耐有機溶劑脂肪酶產生菌的篩選及其粗酶酶學性質
2012-06-01 09:08:29李俊峰李紅芳段效輝門晉名
食品科學 2012年3期
李俊峰,李紅芳,段效輝,門晉名,宿 烽
(1. 青島科技大學化工學院,山東 青島 266042;2. 煙臺出入境檢驗檢疫局,山東 煙臺 264000)
耐有機溶劑脂肪酶產生菌的篩選及其粗酶酶學性質
李俊峰1,李紅芳1,段效輝2,門晉名1,宿 烽1
(1. 青島科技大學化工學院,山東 青島 266042;2. 煙臺出入境檢驗檢疫局,山東 煙臺 264000)
以體積分數2%苯為篩選壓力,利用羅丹明B平板顯色法和搖瓶發酵法,從采集的花生地土壤樣品中分離篩選得到1株中度耐熱、耐堿脂肪酶產生菌,編號為H2。通過形態觀察、生理生化特性實驗及其16S rDNA基因序列對菌種進行鑒定。結果表明,H2菌株與短小芽孢桿菌(Bacillus pumilus)的親緣關系最緊密。通過研究得到該菌株的搖瓶發酵條件:產酶培養基為:蛋白胨3%、酵母膏1%、NaCl 0.5%、橄欖油1%,pH7.0,搖瓶發酵溫度為28℃,搖床轉速為180r/min,發酵周期為48~60h。所產脂肪酶在40℃、pH9.0時酶活性最高,對pH值和溫度的適應范圍較寬,pH6.0~10.0比較穩定,35~50℃具有較高酶活性。
耐有機溶劑;短小芽孢桿菌;脂肪酶;篩選
脂肪酶(EC3.1.1.3)是一類特殊的酰基水解酶,除能夠催化甘油酯類化合物的水解和合成外,還能在油水界面上催化酯的水解和合成、酯交換、生物表面活性劑的合成、多肽合成、高聚合物合成和手性藥物合成等有機合成反應,它可將甘油三酯分解成脂肪酸和甘油或進行其逆反應[1-3],在工業生產中有著廣泛應用。目前,脂肪酶在工業生產中被廣泛應用于食品與營養、輕工業、皮革、香料、化妝品、洗滌劑、有機合成、藥物合成等領域[4-5]。脂肪酶催化的許多反應是在非水相中進行的,由于許多微生物和其分泌的酶在有機溶劑中會被破壞和失活,限制了生物轉化在化學工業上的應用[6],因此從自然界中尋找耐有機溶劑、高活力的脂肪酶菌株,可為脂肪酶開發提供基礎。
芽孢桿菌脂肪酶具有來源廣泛、分子質量小、表達分泌能力強、非水相體系酶學性質突出、催化功能多、穩定性好和底物特異性多樣化等優點,是當前脂肪酶研究領域的熱點之一[7-10]。Hun等[11]利用苯與甲苯混合物作為碳源,成功篩選出耐有機溶劑的脂肪酶生產菌。Sameshima等[12]也從汽油污染的土壤中分離得到了耐苯菌株Rhodococcus opacus B-4,該菌株具有很好的有機溶劑耐受性,在有機溶劑中可存活5d并保持催化活性。本實驗以苯為唯一碳源,從環境樣品中篩選耐受有機溶劑脂肪酶活性較高的細菌,采用形態學特征、理化性質及16S rDNA基因序列鑒定其種屬,并對其酶學性質進行初步研究,為其進一步應用提供理論依據。
1 材料與方法
1.1 材料與試劑
本實驗采集的樣品有:鹽堿地土壤、花生地土壤、稻田土壤、金銀灘土壤、油井邊土壤、紅樹林根基土壤及紅樹林根部水樣,共7個樣本。
對硝基棕櫚酸苯酯(p-NPP) 美國Sigma 公司;橄欖油 天津口維可國際貿易有限公司;羅丹明B 上海試劑三廠;聚乙烯醇(PVA) 天津市巴斯夫化工有限公司;酚酞天津市大茂化學試劑廠;其他化學試劑均為國產分析純。
1.2 培養基
初篩培養基(g/L):Na2HPO43.5、KH2PO41.5、MgSO4·7H2O 0.5、NaCl 0.5,pH7.5,121℃高壓蒸汽滅菌20min,無菌條件下加入體積分數2.0%苯(0.22μm過濾除菌);富集培養基(g/L):Na2HPO43.5、KH2PO41.5、MgSO4·7H2O 0.5、NaCl 0.5、酵母粉 0.2,pH7.5,再加乳化橄欖油(將4g聚乙烯醇加熱溶于100mL蒸餾水中,然后與橄欖油以體積比3:1的比例混合,超聲攪拌乳化2min),121℃高壓蒸汽滅菌20min;油脂同化平板培養基(g/L):Na2HPO43.5、KH2PO41.5、MgSO4·7H2O 0.5、NaCl 0.5、瓊脂20,橄欖油乳化液體積分數12%、調至pH7.5,121℃高壓蒸汽滅菌20min,冷卻至45℃左右加羅丹明B,倒平板備用;LB平板培養基(g/L):蛋白胨10、酵母膏5、NaCl 5、瓊脂15、pH7.0,121℃高壓蒸汽滅菌20min,倒平板備用。
產脂肪酶發酵培養基(g/L):M1:可溶性淀粉25、橄欖油乳化液2.5、蛋白胨15,pH8.5;M2:胰蛋白胨20、豆油3.0、NaNO31、K2HPO41、MgSO45,pH8.0;M3:豆餅粉20、牛肉粉20、可溶性淀粉10、K2HPO45、NaNO35,pH7.0;M4:可溶性淀粉10、豆餅粉20、K2HP4O 5、NaNO32,pH8.5;M5:蛋白胨40、(NH4)2SO41、MgSO4·7H2O 1、K2HPO41,pH 8.0;M6:蛋白胨20、蔗糖10、橄欖油乳化液10、(NH4)2SO45、MgSO45、K2HPO42,pH8.0;M7:蛋白胨30、酵母膏10、NaCl 5、橄欖油10,pH7.0;M8:蛋白胨10、酵母膏5、NaCl 5,pH7.0。
1.3 酶活力測定
采用棕櫚酸-對硝基苯酯法測定[13]。
1.4 菌株的篩選
1.4.1 耐有機溶劑菌株的篩選
稱取樣品5g放入95mL無菌水中小火加熱至沸騰,維持1min,殺滅不產芽孢的細菌,靜置5min,取上清液5mL,加入初篩培養基中28℃、180r/min振蕩培養2d。觀察培養基渾濁度的變化。
1.4.2 產脂肪酶菌株的篩選
取上述渾濁度較大的培養液適量接種到富集培養基中,富集后取少量菌液涂布到油脂同化平板培養基,培養48h后在350nm紫外光照射下觀察菌落周圍熒光圈直徑的大小和變化,選取熒光圈較大的菌落,LB平板培養基劃線進行純培養,溫度28℃。
1.5 粗酶液酶學性質研究
M7培養基接種所篩選的H2菌株,28℃、180r/min振蕩培養2d,發酵液于4℃低溫10000×g離心20min,取上清液,用棕櫚酸-對硝基苯酯法測定脂肪酶酶活力,進行酶學性質研究。
1.5.1 有機溶劑耐受性能考察
在粗酶液中添加終體積分數為25%不同極性常數(logP)的有機溶劑,40℃、180r/min振蕩2h后,測定酶活力。以不加有機溶劑的原酶液為對照計算相對酶活力。
1.5.2 酶作用最適pH值及其pH值穩定性
分別用0.1mol/L HCl或0.1mol/L NaOH溶液調粗酶液pH值為6.0~10.0,40℃反應10min,測定酶活力,以確定該酶的最適pH值。用0.1mol/L HCl或0.1mol/L調粗酶液為不同pH值,4℃處理粗酶液12h,回調至9.0,40℃保溫10min,測定酶活力,以研究該酶的pH值穩定性。以最高酶活力作為100%計算相對酶活力,每組重復3次,結果取平均值。
1.5.3 酶作用最適溫度及其熱穩定性
取粗酶液,調pH9.0,分別于25~65℃水浴中反應10min,測定酶活力,以確定該酶的最適溫度。在不同溫度(40~80℃)條件下進行酶的熱穩定性研究,期間每隔10min測定酶活力,酶促反應條件同上,持續1h。以最高酶活力作為100%計算相對酶活力,每組重復3次,結果取平均值。
1.6 菌株的鑒定
形態學特征觀察和生理生化性質實驗參考文獻[14]方法,根據曾曉希等[15]的方法提取基因組DNA,以其為模板,采用16S rDNA通用引物,通過PCR擴增其16S rDNA基因序列,將擴增片段送上海生工生物工程技術服務有限公司測序,結果在NCBI中檢索比對,獲得有關種的公認標準序列數據后采用Clustal X 1.8軟件比對,使用MEGA 4.0進行序列同源性分析并繪制系統發育樹,算法為Neighbor-Joining。
2 結果與分析
2.1 耐有機溶劑菌株的篩選

表1 培養液渾濁度的變化Table 1 Change in culture turbidity during incubation of strains from different sources
培養液渾濁度變化見表1,結果顯示,稻田土壤和花生地土壤樣品在培養初期就變得渾濁,在第3天變得非常渾濁,表明這兩種樣品培養液中的微生物可以有效利用苯,并迅速生長,其他樣品中微生物利用苯效率較低。因此,選擇來自稻田地土壤和花生地土壤的微生物培養液進行脂肪酶產生菌的篩選。
在2017年結束之前,我們去了很多不同的火鍋店消費,評頭論足著每一家的優缺點,順便攝影。雖然離住家很近,不知不覺路過的機會也很高,而我再三猶豫,還是毅然決然的,放棄走入西瓜甜點涮涮鍋店內消費。
2.2 產脂肪酶菌株的篩選

表2 各菌株的產脂肪酶活力比較Table 2 Comparison of lipase activities produced by strains from different sources
采用油脂同化平板培養基篩選得到6株純培養菌株,分別命名為D1、D2、D3、H1、H2、H3。分別把6株菌接種于LB發酵培養基中,28℃、180r/min的搖床上培養48h后,取上清液測定酶活力。由表2可知,酶活力最高的是來自花生地土壤的菌株H2,選擇該菌株做進一步研究。
2.3 發酵培養基的選擇
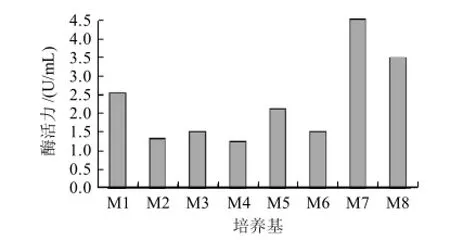
圖1 不同培養基對H2菌株所產脂肪酶的影響Fig.1 Effect of different culture media on lipase production by strain H2

表3 菌株H2粗酶對不同有機溶劑的耐受能力Table 3 Effect of organic solvents on the stability of crude lipase from strain H2
將H2菌株分別接種到發酵培養基M1、M2、M3、M4、M5、M6、M7和M8中,于28℃、180r/min振蕩培養48h,測定其粗酶液的酶活力。由圖1可知,以M7為發酵培養基時,脂肪酶酶活力最高,達到4.53U/mL,以下實驗均選擇M7作為發酵培養基。
2.4 發酵周期考察

圖2 不同發酵時間對H2菌株所產脂肪酶的影響Fig.2 Effect of fermentation time on lipase production by strain H2
取不同發酵時間的粗酶液進行酶活力測定,由圖2可知,發酵48h之前,酶活力上升趨勢比較明顯,發酵48h之后,酶活力變化趨于穩定,到72h酶活力開始略有下降。因此發酵時間選擇48~60h為宜。
2.5 搖床轉速和培養溫度對H2產脂肪酶的影響

圖3 搖床轉速和培養溫度對H2菌株產脂肪酶的影響Fig.3 Effect of rotational speed and temperature on lipase production by strain H2
2.6 酶學性質初步研究
2.6.1H2菌株粗酶液有機溶劑耐受性能
由表3可知,總體而言,隨著有機溶劑logP值的增加,對酶活性影響逐漸減小,說明菌株對有機溶劑的耐受性越好。粗酶對25%苯和甲苯的耐受性較差,苯是篩選H2菌株的壓力條件,說明菌株和其分泌的酶對有機溶劑的耐受性可以有所不同。細菌細胞有細胞壁和細胞膜作為屏障,阻礙了有機溶劑與胞內酶的直接接觸,而釋放到胞外的酶直接同有機溶劑作用,酶活力可能受到影響,如果酶的耐受性不夠,其活性會降低甚至喪失。
2.6.2H2菌株粗酶最適反應pH值及其pH值穩定性

圖4 pH值對H2菌株所產脂肪酶粗酶活性的影響Fig.4 Effect of pH on the activity of crude lipase from strain H2

圖5 pH值對H2菌株所產脂肪酶粗酶穩定性的影響Fig.5 Effect of pH on the stability of crude lipase from strain H2
由圖4可見,相對酶活力最高峰出現在pH 9.0處,表明H2菌株所產脂肪酶在pH值為9.0時酶活性最高。由圖5可見,H2菌株所產脂肪酶在pH 6.0~10.0范圍內比較穩定,當pH值低于5.0時,酶活性受到比較大的影響。
2.6.3 H2菌株粗酶最適反應溫度及熱穩定性

圖6 溫度對H2菌株所產脂肪酶粗酶活性的影響Fig.6 Effect of temperature on the activity of crude lipase from strain H2
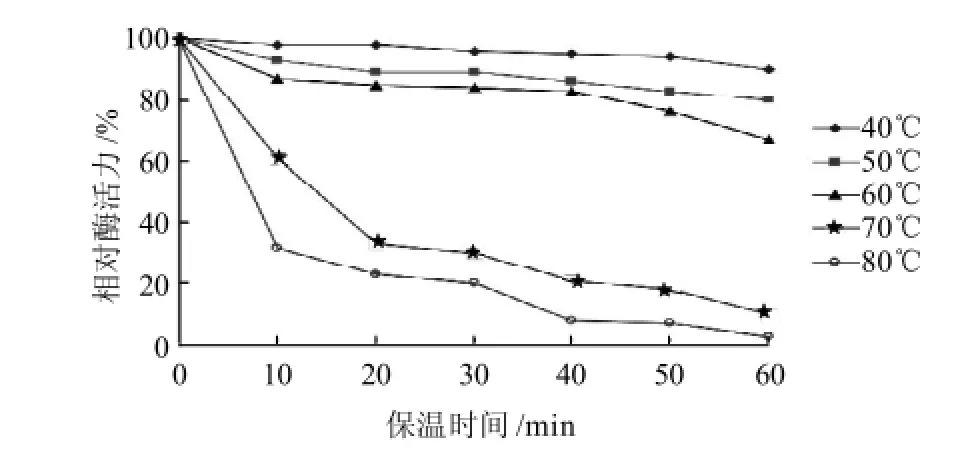
圖7 溫度對H2菌株所產脂肪酶粗酶穩定性的影響Fig.7 Effect of temperature on the stability of crude lipase from strain H2
由圖6 可見,相對酶活力在25~40℃之間隨著溫度的升高而增大,在40℃時達到最大,50℃以上相對酶活力逐漸降低。由圖7可見,H2菌株所產脂肪酶在60℃條件下保溫40min穩定性較好,70℃條件下保溫相對酶活力急劇下降,保溫20min相對酶活力僅為30%;而在80℃條件下僅保溫10min,酶活力就下降了70%,40min后相對酶活力幾乎為0,表明該酶在35~50℃之間具有較高的酶活性,H2菌株所產脂肪酶屬于中度耐熱脂肪酶。
2.7 H2菌株鑒定結果
2.7.1 主要生理生化特性
H2菌株在LB平板培養基上菌落呈圓形,白色,邊緣不整齊,表面粗糙,不透明;經革蘭氏染色,確定為革蘭氏陽性菌,菌體桿狀,好氧,中生芽孢(圖8);可利用D-葡萄糖、L-阿拉伯糖、D-甘露糖、蔗糖產酸;不消化瓊脂,水解明膠,不利用淀粉;利用檸檬酸鹽,硝酸鹽還原反應陰性,并可在含7%以下NaCl的LB培養基上生長;最適生長溫度28℃;接觸酶陽性;V-P實驗反應陽性。參照《伯杰細菌鑒定手冊》[14]初步鑒定為短小芽孢桿菌。

圖8 H2菌株菌落形態(a)和菌體形態(b)(×1600)Fig.8 Colony morphology of strain H2(×1600)
2.7.2 16S rDNA 的擴增
按1.5節方法對菌株H2的16S rRNA 序列擴增,瓊脂糖凝膠電泳后,在1000~2000bp之間可看到一條清晰條帶,約1500bp(圖9)。割膠回收目的片段進行測序。

圖9 H2菌株16S rRNA序列的PCR電泳圖Fig.9 Agarose gel electrophoresis of 16S rRNA sequence of strain H2
2.7.316 S rDNA序列同源性分析及系統進化樹的建立
經上海生工生物工程技術服務有限公司測序得到較完整的16S rDNA序列,與NCB1上已有序列進行Blast同源性檢索,發現其16S rDNA 和多數Bacillus pumilus的同源性達99%以上,對該序列構建系統進化樹,結果見圖10。發現進化樹上H2菌株和Bacillus pumilus屬于同一簇,遺傳距離幾乎為0,因此可初步判斷H2菌株的分類地位屬于Bacillus pumilus。同屬于Bacillus屬的枯草芽孢桿菌集中在另外一簇,而其他一些芽孢桿菌菌株則屬于獨立分支。
3 討 論
從花生地土壤分離篩選得到耐有機溶劑脂肪酶產生菌H2,經產酶發酵條件的初步研究,確定適宜的發酵培養基為M7,培養溫度為28℃,搖床轉速為180r/min,搖瓶發酵周期為48~60h。酶學性質初步研究結果表明:該脂肪酶在40℃,pH 9.0時酶活性最高,可見H2菌株所產脂肪酶屬于中度耐熱、耐堿脂肪酶;穩定性研究表明:該酶在pH 6.0~10.0范圍內比較穩定,在35~50℃之間具有較高的酶活性,說明此酶對溫度和pH值適應的范圍較寬,為其廣泛應用于食品工業提供了有利條件。脂肪酶在油脂加工、洗滌劑、皮革、造紙、食品、醫藥、化妝品、紡織等領域都有十分廣泛的應用。在食品工業中的應用主要集中在食用油脂改性、改善食品風味、改進食品質量及保健食品生產等方面。由于H2菌株所產脂肪酶可耐受部分有機溶劑,因此,在有機溶液中催化酯交換作用來改變低價脂肪性能,生產一些重要的多不飽和脂肪酸,利用菜子油生產生物柴油,在玉米油、葵花油、花生油、橄欖油和大豆油的生產等方面都具有潛在的應用價值。
通過分析H2菌株的16S rDNA基因序列,發現H2菌株與Bacillus pumilus具有最緊密的親緣關系。從形態學、生理生化特性及分子生物學水平上的鑒定結果可確定H2菌株為Bacillus pumilus。H2菌株作為工業生產菌所具有的優勢有:能形成抗性內生孢子——芽孢,有利于菌種的保藏,因其屬于常溫菌,所以可降低工業化成本,如發酵條件和設備要求等。因此,H2菌株具有開發耐熱、耐堿脂肪酶的潛在價值。該菌株的產酶能力還不能滿足工業生產要求,有必要通過誘變和遺傳育種的手段對H2菌株作進一步研究。
[1]彭立鳳, 趙汝淇, 譚天偉. 微生物脂肪酶的應用[J]. 食品與發酵工業, 2000, 26(3): 72-76.
[2]REETZ M T. Lipases as practical biocatalysts[J]. Curr Opin Chem Biol, 2002, 6(2): 145-150.
[3]PABLO D M, JOSE V S, TSAI S W, et al. Carica papaya lipase (CPL): an emerging and versatile biocatalyst[J]. Biotechnol Adv, 2006, 24: 493-499.
[4]UWE T B, COMELIUS B, RAMISETTI S, et al. Optimizing lipases and related enzymes for efficient application[J]. Trends Biotechnol, 2002, 20(10): 433-437.
[5]談重芳, 王雁萍, 陳林海, 等. 微生物脂肪酶在工業中的應用及研究進展[J]. 食品工業科技, 2006, 27(7): 193-195.
[6]詹喜, 王慶利, 鄒慧熙, 等. 有機溶劑中固定化酵母細胞催化香葉醇還原生物轉化的研究[J]. 浙江大學學報: 農業與生命科學版, 2006, 32(4): 391-395.
[7]ZHANG Jinhong, GUAN Rong, TAN Zhilei, et a1. Purification and properties of lipases/esterases from a Bacillus strain for enantioselective resolution of (S)-ketoprofen[J]. Artificial Cells Blood Substitutes and Biotechnology, 2005, 33(4): 435-445.
[8]DANDAVATE V, JINJALA J, KEHARIA H, et al. Production, partial purification and characterization of organic solvent tolerant lipase from Burkholderia multivorans V2 and its application for ester synthesis[J]. Bioresource Technology, 2009, 100: 3374-3381.
[9]van POUDEROYEN G, EGGERT T, JAEGER K E, et al. The crystal structure of Bacillus subtilis lipase: a minimal α/β hydrolase fold enzyme[J]. Journal of Molecular Biology, 2001, 309: 215-226.
[10]溫建新, 施碧紅, 吳偉斌, 等. 脂肪酶產生菌Bacillus subtilis FS321的分離鑒定及其產酶條件的初步優化[J]. 食品與發酵工業, 2008, 32(2): 37-41.
[11]HUN C J, RAHMAN R N Z A, SALLEH A B, et al. A newly isolated organic solvent tolerant Bacillus sphaericus 205y producing organic-solvent stable lipase[J]. Biochemical Engineering Journal, 2003, 15(2): 147-151.
[12]SAMESHIMA Y, HONDA K, KATO J, et al. Expression of Rhodococcus opacus alkB genes in anhydrous organic solvents[J]. Journal of Bioscience and Bioengineering, 2008, 106(2): 199-203.
[13]李俊, 王剛, 呂熹, 等. 響應面法優化耐有機溶劑脂肪酶營養條件[J].生物技術, 2010, 20(4): 66-69.
[14]希坎南R E, 吉本斯N E. 伯杰細菌鑒定手冊[M]. 中國科學院微生物研究所《伯杰細菌鑒定手冊》翻譯組, 譯. 北京: 科學出版社, 1989: 23-46.
[15]曾曉希, 周洪波, 劉飛飛, 等. 一株膠質芽孢桿菌的篩選和鑒定[J].湖南農業大學學報: 自然科學版, 2006, 32(3): 269-272.
Screening of Organic Solvent Tolerant Lipase-Producing Bacteria and Enzymatic Properties of Crude Lipase
LI Jun-feng1,LI Hong-fang1,DUAN Xiao-hui2,MEN Jin-ming1,SU Feng1
(1. College of Chemical Engineering, Qingdao University of Science and Technology, Qingdao 266042, China;2. Yantai Entry-Exit Inspection and Quarantine Bureau, Yantai 264000, China)
A moderately thermostable and alkali-resistant lipase-producing strain named as H2 was isolated from peanut plantation soil using 2% benzene by Rhodamine B plate assay and shake flask fermentation. It was preliminarily identified based on morphological and physiological characteristics. Meanwhile, the 16S rDNA gene of the strain was cloned and sequenced. The physiological and biochemical characterization and phylogenetic tree based on the 16S rDNA sequence showed that the strain was highly homologous to Bacillus pumilus. The optimal culture medium for lipase production by the strain was composed of 3% peptone, 1% yeast extract, 0.5% NaCl, 1% olive oil, pH 7.0, and the optimal fermentation conditions of temperature, rotational speed and time were 28 ℃, 180 r/min and 48ˉ60 h, respectively. The lipase produced under these conditions revealed the highest activity at 40 ℃ and pH 9.0. The lipase was stable over a broad range of pH (6.0ˉ10.0) and temperature (35ˉ50 ℃).
organic solvent tolerant;Bacillus pumilus;lipase;screening
Q939.96
A
1002-6630(2012)03-0116-05
2011-01-07
中國煙草總公司山東省公司科技計劃項目(KN172);國家質量監督檢驗檢疫總局科研基金項目(2007IK154)
李俊峰(1971—),男,講師,博士,研究方向為微生物代謝產物。E-mail:lijf1999@qust.edu.cn
