3種金雞菊的光合特性比較
2012-05-29 13:15:50郭運雪蔣文偉黃建榮
浙江農林大學學報 2012年5期
關鍵詞:植物
郭運雪,蔣文偉,戴 鋒,黃建榮
(1.浙江農林大學 風景園林與建筑學院,浙江 臨安 311300;2.上房園藝有限公司,上海 201114)
金雞菊屬Coreopsis為菊科Compositae多年生宿根草本花卉,原產美洲,是一種觀賞植物。可觀葉,也可觀花。莖直立,多分枝,喜陽光直射,性強健,耐寒,耐干旱瘠薄。春夏之間,花大色艷,常開不絕,適合布置花壇或作花境及背景材料,叢植于山前、籬旁、林中,應用較廣泛,亦可作切花。其種類豐富,管理粗放,適應性強,還能自行繁衍,是極好的疏林地被和屋頂綠化材料,應用非常普遍。因其既耐干旱瘠薄又管理粗放,花后修剪可秋季二次開花,提高了觀賞質量,而且大大節約了養護成本。另外,金雞菊在中國長江流域以南表現常綠狀態,這樣冬季有景可觀,彌補了宿根草本冬季落葉后景觀較差的缺點。由于具有觀賞性及其生長的抗逆性,奧運會、世博會都有金雞菊靚麗的身影,也被作為公路或城市中的其他地段水土保持的植被[1-3]。目前,對金雞菊的研究主要集中在栽培技術[4-5],提取物降壓降脂的作用[6-7],黃酮類化合物的提取和乙醇提取物對菌類的抑制作用[8-9],植株水浸液的化感作用[10]等方面,對金雞菊的光合生理特性等生態適應性方面的研究少見報道。一定程度上,光合特征參數的變化能夠反映植物對環境因子的適應性[11-12]。在自然狀態下,結合環境因子測定該物種的光合作用更能直接反映該植物的光合生理過程[13]。在植物的主要生理生態指標中,光強響應曲線的測定可以明確植物的光飽和點和光補償點,進而確定植物的光照類型,以便正確地在園林中應用[14]。本研究對金雞菊的光合特性進行測定,旨在了解它們的光合生理生態特性,為進一步引種、馴化和栽培提供理論依據。
1 材料與方法
1.1 供試材料
試驗材料為3種金雞菊的扦插苗:大花金雞菊Coreopsis grandiflora,重瓣金雞菊Coreopsis lanceolata,天堂之門金雞菊Coreopsis rosea,由上海上房園藝有限公司提供。將扦插苗栽植于直徑為18 cm的花盆中,栽培土按園土(m)∶泥炭(m)∶珍珠巖(m) =3∶1∶1 的比例配比,常規管理。
1.2 試驗方法
1.2.1 光合作用的光響應曲線的測定 選擇晴天9∶00-11∶00,用Li-6400光合測定儀通過控制光照強度測定光響應曲線。樣本室內氣流速率控制在500 μmol·s-1,溫度控制在當時的外界環境溫度。在控制條件下,利用Li-6400型光合作用系統和紅藍光源進行設置,設定光照強度在0~2000 μmol·m-2·s-1范圍內(由高到低),梯度為 2000,1500,1000,600,300,200,100,80,50,30,20,0 μmol·m-2·s-1,測定相應的凈光合速率(Pn)值,每一個光強下有一定的適應時間,待光合測定儀上的數據穩定后再記錄,每組梯度重復3次。凈光合速率為0時的光強為光補償點(PLC),凈光合速率最大時的光強為光飽和點(PLS)。采用光合助手軟件Photosyn Assistant 1.1.2得出光補償點、光飽和點、最大凈光合速率(Pnmax)以及表觀量子效率(EAQ)。
1.2.2 凈光合速率日變化的測定 采用美國產的Li-6400便攜式光合作用測定系統,葉室選用6400-15葉室,選擇晴朗無風的天氣,在7∶00-19∶00隔2 h測定1次,3個重復,重復記錄5個觀測值,取其平均值作為該時刻的測定值。選擇生長健康的植株,隨機選取該植物向陽面的葉片進行測定。待系統穩定后,取葉片瞬時凈光合速率值,同時讀取蒸騰速率(Tr),氣孔導度(Cond),胞間二氧化碳摩爾分數(Ci),光合有效輻射(PAR),氣溫(Ta),相對濕度(Rh)等生理生態指標。水分利用效率的計算公式為:水分利用效率(EWUE)=Pn/Tr。
1.3 數據處理
使用Microsoft Excel和SPSS 17.0對測得的數據進行處理和分析。
2 結果與分析
2.1 3種金雞菊的光響應差異
統計結果表明:3種金雞菊的光響應曲線變化有顯著性的差異。從圖1可以看出:在0~100 μmol·m-2·s-1的光強范圍內,3種金雞菊的凈光合速率幾乎呈線性增長,在 100~200 μmol·m-2·s-1,隨著光照強度的增加,凈光合速率的增長速度逐漸減慢,當光強達到1500 μmol·m-2·s-1以上時,金雞菊的凈光合速率的增長處于極緩慢狀態,基本維持穩定。說明植物在一定的光照強度下,葉片的光合作用處于飽和狀態,當光照強度達到光飽和點以后,凈光合速率甚至有下降趨勢。
比較3種金雞菊之間的光響應曲線,發現3種金雞菊葉片的凈光合速率對光合有效輻射的響應趨勢基本一致,均是在低光照下快速上升后逐漸穩定。在0~100 μmol·m-2·s-1時,3種金雞菊凈光合速率相差不大。在光合有效輻射增大到100 μmol·m-2·s-1后,3種金雞菊間的凈光合速率值差異明顯變大,重瓣金雞菊最大,大花金雞菊次之,天堂之門金雞菊最低。總體上,金雞菊的光響應與美國紫菀Aster novae-angliae[15]等草本植物先迅速上升后趨于穩定的光響應規律基本一致。表1的數據說明不同種金雞菊的光補償點和光飽和點具有一定的差異。從表1中可以看出:大花金雞菊的光補償點和光飽和點最低,能有效地利用弱光進行光合作用;3種金雞菊中,重瓣金雞菊具有相對較低的光補償點,又具有最高的光飽和點和最大的凈光合速率,說明它對光的適用范圍相對最大,對強光的適應能力最強,光合適應能力也最強;天堂之門金雞菊的光補償點最高,而光飽和點相對較低,說明其光適應范圍較窄,利用強光的能力低于重瓣金雞菊。

表1 3種金雞菊的光合參數Table1 Photosynthetic parameters of three species of Coreopsis
2.2 3種金雞菊的凈光合速率的日變化比較

圖1 3種金雞菊的光響應曲線Figure1 Light-response curve of photosynthesis of three species of Coreopsis

圖2 3種金雞菊凈光合速率的日變化Figure2 Diurnal variation of Pnof three species of Coreopsis
植物光合作用隨著環境條件的變化而有所不同,一般條件下栽培植物的光合作用日變化呈單峰型或雙峰型曲線[16]。由圖2可知:3種金雞菊的凈光合速率日變化表現出相似的規律,均為雙峰型曲線。在上午9∶00,大花金雞菊和天堂之門金雞菊出現第1個峰值,分別為大花金雞菊 (7.53±0.05)μmol·m-2·s-1,天堂之門金雞菊為(6.91 ± 0.10)μmol·m-2·s-1,重瓣金雞菊稍晚,在 11∶00達到第 1個高峰值,為(10.46 ±0.18)μmol·m-2·s-1;之后隨著光合有效輻射和氣溫繼續升高,Rh不斷降低,凈光合速率開始下降,13∶00時跌入低谷,出現光合“午休”現象。
午后隨著光合有效輻射的降低,凈光合速率又逐漸升高,到15∶00出現另一個高峰值,由大至小依次為重瓣金雞菊[(7.86 ± 0.10)μmol·m-2·s-1]>大花金雞菊[(4.77 ± 0.06)μmol·m-2·s-1]>天堂之門金雞菊[(4.23±0.13) μmol·m-2·s-1],之后都迅速下降,至 19∶00 降至最低。方差分析表明,3 種金雞菊間的第 1峰值和第2峰值差異均顯著(P<0.05),每種金雞菊的第1峰值與其第2峰值差異也顯著(P<0.05)。在光合作用日進程中,3種金雞菊的第1峰值均高于其第2峰值。
2.3 環境因子日變化特征
植物的光合速率、蒸騰速率和水分利用效率等生理過程受多種環境因子的影響[17]。圖3結果顯示,光合有效輻射瞬時變化較大,對植物的光合作用有顯著的影響,為先升后降的單峰曲線,13∶00時達到最大值。氣溫呈現先升后降的變化趨勢,與光合有效輻射變化相似,而相對濕度則呈現先降后升的變化趨勢,與光合有效輻射的變化相反。
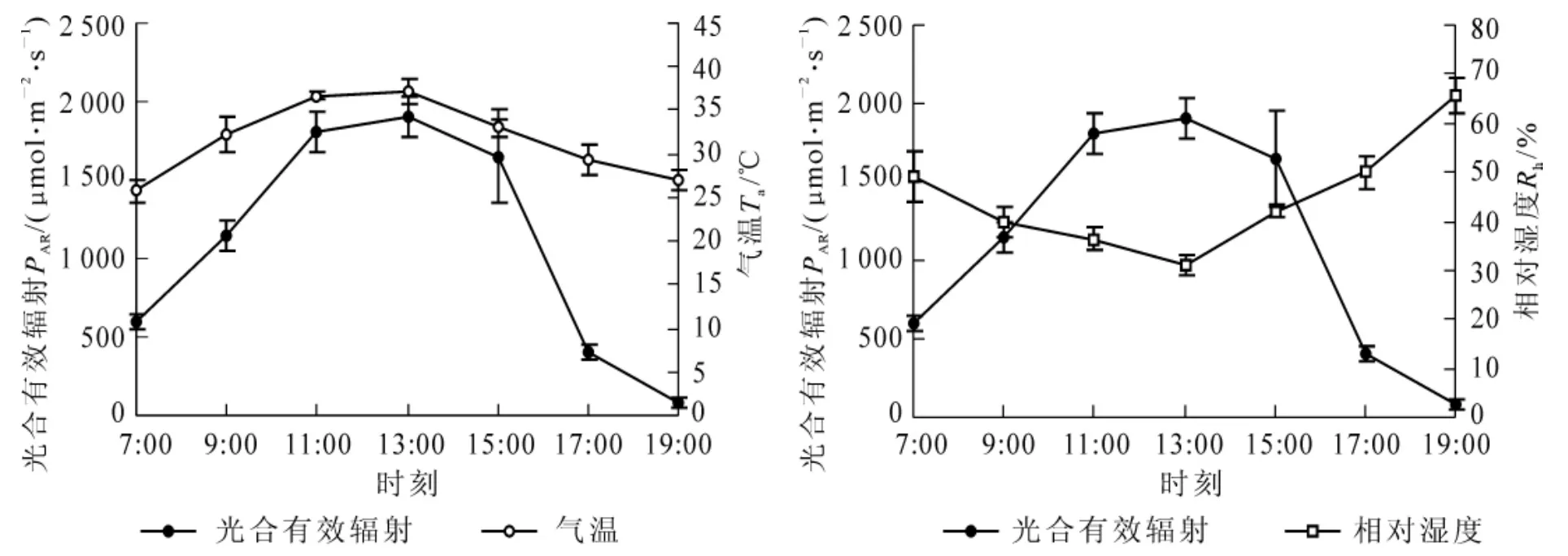
圖3 光合有效輻射、氣溫和相對濕度日變化Figure3 Diurnal changes of photosynthetic active radiation,air temperature and relative humidity
2.4 3種金雞菊的光合參數日變化
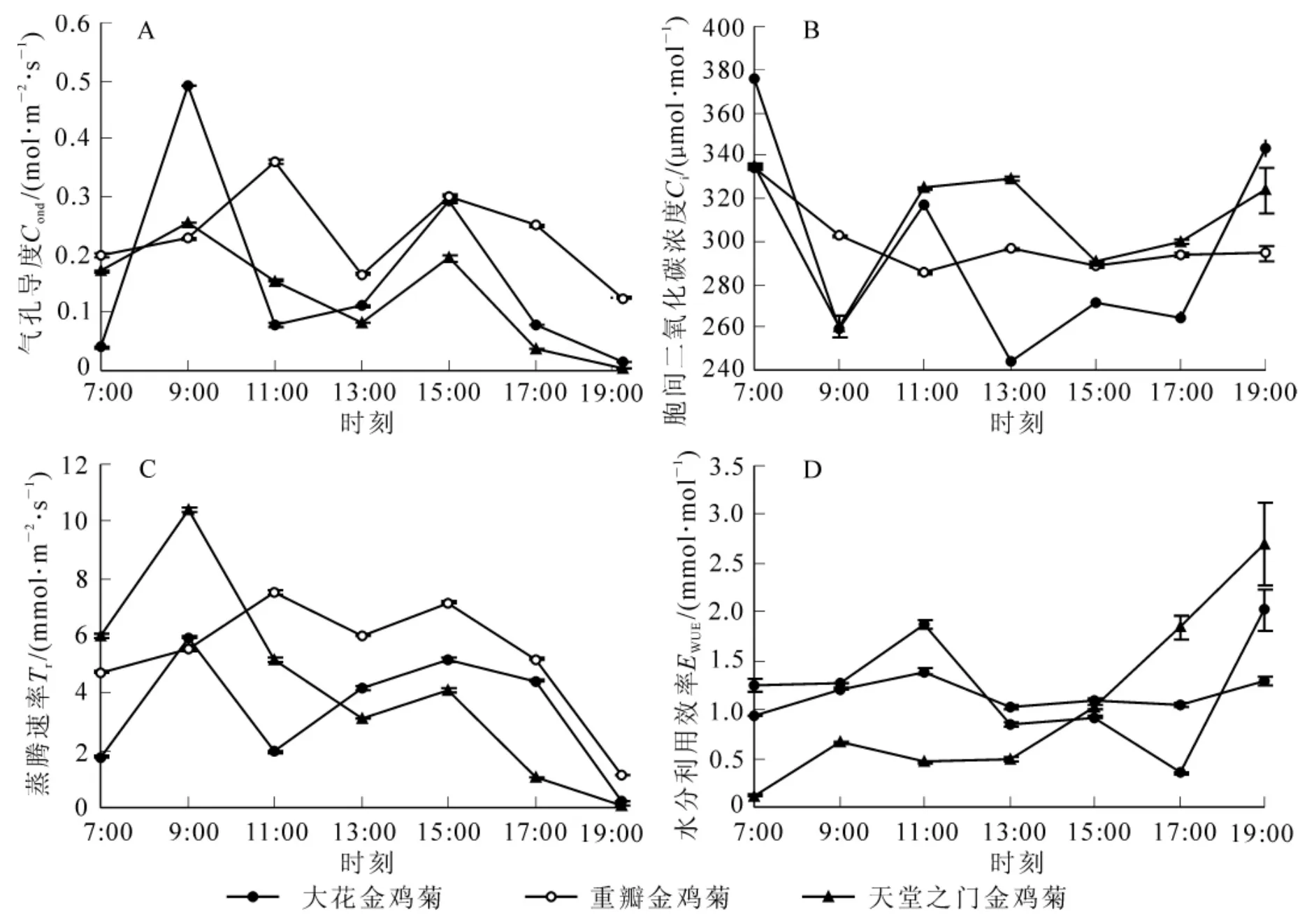
圖4 3種金雞菊的光合特征參數日變化曲線Figure4 Diurnal changes of photosynthetic character parameters of three species of Coreopsis
影響植物光合作用的生理生態因子主要包括:光合有效輻射、葉片氣孔導度、胞間二氧化碳摩爾分數、蒸騰速率等。這些因子共同作用影響植物的光合速率變化。3種金雞菊的氣孔導度日變化均呈雙峰型(圖4),與凈光合速率日變化趨勢相似,大花金雞菊和天堂之門金雞菊的第1次峰值均出現在9∶00,峰值分別為 0.49 μmol·m-2·s-1和 0.26 μmol·m-2·s-1,重瓣金雞菊的第 1 次峰值出現在 11∶00,為 0.36 mol·m-2·s-1,3 種金雞菊的第 2 次峰值均出現在 15∶00,由大至小依次為重瓣金雞菊(0.30 μmol·m-2·s-1)>大花金雞菊 (0.29 μmol·m-2·s-1)>天堂之門金雞菊(0.19 μmol·m-2·s-1)。氣孔導度在 9∶00 以后迅速下降至13∶00達到最低,而這期間正是光合有效輻射迅速增強,氣溫升高,大氣相對濕度降低的時候,植物能針對外界環境條件變化與植株自身狀況,通過調節氣孔的開閉程度,在保持植株正常生理活動的前提下,達到最大二氧化碳固定量與最小水分散失量。這也說明金雞菊對氣孔的調節能力是其能夠耐干旱的生理適應機制之一[18]。3種金雞菊的胞間二氧化碳摩爾分數與氣孔導度呈相反的變化趨勢,基本隨著氣孔導度的升高而逐漸降低,在氣孔導度出現第1峰值的時刻,3種金雞菊的胞間二氧化碳摩爾分數分別出現低谷,谷值大小分別為重瓣金雞菊[(285.3±0.48)μmol·mol-1]>天堂之門金雞菊[(260.57±5.09)μmol·mol-1]>大花金雞菊[(259.38 ± 4.27) μmol·mol-1],說明隨著上午金雞菊葉片光合作用的增強,消耗更多的二氧化碳原料,使得胞間二氧化碳摩爾分數降低。3種金雞菊的蒸騰速率也均呈現雙峰曲線,出現蒸騰 “午休”現象,2次出現峰值時刻與其凈光合速率達到峰值時刻相一致,但峰值大小有所變化,說明3種金雞菊的水分利用效率有差異。由圖4D可知:3種金雞菊的水分利用效率在上午都有1個峰值,在傍晚時都呈現上升的趨勢。大花金雞菊在11∶00達到最大,為(1.87±0.05)mmol·mol-1,17∶00達到最低,隨后急劇上升;重瓣金雞菊水分利用效率日變化不大,在11∶00達到峰值 (1.39±0.03)mmol·mol-1之后有稍微下降,然后呈現緩慢上升趨勢;天堂之門金雞菊在9∶00時即達到1個峰值為(0.66±0.01)mmol·mol-1,至13∶00較平穩,之后呈上升趨勢。3種金雞菊的日平均水分利用效率大小依次為大花金雞菊[(1.22 ± 0.015) mmol·mol-1]>重瓣金雞菊[(1.14 ± 0.008) mmol·mol-1]>天堂之門金雞菊[(1.03 ±0.004)mmol·mol-1]。方差分析表明:大花金雞菊和天堂之門金雞菊之間的日平均水分利用效率差異顯著(P<0.05),與重瓣金雞菊差異顯著(P<0.05),重瓣金雞菊與天堂之門金雞菊之間差異也顯著(P<0.05)。對于傍晚水分利用效率上升的現象,李小俊等[17]認為在午間,植物的大部分水分被用于蒸騰作用,從而相對減少了用于光合作用的水分,傍晚由于根部及時補水應用于植物葉片的光合作用,所以水分利用效率有所提高,據此可以更好地進行金雞菊的水分管理。
3 討論與結論
光合能力強弱在一定程度上取決于物種的遺傳特性[19],光飽和點和光補償點的變化具有一定的物種遺傳穩定性[20]。3種金雞菊的光補償點大小排序依次為天堂之門金雞菊>重瓣金雞菊>大花金雞菊;光飽和點大小順序為重瓣金雞菊>天堂之門金雞菊>大花金雞菊;最大凈光合速率排序為重瓣金雞菊>大花金雞菊>天堂之門金雞菊。一般情況下,光補償點和光飽和點均較低屬于耐陰植物,能充分利用弱光進行光合作用,反之屬于陽性植物。光補償點較高、光飽和點較低的植物對光照的適應性較弱,而光補償點較低、光飽和點較高的植物對光環境的適應性較強[21]。由此得出:重瓣金雞菊光適應范圍最廣,天堂之門金雞菊光適應范圍較窄。綜合比較3種金雞菊對光的適應范圍大小為重瓣金雞菊>大花金雞菊>天堂之門金雞菊,對強光的利用能力依次為重瓣金雞菊>天堂之門金雞菊>大花金雞菊。
通過分析3種金雞菊的光合日進程,結果發現:凈光合速率均表現為雙峰型,最大凈光合速率均出現在上午和下午,說明3種金雞菊在夏季出現明顯的光合“午休”現象。一般認為:午間強光引起凈光合速率下降,產生光抑制[18],導致植物的光合“午休”現象。Berry等[22]研究認為,導致產生光合“午休”現象的主要原因可分為氣孔因素和非氣孔因素,如果氣孔導度減小而光合作用增強,胞間二氧化碳摩爾分數降低時,可表明氣孔因素是使植物凈光合速率減少,產生光合“午休”的主要原因;如果相反,當光合速率降低伴隨著胞間二氧化碳摩爾分數升高時,則表明影響植物光合“午休”的原因主要是非氣孔因素[23]。研究結果表明:大花金雞菊、重瓣金雞菊、天堂之門金雞菊的光合“午休”主要是受非氣孔限制因素即細胞光合性能下降的影響,跟Farquhar等[24]的研究結果相似。
凈光合速率的變化是由植物生理生態因子共同作用的結果,而生理生態因子則受到環境因子的影響。在光合有效輻射、相對濕度、氣溫等環境因子相同的條件下,3種金雞菊的凈光合速率與葉片氣孔導度、胞間二氧化碳摩爾分數、蒸騰速率等生理因子各不相同,說明植物的生長受多個環境因子的綜合影響,各環境因子間相互制約,不同時期起主導作用的環境因子對凈光合速率的影響有所差異,同一環境因子對不同植物的凈光合速率、胞間二氧化碳摩爾分數和葉片氣孔導度的影響也不同[18]。
水分利用效率反映了植物對水分的利用效率,也在一定程度上也反映了植物的耗水性和抗旱性[17]。僅根據水分利用效率就可以得出3種金雞菊抗旱能力大小依次為大花金雞菊>重瓣金雞菊>天堂之門金雞菊。由于水資源的日益緊缺與城市環境對于園林綠化發展用水需求之間的矛盾,在全國大部分城市,尤其是北方城市尤為突出,已經成為長時期以來城市園林綠化發展的“瓶頸”之一。如何調整城市園林綠化的品種結構入手,大力推廣節水型園林植物來綜合解決城市水資源緊缺與綠化發展用水需求這一矛盾,已成為擺在全國各地園林綠化建設工作者面前的緊要課題。大花金雞菊和重瓣金雞菊就是在中國北方城市推廣節水型園林植物的一個很好的選擇。
[1]秦賀蘭.北京奧運用花品種系列介紹之十一大花金雞菊生產技術[J].中國花卉園藝,2008(8):28.QIN Helan.Beijing Olympic Games with the introduction of flower varieties series(Ⅺ)production technology of Coreopsis grandiflora [J].China Flowers&Hortic,2008 (8):28.
[2]孫磊,黃建榮.新優地被“靚”世博[J].園林,2009 (8):62.SUN Lei,HUANG Jianrong.New excellent ground cover plants on the Ecpo [J].Garden,2009 (8):62.
[3]梁玉.外來種大花金雞菊入侵的影響因子及其遺傳多樣性研究[D].濟南:山東大學,2007.LIANG YU.The Study on Factors Influencing Invasions and Genetic Diversity in Alien Species Coreopsis grandiflora Hogg [D].Ji’nan:Shandong University,2007.
[4]任真,趙洪波.大花金雞菊栽培[J].新農業,2008 (5):52 - 53.REN Zhen,ZHAO Hongbo.Cultivation of Coreopsis grandiflora [J].New Agric,2008 (5):52 - 53.
[5]常爽,閆瑞鳳,史根花.金雞菊花草栽培管理技術[J].北京農業,2008(36):42-43.CHANG Shuang,YAN Ruifeng,SHI Genhua.Cultivation and management techniques of Coreopsis basalis [J].Beijing Agric,2008 (36):42 - 43.
[6]梁淑紅,哈木拉提,龐市賓,等.金雞菊提取物降血壓化學成分實驗研究[J].時珍國醫國藥,2010,21(7):1619-1621.LIANG Shuhong,Hamulati,PANG Shibin,et al.Experimental study on hypertensive effects of extracts of Coreopsis basalis [J].Lishizhen Med Mat Med Res,2010,21 (7):1619 - 1621.
[7]梁淑紅,龐市賓,劉曉燕,等.金雞菊提取物降血脂作用的研究[J].中國實驗方劑學雜志,2010,16(8):234-235.LIANG Shuhong,PANG Shibin,LIU Xiaoyan,et al.Study on lipid-lowering effects of extracts of Coreopsis basalis[J].Chin J Exp Trad Med Form,2010,16 (8):234 - 235.
[8]朱篤,陳飛彪,夏劍輝,等.金雞菊總黃酮的提取及含量測定[J].食品科學,2005,26(9):314-316.ZHU Du,CHEN Feibiao,XIA Jianhui,et al.Extraction and determination of flavonoids in flowers of Coreopsis basalis [J].Food Sci,2005,26 (9):314 - 316.
[9]宗磊,李秀嵐,程永芳,等.大花金雞菊乙醇提取物對植物病原真菌的抑制活性測定[J].安徽農業大學學報,2010,37 (2):200 - 203.ZONG Lei,LI Xiulan,CHENG Yongfang,et al.Antifungal activity of ethanol extract from Coreopsis grandiflora[J].J Anhui Agric Univ,2010,37 (2):200 - 203.
[10]杜明利,高巖,張汝民,等.大花金雞菊水浸液對6種常見園林植物種子萌發的化感作用[J].浙江農林大學學報,2011,28 (1):109- 114.DU Mingli,GAO Yan,ZHANG Rumin,et al.Allelopathic effects from aqueous extracts of exotic Coreopsis grandiflora on seed germination of six native ornamental plant species [J].J Zhejiang A & F Univ,2011,28 (1):109 - 114.
[11]尤鑫,龔吉蕊,葛之葳,等.2種雜交楊葉綠素熒光特性及光能利用[J].植物生態學報,2009,33(6):1148-1155.YOU Xin,GONG Jirui,GE Zhiwei,et al.Light energy utilization and chlorophyll fluorescence in two crossbreed poplars [J].Chin J Plant Ecol,2009,33 (6):1148 - 1155.
[12]王凱,朱教君,于立忠,等.遮陰對黃波羅幼苗的光合特性及光能利用效率的影響[J].植物生態學報,2009,33(5):1003 - 1012.WANG Kai,ZHU Jiaojun,YU Lizhong,et al.Effects of shading on the photosynthetic characteristics and light use efficiency of Phellodendron amurense seed [J].Chin J Plant Ecol,2009,33 (5):1003 - 1012.
[13]林夏珍,盧婷.遮光對窄頭橐吾形態及光合特性的影響[J].浙江林學院學報,2008,25(5):614-618.LIN Xiazhen,LU Ting.Morphological and photosynthetic characteristics of Ligularia stenocephala with shading [J].J Zhejiang For Coll,2008,25 (5):614 - 618.
[14]張真珍.紅花檵木的光合生理特性及其在園林中的應用研究[J].青海農林科技,2011(2):17-19.ZHANG Zhenzhen.Photosynthetic character of Loropetalum chinense and its application in the landscape and architecture [J].Sci Technol Qinghai Agric For,2011 (2):17 - 19.
[15]蔣文偉,陳婭瓊,黃建榮,等.4個美國紫菀品種光合生理特征比較[J].浙江林學院學報,2010,27(6):865-871.JIANG Wenwei,CHEN Yaqiong,HUANG Jianrong,et al.Photosynthetic characteristics with four cultivars of Aster novae-angliae [J].J Zhejiang For Coll,2010,27 (6):865 - 871.
[16]吳澤民,黃成林,韋朝領.黃山松群落林隙光能效應與黃山松的更新[J].應用生態學報,2000,11(1):13-18.WU Zemin,HUANG Chenglin,WEI Chaoling.Light effect of gaps in Huangshan pine community and regeneration of Huangshan pine [J].Chin J Appl Ecol,2000,11 (1):13 - 18.
[17]李小俊,張明如,張利陽,等.太行山低山丘陵區5種木本植物光合特性的比較[J].浙江農林大學學報,2011,28(2):180 - 186.LI Xiaojun,ZHANG Mingru,ZHANG Liyang,et al.Photosynthesis,light-use efficiency and water-use efficiency for woody species in the hilly area of the Taihang Mountains [J].J Zhejiang A & F Univ,2011,28 (2):180 - 186.
[18]林保花,劉金祥,肖生鴻,等.粵西鄉土香根草光合生理生態特征日動態分析[J].應用生態學報,2006,17(11):2041 - 2045.LIN Baohua,LIU Jinxiang,XIAO Shenghong,et al.Diurnal change of photosynthetic characteristics of native Vetiveria zizanioides in Western Guangdong [J].Chin J Appl Ecol,2006,17 (11):2041 - 2045.
[19]何文興,易津,李洪梅.根莖禾草乳熟期凈光合速率日變化的比較研究[J].應用生態學報,2004,15(2):205-209.HE Wenxing,YI Jin,LI Hongmei.Comparative study on daily change of photosynthesis rate of the rhi zomatous grasses in milky ripe stage [J].Chine J Appl Ecol,2004,15 (2):205 - 209.
[20]BJORKMAN O,HOLMGREN P.Adaptability of the photosynthetic apparatus to light intensity in ecotypes from exposed and shaded habitat[J].Physiol Plant,1963,16 (4):889 - 914.
[21]金則新,柯世省.云錦杜鵑葉片光合作用日變化特征[J].植物研究,2004,24(4):447-452.JIN Zexin,KE Shisheng.The diurnal variation of photosynthesis in leaves of Rhododendron fortunei[J].Bull Bot Res,2004,24 (4):447 - 452.
[22]BERRY J A,DOWNTON W J S.Environmental Regulation of Photosynthesis [M].New York:Academic Press,1982:263-345.
[23]許大全.氣孔的不均勻關閉與光合作用的非氣孔限制[J].植物生理學通訊,1995,31(4):246-252.XU Daquan.Non-uniform stomatal closure and non-stomatal limitation of photosynthesis [J].Plant Physiol Commun,1995,31 (4):246 - 252.
[24]FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis [J].Ann Rev Plant Physiol,1982,33:317- 345.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
