4 混種方式對后茬大豆胞囊線蟲侵染源影響
大豆胞囊線蟲是土傳性病害,病原物以胞囊的形式在土壤中越冬,在第二年春季胞囊中的卵孵化后對春播大豆進行侵染,引起病害發生[18]。控制大豆胞囊線蟲在秋季收獲時的種群數量就可以有效的降低第二年病害發生的侵染來源,使病害發生得到持續的控制,因此收獲后土壤中線蟲卵的數量致關重要。
大豆收獲后對土壤中線蟲卵數量調查見表3。大豆迎茬田中,種植黑農35后線蟲種群數量明顯著增加,相當于播前數量的3倍左右。抗線4號清種處理線蟲種群數量降低,低于播種前水平,卵數量相當于播種前水平的20 %。各混種方式線蟲種群數量較播種前均有所降低,但差異不顯著。以1∶1混種植降低最多,相對于播前減少了30 %,強于其它混種方式,對大豆胞囊線蟲后茬侵染源具有較強的控制作用。15 a大豆連作田中收獲后土壤中大豆胞囊線蟲卵數量總體上要低于大豆迎茬田,分析原因可能是大豆長期連作后土壤中產生了抑制大豆胞囊線蟲的天敵,如真菌[19]。黑農35清種在收獲后線蟲數量明顯增加,與播種前相比差異顯著,是播前的3倍以上。抗線4號線蟲數量明顯降低,與播種前差異顯著,卵數量減少到播前的30 %以內。各混種方式大豆胞囊線蟲的種群數量較播種前有所降低,其中按1∶1混種種植的卵數量與播前差異顯著,降低到了播前水平的50 %。其它混種方式均有所降低,但未達到顯著差異水平。1∶1混種對線蟲的控制作用15 a連作田效果要好于迎茬地塊,在所有混種方式中效果最好。
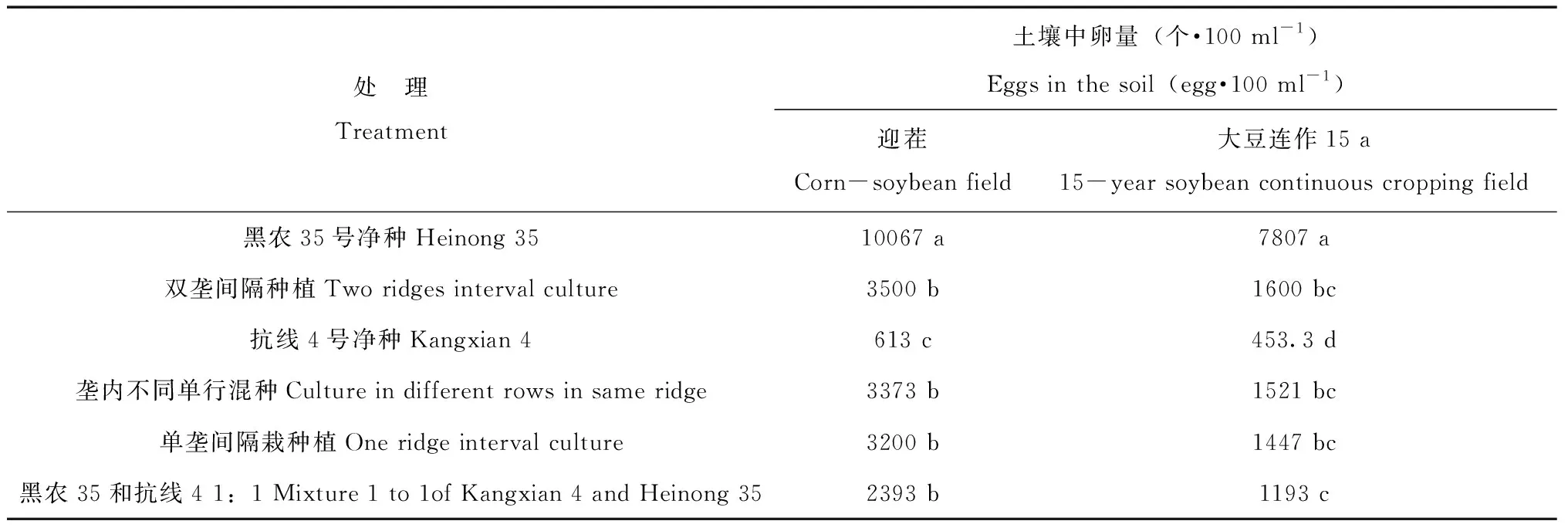
表3 迎茬和15年大豆中不同混種方式收獲后土壤中SCN卵數量Tab.3 The eggs of SCN under different mixture cultivar models in 2-year and 15-year soybean continuous fields after harvest time
4 討 論
通過抗性和非抗性線蟲品種清種、不同間隔方式和混合等種植方式比較,對大豆胞囊線蟲發病情況的調查以及對大豆產量的測定,明確了利用抗性品種與非抗性品種進行種子混合種植為最佳的方式,對大豆胞囊線蟲病害發生的控制能力以及增產效果均要優于成行間隔種植和清種。
利用生物的遺傳多樣性控制植物病害,是應用了種植管理和抗病品種等多種方法的綜合控制措施,目前生產上主要是應用于控制水稻稻瘟病以及小麥銹病等空氣傳播的病害,取得了較好的防治效果[20]。氣傳病害的特點是病原微生物隨空氣的流動而在田間擴散傳播,從而引起病害的暴發流行。因此在利用作物的遺傳多樣性控制此類病害發生時,所采用的種植方式多為將抗性品種與非抗性品種成行的間隔種植[21]。其主要原因可能是在非抗性品種中間隔種植抗性品種可以形成有效的病原微生物阻隔帶,使其難以在田間順利地隨氣流傳播,達到控制病害蔓延流行的目的。而且在大多數情況下,非抗性品種的品質較好經濟價值高,成行間隔種植有利于與抗性品種分開收獲。水稻種植一般為旱地育苗后在水田內插秧,成行種植有利于機械耕作,節約勞動成本[22]。
大豆胞囊線蟲病為土壤傳播病害,發病田中的病原生物在大豆種植前已經存在于土壤中,而且大豆胞囊線蟲自身雖然可以在土壤中運動遷移,但是運動距離十分有限,一般水平距離在1 m之內,垂直距離在30 cm之內。對利用作物遺傳多樣性控制土壤傳播病害的種植方式進行了研究,結果表明,將抗性品種與非抗性品種進行單壟或雙壟的成行間隔種植并不能取得很好的控制胞囊線蟲病效果,可能是胞囊線蟲無法憑借自身運動進行壟與壟之間的移動傳播。而對于在同壟內不同單行中分別種植抗性品種和非抗性品種,可能是由于抗胞囊線蟲品種和非抗線蟲品種種植不如1∶1混合種植均勻,非抗性品種土壤中胞囊線蟲仍然可以像清種一樣正常繁殖,胞囊線蟲在第二代時可以沿著非抗性品種種植單行的一側進行二次侵染。在將抗性品種與非抗性品種種子混合后種植的處理中,由于種子是完全混合后播種,抗性品種可以在非抗性品種之間隨機形成有效的抗性阻隔帶,使胞囊線蟲無法憑借自身運動在壟內自由移動,降低了病害的傳播速度,并且有效地控制了第二代線蟲對非抗性品種的再侵染程度。由于在大豆生產上多為大型機械直接播種,種子混合處理與成行間隔相比更加簡便易行,節約生產成本由此看來在大豆迎茬和15 a連作地按1∶1種子混種,控制線蟲和產量結果均要明顯優于其它種植方式。
參考文獻:
[1] 劉維志.植物病原線蟲學 [M].北京:中國農業出版社,2000.
[2] 王金陵.大豆 [M].哈爾濱:黑龍江科學技術出版社,1982.
[3] Chen S Y,P M Porter,C D Reese,W C Stienstra.Crop sequence effects on soybean cyst nematode and soybean and corn yields [J].Crop Science,2001,41(6):1843-1849.
[4] Chen F J,S Y Chen. Mycofloras in cysts females and eggs of the soybean cyst nematode in Minnesota [J].Applied Soil Ecology,2002,19(1):35-50.
[5] 田中艷,周長軍,吳耀坤,等.抗線6號大豆品種的選育[J].作物研究,2007(3):373.
[6] Mundt C C.Performance of wheat cultivars and cultivar mixtures in the presence of Cephalosporium stripe [J].Crop Protection,2002,21(2):93-99.
[7] Mundt C C.Use of multiline cultivars and cultivar mixtures for disease management [J].Phytopathology,2002,40:381-410.
[8] Zhu Y Y,H R Chen,et al.Genetic Diversity and Disease Control in Rice [J].Nature,2000,406:718-722.
[9] 朱有勇.遺傳多樣性與作物病害持續控制 [M].北京:科學出版社,2007.
[10] 楊昌壽,孫茂林.對利用多樣化抗性防治小麥條銹病的作用的評價 [J].西南農業學報,1989,2(2):53-56.
[11] 朱有勇,陳海如,范靜華,等.利用水稻品種多樣性控制稻瘟病研究[J].中國農業科學,2003,36(5):521-527.
[12] 陳企村,朱有勇,李振岐,等.小麥品種混種對條銹病發生程度的影響 [J].西北農林科技大學學報,2008,36(5):119 - 123.
[13] Akanda S I,C C Mundt.Effects of two-component wheat cultivar mixtures on stripe ruet severity [J].Phytopathology,1996,86(4):347-353.
[14] 薛 津.2003年黑龍江省審定推廣的大豆新品種 [J].大豆科學,2003,22(2):157.
[15] 邵玉彬,胡興國,袁淑明,等.抗線4號大豆引育試驗與推廣 [J].內蒙古農業科技,2006(3):62-63.
[16] 崔文馥.1990年黑龍江省審定推廣的四個大豆品種 [J].大豆科學,1990,9(2):184.
[17] 司兆勝,許艷麗,李兆林,等.不同茬口種植的大豆品種根滲出物對大豆胞囊線蟲卵孵化的影響 [J].中國油料作物學報,2004,26(3):62 - 66.
[18] 許艷麗,陳伊里,司兆勝,等.不同茬口條件下的作物根滲出物對大豆胞囊線蟲(Heterodera glycines)卵孵化的影響 [J].植物病理學報,2004,34(6):481-486.
[19] 孫漫紅,劉杏忠.連作土壤中大豆胞囊線蟲種群數量減少的原因探討 [J].植物病理學報,30(4):353-356,363.
[20] 劉二明,朱有勇,肖放華,等.水稻品種多樣性混栽持續控制稻瘟病研究[J].中國農業科學,2003,36(2):164-168.
[21] Leung H,Y Y Zhu,R M Imelda,et al.Using genetic diversity to achieve sustainable rice disease management[J].Plant Disease,2003,87(10):1156-1169.
[22] Smithson J B,J M Lenne.Varietal mixtures:a viable strategy for sustainable productivity in subsistence agriculture [J].Annals of Applied Biology,1996,128(1):127-158.
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年13期)2021-11-26 11:50:54
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
