中原牡丹傳統品種的核型及進化關系1)
2012-06-28 09:12:02史倩倩
東北林業大學學報 2012年11期
史倩倩 王 雁 周 琳
(林木遺傳育種國家重點實驗室(中國林業科學研究院林業研究所),北京,100091)
黃國偉
(湖北林業科學研究院)
牡丹(Paconia suffruticosa Andr.)是芍藥科芍藥屬名貴的觀賞植物和藥用植物,作為中國傳統名花的優秀代表,其姿態萬千、色彩豐富,深受人們喜愛。中國中原牡丹品種數量最多,栽培歷史悠久、分布最廣,形態變異最豐富。牡丹品種分類的方法較多,目前以花型分類的方法得到了廣泛的應用,共分為2類 4 亞類 16 型[1]2-17。
核型是遺傳物質在細胞水平上的表征,與外部形態相比核型受外界環境影響小,是推測物種起源、演化、分類和鑒別的重要依據[2]。譚遠德等[3]在1993年提出,以核型數據為基礎的核型似近系數聚類法,能較好地克服傳統形態分析帶來的局限性,并做到了物種間核型相似的數量化,且多向、立體、多維地考察物種間的親緣關系,使結果客觀可靠。牡丹傳統品種[1]8-9有別于現代品種,具有更豐富的遺傳信息,但前人的相關研究均未區分傳統品種與現代品種。前人對牡丹野生種及少量栽培品種進行了細胞學研究[4]286-292,[5-17],探討了野生種及栽培品種的核型參數及野生種間的演化關系,但都未深入研究形態和核型參數之間的關系,沒有反映傳統品種及現代品種的進化關系及現代品種遺傳多樣性的丟失,并且涉及傳統品種的研究中品種類型的代表性不足。本試驗通過對21個典型不同花型的傳統中原牡丹品種核型進行比較和聚類分析,探討傳統品種間核型差異,進而探討染色體變異和花型之間的關系,旨在為牡丹品種演化研究提供線索,同時為深入研究牡丹傳統品種的細胞分類學問題奠定基礎。
1 材料與方法
試驗材料取自中國林業科學研究院玉泉山牡丹基地。21個中原牡丹傳統品種涵蓋了7個花型[1]52-207(表 1)。

表1 供試材料
2011年3月中下旬取直徑為0.5~1.5 cm的花蕾,剝除花瓣,只留花藥和子房,子房切開,用V(0.05%秋水仙素)∶V(0.02 mol·L-18 - 羥基喹啉)=1∶1混合液室溫下預處理5 h,經卡諾固定液(V(95%乙醇)∶V(冰乙酸)=1∶3)于4℃固定24 h,經95%乙醇沖洗2次后,轉入70%乙醇中,再放入1 mol·L-1鹽酸中,并于60℃恒溫金屬浴解離10~15 min,蒸餾水洗凈后,用卡寶品紅染色和常規壓片法制片,用蔡司AXIOIMAGER A1光學顯微鏡分別對各材料的50個染色體分散良好的中期分裂相進行拍照。
核型分析:每個品種選出形態清晰、伸展較佳的5個細胞[2],借助AXIOIMAGER軟件分別測量染色體及長短臂的大小。核型按Levan的分類法[18]和李懋學等[4]5-22的植物染色體標準化的規定分析;核型分類參照Stebbins[19]的核型分類標準;核型不對稱系數參照 Arano[20]公式。
聚類分析:按譚遠德等[3]提出的核型似近系數聚類分析方法和公式計算核型似近系數(λ);按吳昌謀[21]提出的公式計算核型進化距離(De),對似近系數利用平均聚類法(UPGMA)進行聚類。
2 結果與分析
2.1 21個中原傳統牡丹品種染色體核型的屬性特征與品種間的相關性
研究結果表明,供試21個品種的染色體核型穩定,但品種間存在一定的差異。21個中原牡丹品種的染色體照片及核型模式見圖1、圖2,主要核型參數見表2。從圖1、表2可以看出,除‘首案紅’是三倍體,有15條染色體外,其他全部為二倍體,染色體數目為10;除‘茄藍丹砂’的核型類型為2B型外,其余品種的核型類型均為2A,核型不對稱系數介于59.21% ~95.70%,屬于比較對稱的核型,說明‘茄藍丹砂’的進化程度相對較高。其觀察統計結果表明:只有‘脂紅’沒有發現隨體,其余20個品種均具有隨體染色體。其中‘藍田玉’含隨體最多,具有8個隨體染色體,分別分布在第1、3、4、5對染色體上。同時發現花型為皇冠型的牡丹品種具有的隨體染色體數目普遍較多。
21個傳統中原牡丹品種的核型公式有5種形式(表2):2n=6m+2sm+2st,有‘黃花魁’、‘酒醉楊妃’、‘茄藍丹砂’、‘大棕紫’、‘錦袍紅’、‘萬花盛’、‘姚黃’、‘趙粉’、‘魏紫’、‘小魏紫’、‘假葛巾紫’、‘桃紅獻媚’和‘青龍臥墨池’共13個品種;2n=8m+2st,有‘大棕紫’、‘白玉’、‘豆綠’、‘金玉交章’和‘藍田玉’共5個品種;2n=4m+6sm,品種為‘脂紅’;2n=8m+2sm,品種為‘赤龍煥彩’;2n=15=9m+3sm+3st,品種是‘首案紅’。
21個中原牡丹傳統品種的臂比均值為1.65~2.21,其中‘錦袍紅’的臂比均值最大,為 2.21,‘赤龍煥彩’的臂比均值最小,為1.65(表2)。表3、表4表明各花型之間的臂比均值差異顯著,薔薇型的臂比均值最大,為2.14;其次為菊花型,均值為2.04;千層臺閣型的臂比均值最小,為1.76。六大花型中樓子臺閣型的臂比均值方差最大,為0.024 0;菊花型的臂比均值方差最小,為0.000 1;說明樓子臺閣型內的臂比均值差異最大,其次為皇冠型、薔薇型、千層臺閣型、荷花型,而菊花型最一致。
通過6個花型的核型不對稱系數的比較得出(表3),6個花型間差異不顯著,核型不對稱系數為60.22~79.12。但花型內部的方差從大到小依次為皇冠型、樓子臺閣型、薔薇型、菊花型、荷花型和千層臺閣型。其中花型為皇冠型的‘首案紅’的核型不對稱系數明顯最大,為95.70%。
2.2 21個中原傳統牡丹品種親緣關系相關性的聚類分析
依據核型似近系數和核型進化距離的計算原理與方法計算出21種中原牡丹傳統品種兩兩之間的核型似近系數和進化距離,再利用平均聚類法(UPGMA)進行聚類,結果見圖3。
在21種中原牡丹傳統品種中,‘豆綠’與‘姚黃’的核型似近系數最大,為0.998 2,進化距離最小,為0.001 8,親緣關系最近;‘白玉’與‘魏紫’的核型似近系數最小,為0.852 6,進化距離最大,為0.159 5,親緣關系最遠。經統計,各供試材料間的核型近似系數均在0.850 0以上,進化距離間隔很小,這說明了21個中原牡丹傳統品種在核型上穩定性和相似性很高。由圖3可見,21個中原牡丹傳統品種在核型進化距離為0.0552、似近系數為0.9420處聚成兩大類:‘白玉’和‘魏紫’獨立于其他19個品種聚成一類。其中‘首案紅’在進化距離0.040 5、似近系數0.960 3時與其他18個品種聚成一類,而這18個品種在進化距離0.014 0、似近系數0.986 1時又分為兩大類:‘黃花魁’、‘桃紅獻媚’、‘大棕紫’和‘黑花魁’4個品種聚成一類,其他14個品種聚成一類。屬于托桂型的‘青龍臥墨池’和屬于薔薇型的‘錦袍紅’在似近系數0.975 6時聚為一類。7類花型21個中原牡丹傳統品種的核型聚類結果并沒有完全依據花型分類。
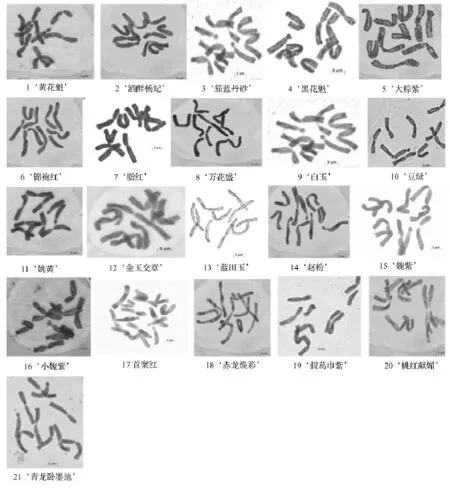
圖1 21個中原傳統牡丹品種有絲分裂中期圖
3 結論與討論
本研究通過對7類花型的21個中原牡丹傳統品種核型參數的研究,發現‘首案紅’的染色體為2n=15,三倍體;其他20個品種的染色體為2n=10,二倍體,與前人研究的結果相吻合[17],同時說明了‘首案紅’的進化較其余品種高。
楊滌清等[7]和張贊平等[11,15]都認為牡丹種間核型的差異主要表現在隨體的數目和位置上,并認為隨體數目及隨體染色體的雜合性是牡丹種間差異的一個重要的細胞學特征。本試驗的21個品種中,只有‘脂紅’沒有發現隨體,說明該品種的進化程度相對其他品種有所不同。隨體位置主要涉及到第1、2、4、5對染色體短臂上,而‘藍田玉’、‘假葛巾紫’和‘桃紅獻媚’在第3對染色體的短臂也出現了隨體,說明這3個品種的進化程度高。觀察發現花型為皇冠型的品種具有隨體的數目較多。因此,隨體數量及位置的差異是否反映了供試21個品種起源的差異有待進一步研究。

圖2 21個中原傳統牡丹品種的染色體核型模式圖
著名植物分類學家和進化學家 Stebbins[19]提出,被子植物核型進化的基本趨勢是由對稱向不對稱發展的。在系統演化上處于較古老或原始的植物,往往具有較對稱的核型,而不對稱的核型則通常出現在較進化或特化的植物中。本試驗中的20個牡丹傳統品種核型類型是較為對稱的2A型,只有‘茄藍丹砂’的核型類型為2B型,說明‘茄藍丹砂’比其他20個品種進化程度高,可能相對而言產生的年代要晚一些。
依據侯小改等[17]的牡丹進化觀點,牡丹品種的核型存在著多樣性,這也許跟牡丹擁有多種花型及花型易變異的現象有著密切的聯系。得出以下結論:具有2n=6m+2sm+2st的品種不對稱性最高;具有2n=4m+6sm和2n=8m+2st的品種不對稱性次之;具有2n=8m+2sm的品種最原始,即‘赤龍煥彩’。核型不對稱系數是反映染色體對稱與否、進化與否的另一個參數指標[4]171。在本試驗中核型不對稱系數以‘首案紅’最大,而‘赤龍煥彩’則屬于較小的,說明‘赤龍煥彩’的產生時間早于其他供試品種,較原始,而‘首案紅’比較復雜,是相對進化或原始,有待進一步研究。
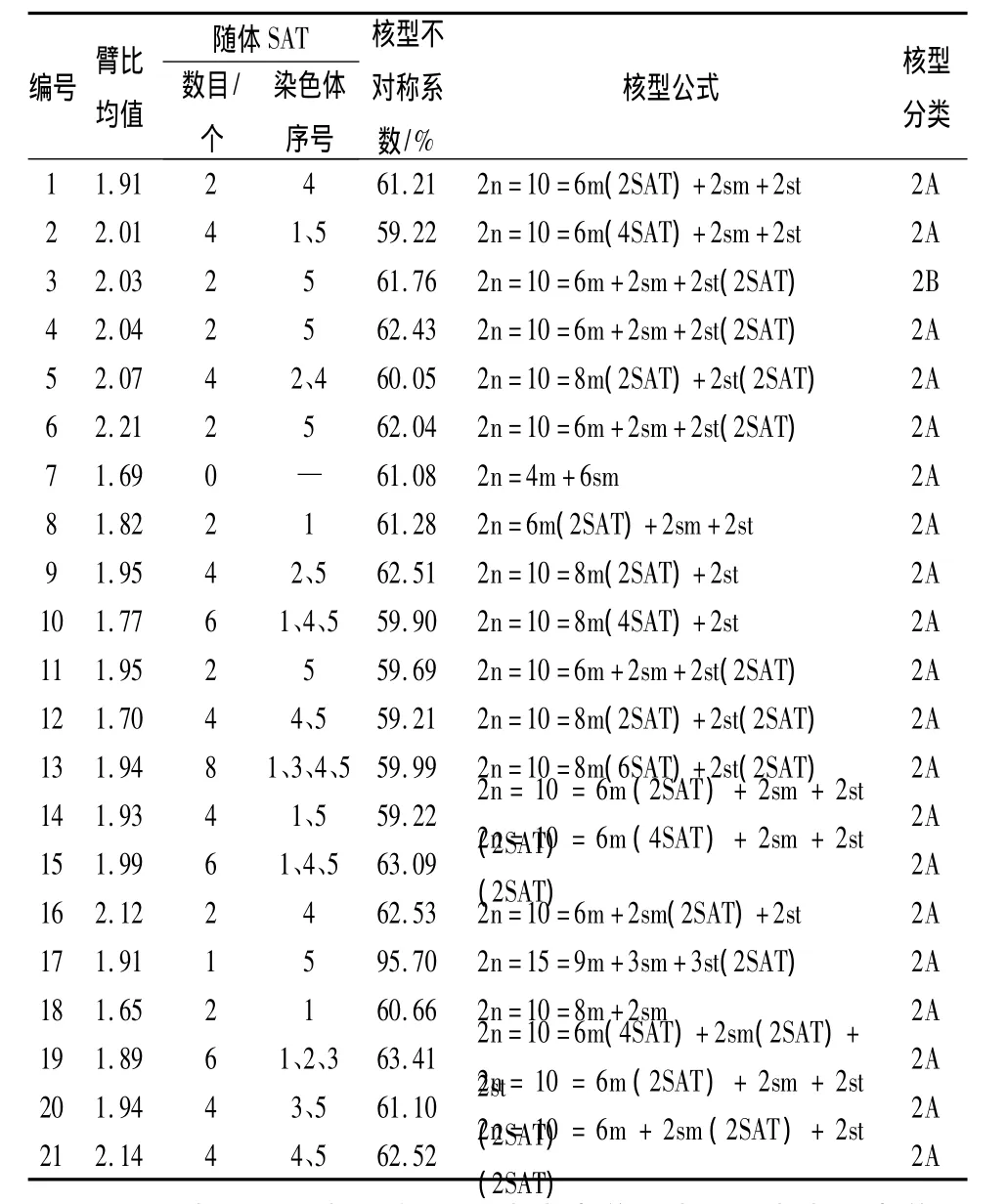
表2 21個中原傳統牡丹品種的主要核型參數
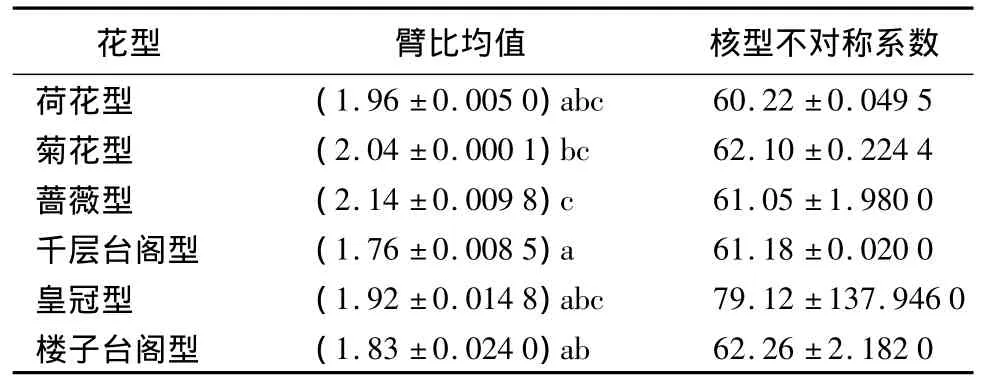
表3 6個花型染色體臂比均值、核型不對稱系數方差分析及多重比較
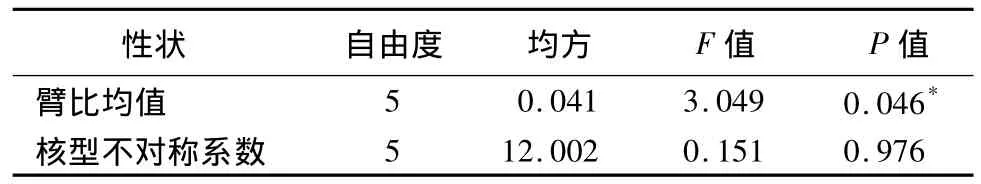
表4 6個花型間染色體臂比均值、核型不對稱系數方差分析
傳統品種花型間的核型不對稱系數差異不顯著,由低到高依次為:荷花型、薔薇型、千層臺閣型、菊花型、樓子臺閣型、皇冠型,這與王蓮英[1]14-19提出牡丹品種按花型演進的途徑的觀點基本一致,而試驗中的菊花型和千層臺閣型的品種沒有遵循傳統的花型演進途徑,極有可能是因為這2個花型的品種栽培年代晚一些或栽培條件不同。傳統品種花型內部的核型不對稱系數差異較大,從小到大依次為千層臺閣型、菊花型、荷花型、薔薇型、樓子臺閣型和皇冠型,說明千層臺閣型品種的核型類型較一致,而皇冠型品種的核型類型差異最大。
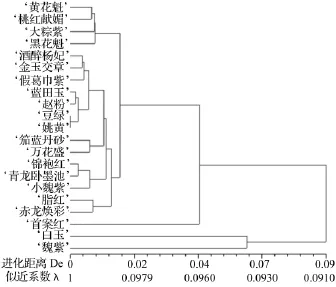
圖3 21個中原傳統牡丹品種核型似近系數聚類圖
臂比均值體現的是一個核型內所有染色體整體的不對稱性。觀察的21個品種中‘赤龍煥彩’臂比均值(1.65)最小,核型較對稱,跟核型類型分析的結果一致,也證明了其原始性。還發現各花型之間的臂比均值差異顯著,菊花型的臂比均值較大,說明該花型的核型不對稱性。通過各花型的臂比均值方差比較得到菊花型的核型類型最一致,樓子臺閣型和皇冠型一致性最差,與核型不對稱系數得出的結論基本吻合,表明材料中的菊花型品種進化程度較高。
依據數值分類學原理和似近分析理論得出的核型似近系數及核型進化距離能夠多向、立體、多維地考察物種間的相似性,從而判斷物種間的親緣關系和遺傳距離[2]。目前核型似近系數聚類分析方法應用較少,僅分析過蝗蟲、雞、禽類、豬、牦牛、榛子、風毛菊屬植物、紫花苜蓿、青稞和大花蕙蘭,發現聚類結果與品種分化歷史基本一致[2,22-30]。文中對21個中原牡丹傳統品種的親緣關系與進化距離進行了分析,結果與傳統分類結果不完全相同。似近系數為0.850 0以上,進化距離間隔小,這說明了21個中原牡丹傳統品種在核型上穩定性和相似性很高。同屬于皇冠型的‘豆綠’和‘姚黃’,‘藍田玉’和‘趙粉’親緣關系最近,遺傳背景相似。在進化距離為0.014 0、似近系數為 0.9861 時,‘黃花魁’、‘桃紅獻媚’、‘大棕紫’和‘黑花魁’4個品種聚成一類。而這4個品種分屬于4個不同的花型。屬于托桂型的‘青龍臥墨池’和屬于薔薇型的‘錦袍紅’在似近系數為0.975 6時聚為一類。‘首案紅’在似近系數0.960 3處自成一組,這充分說明其在進化過程中的特殊性。文中的研究結果顯示,在細胞學水平上大致符合牡丹組依據花型分類的結果,可能原因是由于這些傳統品種栽培年代、條件不同,也許經品種雜交選育,花型容易發生變異,核型分析中的人為誤差以及核型似近系數聚類分析方法的適用性等。因此,中原牡丹傳統品種分類和鑒定問題及細胞學與形態學之間的關聯還需進一步驗證。
[1]王蓮英.中國牡丹品種圖志[M].北京:中國林業出版社,1997.
[2]徐琪,陳國宏,張學余,等.3個地方雞種的核型及其似近系數分析[J].畜牧獸醫學報,2004,35(4):362 -366.
[3]譚遠德,吳昌謀.核型似近系數的聚類分析方法[J].遺傳學報,1993,10(4):21 -25.
[4]李懋學,張贊平.作物染色體及其研究技術[M].北京:中國農業出版社,1996:287.
[5]李懋學,張效方.三倍體牡丹的細胞遺傳學觀察[J].遺傳,1982,4(5):19 -21.
[6]張贊平.栽培牡丹的核型研究[J].豫西農專學報,1988(2):5-12.
[7]楊滌清,朱燮桴.草芍藥、野牡丹和黃牡丹的核型研究[J].云南植物研究,1989,11(2):139 -144.
[8]李思峰,于兆英,周俊彥.黃牡丹的核型分析[J].武漢植物研究,1989,7(2):107 -111.
[9]肖調江,龔洵,夏麗芳.滇牡丹復合群的Giemsa C-帶比較研究[J].云南植物研究,1997,19(4):395 -401.
[10]裴顏龍.牡丹復合體的研究[D].北京:中國科學院植物研究所,1993:57.
[11]張贊平,侯小改.楊山牡丹的核型分析[J].遺傳,1996,18(5):3-6.
[12]于兆英,李思峰,周俊彥.珍稀植物:紫斑牡丹和矮牡丹的核型分析[J].西北植物學報,1989,7(1):12 -16.
[13]洪德元,張志寬,朱相云.芍藥屬研究(Ⅰ):國產幾個野生種核型報道[J].植物分類學報,1988,26(1):33 -43.
[14]王蓮英,劉淑敏,秦魁杰,等.牡丹及其栽培品種的染色體組型[J].北京林學院學報,1983(1):63 -69.
[15]張贊平,李懋學,袁甲正.牡丹染色體的Ag-NoR和Giemsa C-帶研究[J].武漢植物學研究,1990,8(2):101-105.
[16]于玲,何麗霞,李嘉玨.甘肅紫斑牡丹與中原牡丹類群染色體比較研究[J].園藝學報,1997,24(1):79 -83.
[17]侯小改,段春燕,劉素云,等.中國牡丹染色體研究進展[J].中國農學通報,2006,22(2):307 -309.
[18]Levan A,Fredga K,Sandberg A A.Nomenclature for centromere position on chromosome[J].Hereditas,1964,52(3):201 -220.
[19]Stebbins G L.Chromosome evolution in higher plants[M].London:Academicn Press,1971:87 -123.
[20]Arano H.Cytological studies in subfamily Carduoideae of Japan IX[J].Bot Mag,1963,76:32.
[21]吳昌謀.核型似近系數和進化距離的估計[J].動物分類學報,1996,21(3):338 -344.
[22]姚世鴻,吳昌謀.三科五屬18種蝗蟲的核型似近系數聚類分析研究[J].貴州師范大學學報:自然科學版,2004,22(3):19 -24.
[23]熊家軍.七種禽類的核型似近系數聚類分析和雞W染色體相對長度與其主要經濟性狀的相關分析[D].武漢:華中農業大學,2002.
[24]吳圣龍,包文斌,束靖婷,等.安徽地方豬種間核型似近系數和進化距離的聚類分析[J].中國畜牧獸醫,2006,42(21):4-6.
[25]鐘金誠,陳智華,張成忠.牦牛品種(類群)間核型似近系數分析[J].草食家畜,1996(3):11-13.
[26]郭媛媛,邢世巖,馬穎敏,等.15種榛子種質的染色體核型分析[J].園藝學報,2009,36(1):27 -32.
[27]喬永剛,宋蕓.15種風毛菊屬植物核型似近系數聚類分析[J].中國草地學報,2011,33(2):38 -43.
[28]張雪婷,楊文雅,楊芳萍,等.6個紫花苜蓿材料的核型及其親緣關系分析[J].西北植物學報,2011,31(4):671 -676.
[29]段瑞君,任又成,熊輝巖.4種青海青稞品種的染色體核型分析[J].中國農學通報,2010,26(15):78 -82.
[30]王利民,王四清,董曉宇,等.核型似近系數在大花蕙蘭雜交育種中的應用初探[J].河南農業科學,2010(8):106-113.
