寄主植物對草地螟中腸解毒酶及保護性酶活性的影響
2012-02-28 07:47:44馮紅林李克斌曹雅忠
植物保護 2012年1期
關鍵詞:影響
尹 姣, 馮紅林, 李克斌, 曹雅忠
(中國農業科學院植物保護研究所,植物病蟲害生物學國家重點實驗室,北京 100193)
昆蟲與植物在長期協同進化過程中形成了各種針對性的適應機制。昆蟲會有目的地選擇性取食不同的寄主植物,通過消化和吸收植物食料中的營養物質,并分解排泄有毒物質,才能正常地生長發育及繁殖。由于不同寄主植物的營養成分及次生代謝物質不同,昆蟲取食后體內的生理生化反應亦有所不同,進而會對其生長發育產生明顯影響。因此,在長期的演化過程中,昆蟲在生理功能上產生了相應的適應機制,其中一系列解毒酶系的出現,是這種適應的主要方式之一[1]。
正常情況下,生物體細胞內自由基如單線態氧(1O2)、過氧化氫(H2O2)、氫氧自由基(HO·)等的產生與清除是在超氧化物歧化酶(SOD)、過氧化氫酶(CAT)和過氧化物酶(POD)3種酶協調一致,處于一種動態平衡狀態,使自由基維持在一個低水平,從而防止自由基毒害,一旦這種平衡受到破壞,就可能產生傷害作用[2]。因此,Fridovich把以上3種酶統稱為保護酶系統[3]。保護酶系統的形成是昆蟲在進化過程中逐步形成的抵抗外源刺激物和不良因素影響的有效途徑。
草地螟(Loxostege sticticalis L.)是在我國東北、華北和西北地區嚴重發生的多食性害蟲之一,可嚴重為害大豆、甜菜、苜蓿、向日葵等作物及牧草,對農牧業生產造成了很大威脅。草地螟具有廣泛的寄主范圍,同時對其寄主植物又有一定的選擇性,這在其生物學記載中已有相應研究報道[4-7],研究發現草地螟的寄主植物可分為適宜寄主植物和非適宜寄主植物,草地螟幼蟲在3齡以前對寄主植物的專化性要求較高,但老齡幼蟲卻存在明顯的轉主為害習性,取食非適宜寄主植物亦可發育、繁殖,表明老齡幼蟲體內對于非適宜寄主存在一定的適應機制[8-9]。目前有關不同寄主植物對昆蟲體內解毒酶的影響作用已經進行了較廣泛研究[10-17],但是草地螟對不同寄主植物的生理適應機制仍然缺乏系統研究,尤其老齡幼蟲取食寄主范圍擴大的內在機理尚不夠清晰。因此,作者利用5種不同寄主植物飼養草地螟4齡幼蟲,探討了草地螟老齡幼蟲中腸酶系對不同寄主植物的反應變化,為闡明草地螟老齡幼蟲對不同寄主植物生理適應機理奠定基礎,進而為解析草地螟與寄主植物之間的相互關系以及在長期進化過程中的適應性對策提供理論依據。
1 材料與方法
1.1 蟲源及飼養條件
蟲源來自內蒙古烏蘭浩特市郊采集的越冬代蛹,經室內飼養繁殖后作為試驗材料。羽化后的草地螟成蟲用5%的葡萄糖水飼養;卵孵化后幼蟲用藜(Chenopodium album L.,俗名灰菜)飼養至4齡。成蟲和幼蟲的飼養條件為(22±1)℃,L16∥D8,RH 70%~80%。
1.2 寄主植物
試驗所用植物材料分別選取適宜寄主藜、向日葵、大豆和非適宜寄主玉米、馬鈴薯5種植物的幼苗葉片,其中向日葵、大豆、玉米、馬鈴薯在北京郊區的中國農科院植保所試驗田種植,藜采自試驗田內和周邊的自生植株。試驗前用蒸餾水沖掉各試驗植物葉面的異物,晾干后飼喂試蟲。
1.3 寄主植物對草地螟幼蟲中腸解毒酶的影響
1.3.1 酶液的制備
每種材料加入1.5 mL預冷的相應的酶緩沖液,羧酸酯酶用0.04 mol/L、p H7.0的PBS提取;乙酰膽堿酯酶用0.1 mol/L、p H7.4的PBS提取。材料在冰浴中勻漿,勻漿12 000 g,10 min,取上清液作為酶液。
1.3.2 羧酸酯酶(Car E)活性測定
參考Van Asperen的方法[18],并加以改進。取3.6 mLα-乙酸萘酯(α-NA),0.1 mL酶液混勻后,置30℃水浴15 min后,加1 mL DBLS試劑(1% 固藍B與5% 十二烷基硫酸鈉以2∶5混合),反應15 min后,于600 n m波長測吸光值。
α-乙酸萘酯貯液配制方法:取5.58 mgα-乙酸萘酯,8.25 mg毒扁豆堿,1 mL丙酮溶于0.04 mol/L、p H7.0的PBS中,并在100 mL容量瓶中定容制成α-乙酸萘酯貯液。
1.3.3 谷胱甘肽S-轉移酶活力測定
參照Boot h方法[19]。反應混合液含0.1 mL酶液,2.9 mL 0.1 mol/L 的 磷 酸 緩 沖 液 (p H 8.0),0.7 mL 0.03 mol/L的谷胱甘肽液,50μL 0.1 mol/L的CDNB液,在25℃水浴中反應30 min,加熱使其失活,在350 n m比色。
1.3.4 乙酰膽堿酯酶(Ach E)活力測定
參照Ell man方法[20]。反應混合液含有50μL酶液,2.8 mL 0.1 mol/L的磷酸緩沖液(p H 8.0),50μL 0.075 mol/L的碘化乙酰膽堿,0.1 mL 0.01 mol/L的DTNB液,在25℃水浴中保溫15 min,加入0.1 mL 10-3mol/L毒扁豆堿。410 n m 處比色。用谷胱甘肽作標準曲線。
1.3.5 蛋白含量測定
參照 Bradfor d的方法[21],取0.1 mL 酶液與5 mL考馬斯亮藍G250混勻,測定各酶液中可溶性蛋白含量,以牛血清蛋白制作標準曲線。
1.4 寄主植物對草地螟幼蟲中腸保護酶的影響
草地螟幼蟲取食不同寄主植物后中腸超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、過氧化物酶(POD)活力測定按南京建成生物工程研究所生產的SOD、CAT和POD試劑盒說明進行。
1.5 數據分析
應用SAS軟件對試驗數據進行分析,用Duncan氏多重比較進行處理間的差異顯著性檢驗。
2 結果與分析
2.1 不同寄主植物對草地螟幼蟲中腸解毒酶的影響
2.1.1 寄主植物對草地螟中腸羧酸酯酶的影響
試驗結果表明,取食不同寄主植物會顯著影響草地螟幼蟲中腸羧酸酯酶活力(圖1)。其中,取食藜時中腸羧酸酯酶活力最高,顯著高于取食其他寄主植物時羧酸酯酶活力。同時,隨著取食時間的延長,羧酸酯酶活力明顯增加,其中,取食非適宜寄主植物(如馬鈴薯、玉米)比適宜寄主的羧酸酯酶(藜、大豆)活力增加迅速。盡管取食非適宜寄主植物能夠顯著刺激羧酸酯酶活力的增加,但是取食適宜寄主植物時中腸羧酸酯酶活力一直顯著高于取食非適宜寄主植物時的羧酸酯酶活力,說明取食適宜寄主植物會在短期內刺激羧酸酯酶的快速增加,隨著取食時間的延長,羧酸酯酶活力不再顯著變化;取食非適宜寄主植物則會在較長時間內刺激幼蟲中腸內羧酸酯酶活力的改變,可能是草地螟幼蟲對于非適宜寄主植物的一種適應機制。
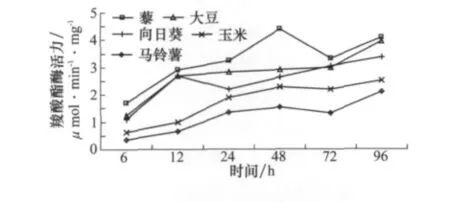
圖1 取食不同寄主植物后草地螟幼蟲體內羧酸酯酶活力的時序變化
2.1.2 寄主植物對草地螟中腸谷胱甘肽S-轉移酶的影響
取食不同寄主植物對草地螟中腸谷胱甘肽S-轉移酶活力會產生顯著影響(圖2)。在取食時間較短時,取食藜的幼蟲中腸酶活力顯著高于取食其他寄主植物的酶活力,但是,隨著取食時間的延長,取食藜的幼蟲體內谷胱甘肽S-轉移酶活力變化不大,但是取食另外4種寄主植物時幼蟲中腸的谷胱甘肽S-轉移酶活力具有非常顯著的變化,到24 h時取食5種不同寄主植物的中腸谷胱甘肽S-轉移酶活力已經不再具有顯著差異。
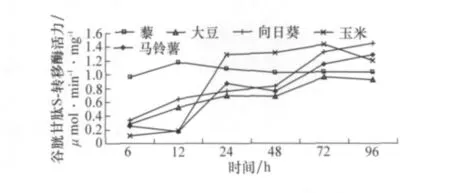
圖2 取食不同寄主植物后草地螟幼蟲體內谷胱甘肽S-轉移酶活力的時序變化
2.1.3 寄主植物對草地螟中腸乙酰膽堿酯酶的影響
取食不同寄主植物后草地螟幼蟲中腸的乙酰膽堿酯酶活力具有顯著差異(圖3)。在取食時間較短時(如6 h),草地螟幼蟲中腸乙酰膽堿酯酶活力無顯著差異。但是,隨著取食時間的延長,取食適宜寄主后中腸的乙酰膽堿酯酶活力增長顯著高于取食非適宜寄主植物后的中腸酶活力,取食96 h后,取食適宜寄主植物后的中腸乙酰膽堿酯酶活力比6 h時增加了6~8倍,而取食非適宜寄主植物時則酶活力變化不大,說明適宜寄主植物可以刺激幼蟲中腸內乙酰膽堿酯酶活力。
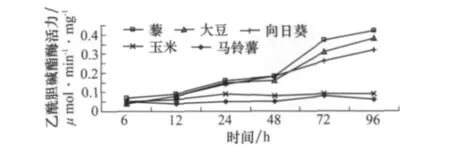
圖3 取食寄主植物不同時間后草地螟幼蟲體內乙酰膽堿酯酶活力的變化
由圖1~3可以看出,寄主植物對于幼蟲中腸的解毒酶影響顯著,取食適宜寄主植物時酶活力較高,但是隨著取食時間的延長,取食非適宜寄主植物的羧酸酯酶和谷胱甘肽S-轉移酶的酶活力也會逐漸增加,表明取食適宜寄主植物時可刺激草地螟幼蟲解毒酶活力立即升高,而非適宜寄主植物長時間的刺激可以引起部分解毒酶的快速增加。
2.2 不同寄主植物對草地螟幼蟲中腸保護酶的影響
2.2.1 寄主植物對草地螟中腸超氧化物歧化酶的影響
圖4顯示,取食初期(6、12、24 h),取食不同寄主植物的幼蟲中腸SOD活力差異顯著,其中取食藜、大豆和馬鈴薯時SOD活力較高。但是隨著取食時間的延長,酶活力不再具有顯著差異,表明不同寄主植物可以明顯刺激草地螟幼蟲中腸超氧化物歧化酶活力變化。
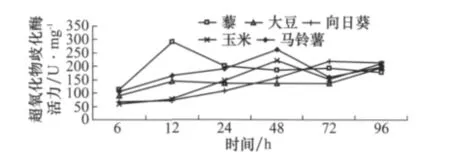
圖4 取食寄主植物不同時間后草地螟幼蟲體內超氧化物歧化酶活力的變化
2.2.2 寄主植物對草地螟中腸過氧化氫酶的影響
圖5顯示,取食不同寄主植物對草地螟中腸CAT活力有一定影響,其中取食馬鈴薯48 h時酶活力較高,但是隨著取食時間的延長,取食不同寄主植物的CAT活力逐漸趨于一致。
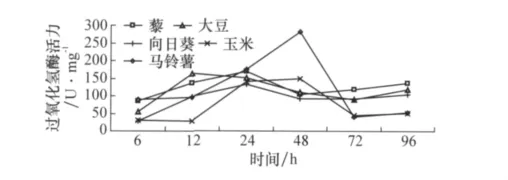
圖5 取食寄主植物不同時間后草地螟幼蟲體內過氧化氫酶活力的變化
2.2.3 寄主植物對草地螟中腸組織過氧化物酶的影響
取食不同寄主植物對草地螟中腸過氧化物酶產生一定的影響(圖6),除馬鈴薯之外,取食其他寄主植物隨取食時間的延長變化幅度不大。
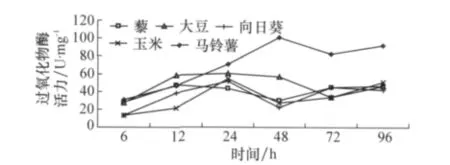
圖6 取食寄主植物不同時間后草地螟幼蟲體內組織過氧化物酶活力的變化
從圖4~6可以看出,盡管在取食初期不同寄主植物會對幼蟲中腸不同的保護酶產生影響,但是隨著取食時間的延長,取食不同寄主植物的草地螟其中腸保護酶活力趨于一致,相互間不再具有顯著差異。
3 討論
羧酸酯酶、谷胱甘肽-S-轉移酶和乙酰膽堿酯酶等是昆蟲體內重要的解毒酶,對分解外源毒物、維持正常生理代謝起重要作用。其中谷胱甘肽-S-轉移酶主要功能是催化某些內源性或外來有害物質的親電子基團與還原型谷胱甘肽的巰基偶聯,增加其疏水性使其易于穿越細胞膜,分解后排出體外,從而達到解毒的目的[22]。已有研究表明,虎鳳蝶(Papilio glaucus)、月尾大蠶蛾(Actias l una)、棉蚜(Aphis gossypii)、小菜蛾(Pl utell a xylostella)、繡線菊蚜(Aphis citricol a)、棉鈴蟲(Helicover pa ar miger a)、南美斑潛蠅(Liriomyza huidobrensis)、甜菜夜蛾(Spodoptera exigua)、豆莢野螟(Mar uca testulalis)、斜紋夜蛾(Spodoptera litur a)等昆蟲取食不同寄主植物時其體內各種解毒酶活性也不同[10-17,23-25]。 本 文 研 究 結 果 亦 表 明 草 地 螟 幼 蟲 取食不同寄主植物對中腸解毒酶羧酸酯酶、谷胱甘肽S-轉移酶和乙酰膽堿酯酶活性產生顯著影響。比較取食適宜寄主與非適宜寄主的幼蟲中腸解毒酶活性差異發現,其相互間酶活力不僅存在量的差異,而且存在質的差異,相對而言,較短時間內,取食適宜寄主植物時中腸解毒酶活力較高,但是隨著取食時間的延長,取食非適宜寄主的幼蟲中腸酶活力快速上升,96 h后酶活力可增加3~7倍,從而縮小取食不同寄主植物的幼蟲中腸解毒酶活力的差值。分析中腸解毒酶活力的這種適應性變化,推測可能是由于草地螟老齡幼蟲取食量暴增,僅靠適宜寄主無法滿足種群生存的需要,因而需要擴展取食范圍,不斷試探取食非適宜寄主,而非適宜寄主植物對中腸解毒酶系不斷產生刺激,在長期進化過程中誘導其對非適宜寄主產生了適應性,這種誘導是長期積累的結果,也需要一定時間的刺激才能獲得,所以草地螟幼蟲在取食非適宜寄主一定時間后酶活力會顯著升高。但是,由于不同寄主植物次生物質種類和含量不同,所以刺激中腸產生的解毒酶含量和種類也不盡相同,從而幼蟲對不同寄主植物的適應程度也各不相同,與原來的試驗結果相符[8]。另外,非適宜寄主植物對中腸乙酰膽堿酯酶活力改變不大,表明寄主植物僅對部分中腸解毒酶具有明顯的刺激作用。總之,這是昆蟲在進化過程中對食物代謝途徑的適應性調整,另外可滿足解毒和節省能源兩種需要。
目前已有試驗表明實驗昆蟲體內存在的過氧化氫酶(CAT)和過氧化物酶(POD)的活性和昆蟲對外界刺激物相應強度的反應有關[26-30]。超氧物歧化酶(SOD)雖然是生物細胞內最重要的清除自由基的酶,但其清除O2而成H2O2后,H2O2與O2形成毒性更強的HO·必須由過氧化氫酶和過氧化物酶來分解H2O2,所以CAT和POD兩種酶在生命系統的保護作用上顯得更為重要。本文研究結果表明草地螟幼蟲取食不同寄主植物后對中腸保護酶SOD、CAT和POD活力均有一定影響,但是隨著取食時間的延長,不同處理之間酶活力不再具有顯著差異,推測可能是取食初期,草地螟幼蟲對于不同寄主植物的刺激產生了較強的反應,所以酶活力變化較大,但是隨著取食時間的延長,幼蟲逐步適應了不同的寄主植物從而酶活力也趨于較平穩的狀態。草地螟幼蟲對于不同寄主植物的這種適應性是其種群發展和延續的保證。
[1] 朱麟,古德祥.昆蟲對植物次生性物質的適應策略[J].生態學雜志,2000,19(3):36-45.
[2] 李周直,沈惠娟,蔣巧根,等.幾種昆蟲體內保護酶活力的研究[J].昆蟲學報,1994,37(4):399-403.
[3] Fridovich I.Oxygen is toxic[J].Bioscience,1977,27(7):462.
[4] 顧成玉,梁艷春,張廣芝.草地螟發生為害特點與防治策略的探討[J].病蟲測報,1987(S1):32-34.
[5] 劉光濤,郭向東,張金,等.草地螟的生物學特性及其發生與環境條件的關系[J].病蟲測報,1987(S1):59-64.
[6] 康愛國,樊榮賢,張玉慧,等,草地螟卵和越冬幼蟲發生分布調查[J].植保技術與推廣,1999,19(2):3-4.
[7] 陳玉寶.草地螟產卵習性觀察[J].內蒙古農業科技,1999(3):27-30.
[8] 尹姣,曹雅忠,羅禮智,等.寄主植物對草地螟種群增長的影響[J].植物保護學報,2004,31(2):173-178.
[9] 尹姣,曹雅忠,羅禮智,等.草地螟對寄主植物的選擇性及其化學生態機制研究[J].生態學報,2005,25(8):1844-1852.
[10]高希武.寄主植物對棉蚜羧酸酯酶活性的影響[J].昆蟲學報,1992,35(6):267-272.
[11]李云壽,羅萬春,慕立義,等.不同寄主植物對小菜蛾艾氏劑環氧化酶和乙酰膽堿酯酶活性的影響[J].植物保護學報,1996,23(2):181-184.
[12]王建軍,戴志一,楊益眾.寄主植物對棉鈴蟲體內解毒酶活性的影響[J].江蘇農業研究,2000,21(2):58-61.
[13]陳洪國,袁天文.寄主植物對斜紋夜蛾藥劑敏感性及中腸羧酸酯酶等的影響[J].湖北農業科學,2003(6):65-67.
[14]魏秋學,何玉仙,楊秀娟,等.寄主植物對南美斑潛蠅羧酸酯酶及乙酰膽堿酯酶活力的影響[J].江西農業大學學報,2004,26(3):345-347.
[15]李子玲,韋綏概,韋飚,等.寄主植物對甜菜夜蛾的發育和繁殖及 體 內 酯 酶 活 性 的 影 響 [J].昆 蟲 知 識,2005,42(3):284-289.
[16]潘亞飛,孟建玉,張小亞,等.寄主對豆野螟的藥劑敏感性和體內解毒酶活性的影響[J].昆蟲知識,2006,43(4):496-500.
[17]呂朝軍,韓巨才,劉慧平,等.寄主植物對蘋果黃蚜藥劑敏感性及解毒酶活性的影響[J].植物保護學報,2007,34(5):534-538.
[18]Asperen K Van.A study of housefly esterase by means of a sensitive colori metric met hod[J].J Ins Physiol,1962(8):401-416.
[19]Booth J,Boyland E,Sins P.An enzy me from the rat liver catalyzing conjugation with glutat hione[J].J Biochem,1961,79:516-524.
[20]Ell man G,Courtney K,Andres V,et al.A new and rapid colori metric deter mination of acetylcholinesterase activity[J].Biochem Phar macol,1961,7:88-92.
[21]Bradfor d M.A rapid and sensitive met hod for the quantitation of microgra m quantities of protein utilizing the principles of protein-dye binding[J].Analyt Biochem,1976,72:248-254.
[22]楊海靈,聶力嘉,朱圣庚,等.谷胱甘肽硫轉移酶結構與功能研究進展[J].成都大學學報,2006,25(1):19-24.
[23]Lindr ot h R L.Host plant alteration of detoxification enzy me in Papilio gl aucus glaucus[J].Ento mol Exp Appl,1989,50:29-35.
[24]Lindrot h R L.Chemical ecology of the luna mot h:effects of host planton detixification enzy me activity[J].J Chem Ecol,1989,15:2019-2029.
[25]高希武,鄭炳宗,陳仲兵.小菜蛾羧酸酯酶性質的研究[J].南京農業大學學報,1996,19(S1):122-126.
[26]戈峰,李典謨,邱業先,等.松樹受害后一些化學物質含量的變化及其對馬尾松毛蟲種群參數的影響[J].昆蟲學報,1997,40(4):338-342.
[27]Dowd P F,Lagri mini P F.Exa mination of different tobacco(Nicotiana spp.)types under and over producing tobacco anionic per oxidase for their leaf resistance to Helicover pa zea[J].Chem Ecol,1997,23(10):2357-2370.
[28]Moran,Patrick J.Cross-effects of infection of cucu mber with f ungal and bacterial pat hogens,and the r ole of per oxidase[J].Bull Ecol Soc Amer,1997,78(4):287.
[29]吳小鋒,徐俊良,崔為正.家蠶血液過氧化氫酶活力及其與蠶體抗逆性的關系[J].昆蟲學報,1998,41(2):124-128.
[30]張瑛,嚴福順.蟲害誘導的植物揮發性次生物質及其在植物防御中的作用[J].昆蟲學報,1998,41(2):210.
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
