固態混菌發酵生產飼用復合酶制劑營養條件和培養條件的研究
2012-06-29 10:27:02浙江省農業科學院植物保護與微生物研究所許少春李艷麗許堯興
中國飼料 2012年2期
關鍵詞:生產
浙江省農業科學院植物保護與微生物研究所 許少春 李艷麗 柳 永 許堯興*
研究表明,飼料中添加復合酶制劑的飼喂效果優于單一酶制劑(李吉祥等,2007)。然而現在的復合酶一般是由單一酶復配而來的,應用成本較高。因此,研究復合酶的高效生產方法勢在必行。本文在前期篩選獲得高產α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶的單一黑曲霉菌株的基礎上,進一步對混合菌進行固態發酵條件研究,為復合酶制劑的高效產業化生產提供理論依據。
1 材料和方法
1.1 菌種與培養基 黑曲霉變種(A.niger v.Tiegh CGMCC1182)為 α-半乳糖苷酶(α)生產菌株,酶活力可達245 IU/g(許堯興等,2009);黑曲霉MA-56(A.niger MA-56 CGMCC2722)為 β-甘露聚糖酶(β)生產菌株,酶活力可達1101 IU/g(李艷麗等,2009);黑曲霉 XY-1(A.niger XY-1 CGMCC1182)為木聚糖酶(X)生產菌株,酶活力可達9478 IU/g(Xu 等,2008)。
斜面保藏培養基(PDA):馬鈴薯 200 g/L,蔗糖20 g/L,瓊脂20 g/L,自然pH。
混菌發酵基礎培養基(M-M),根據三種酶單一生產的培養基配方進行組合確定:麩皮8 g,豆粕 2 g,玉米芯 2 g,魔芋粉 0.1 g,葡萄糖 0.5 g,(NH4)2SO40.2 g,NaNO30.2 g,MgSO40.1g,KH2PO40.2 g,H2O 11 mL。進行營養因子條件試驗時,各成分的含量根據實際情況確定。
1.2 培養條件 將在PDA斜面上生長4~5 d的三種酶制劑的黑曲霉菌株的孢子用30 mL無菌水洗脫,并按1∶1∶1的比例制成混合孢子懸浮液。吸取2 mL孢子懸浮液至裝有11 g(按干基質)發酵培養基的300 mL三角瓶中,并攪拌均勻。28~30℃培養56 h。
1.3 酶活測定
1.3.1 粗酶液制備 將發酵后的培養物于40℃烘干,粉碎。稱取一定量的固體酶樣,加入適量的水,40℃水浴抽提30 min,用濾紙過濾,濾液即為粗酶液。依據發酵產物中α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶的酶活高低,粗酶液再作適當的二次稀釋供測定。
1.3.2 酶活測定 α-半乳糖苷酶采用許堯興等(2004)的pNPG法并略作修改。取經適當稀釋的酶液0.5 mL,加入0.1 mol/L pH4.0的磷酸氫二鈉-檸檬酸緩沖液1.0 mL,5 mmol/L對硝基苯-D-α-半乳糖苷(pNPG,Sigma)0.5 mL,于 40 ℃水浴精確反應10 min,加入0.5 mol/L Na2CO3溶液2.0 mL終止反應,于400 nm處測分光光度值。酶活力定義:在上述條件下,每分鐘分解底物生成1 μmol對硝基苯酚所需的酶量為1個酶活單位(IU/g)。β-甘露聚糖酶和木聚糖酶酶活測定采用DNS法,測定條件為pH 5.5和40℃。一個單位的酶活定義為在測定條件下1 min內水解底物產生相當于1 μmol還原糖的酶量。
1.4 數據統計 運用SAS軟件對數據進行統計分析,并在0.05水平上進行顯著性檢驗。
2 結果與討論
2.1 發酵培養基組成對黑曲霉混合菌發酵生產復合酶的影響
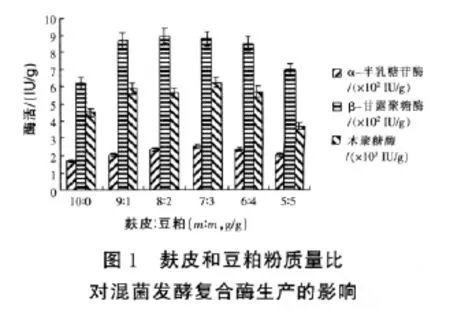
2.1.1 發酵培養基中麩皮、豆粕質量比對固態混菌發酵復合酶生產的影響 在發酵基礎培養基中,其他因素不變通過改變麩皮與豆粕粉的質量比對復合酶產量的影響見圖1。結果表明,麩皮與豆粕粉質量比在7∶3時,α-半乳糖苷酶酶活達到最大251 IU/g;麩皮與豆粕粉質量比在9∶1~6∶4時,β-甘露聚糖酶和木聚糖酶的活力都較高,且酶活相差不大,說明在一定范圍內麩皮與豆粕粉的質量比對β-甘露聚糖酶和木聚糖酶的活力影響不大。綜合α-半乳糖苷酶酶、β-甘露聚糖酶和木聚糖酶三種酶活力和價格因素,在以后的試驗中選用麩皮與豆粕粉質量比為7∶3,三種酶制劑的活力分別可以達到 251、879、6144 IU/g。
2.1.2 玉米芯添加量對固態混菌發酵復合酶生產的影響 絕大多數產木聚糖酶菌株的產酶可以被木聚糖所誘導。玉米芯中木聚糖的含量可達35%以上,但玉米芯的添加對α-半乳糖苷酶和β-甘露聚糖酶的生產影響卻是未知的。因此,以培養基中麩皮與豆粕粉質量比7∶3為基礎,其他因素不變,研究了玉米芯不同用量對混菌發酵復合酶生產的影響,結果見圖2。由圖2可見,玉米芯的添加對α-半乳糖苷酶和β-甘露聚糖酶的生產有抑制作用,當培養基中玉米芯添加量在1.0 g以下時,二者酶活均平穩下降,而隨著玉米芯添加量的逐漸增大,酶活呈急劇下降的趨勢;但玉米芯的添加卻對木聚糖酶的生產有激活作用,當培養基中玉米芯添加量在0.5 g時,木聚糖酶酶活顯著上升,當玉米芯添加量在1.0 g時,木聚糖酶酶活仍繼續上升,而隨玉米芯添加量繼續增加,木聚糖酶酶活基本趨于穩定。綜合考慮玉米芯添加量在1.0 g時,α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶三種酶活力可以達到酶活最優化,此時,三種酶制劑的活力分別可以達到225、781、7656 IU/g。

2.1.3 魔芋粉添加量對固態混菌發酵復合酶生產的影響 大多數微生物的β-甘露聚糖酶是誘導酶,只有在培養基中含有β-甘露聚糖時才進行β-甘露聚糖酶的合成。魔芋粉中含有44%~64%葡甘露聚糖,可以誘導β-甘露聚糖酶的生產,但對α-半乳糖苷酶和木聚糖酶的影響尚未考察。因此,根據2.1.1和2.1.2的結果,采用麩皮7 g,豆粕3 g,玉米芯粉1.0 g,其他因素不變,作為進一步進行研究的固態發酵培養基,對混菌發酵培養基中魔芋粉的添加量對復合酶生產的影響進行研究,結果見圖3。由圖3可見,混菌發酵培養基中添加魔芋粉,對黑曲霉α-半乳糖苷酶和木聚糖酶的酶活無顯著影響;而培養基中添加0.1 g魔芋粉與不添加魔芋粉相比,β-甘露聚糖酶酶活力顯著提高,但隨魔芋粉的添加量繼續增加β-甘露聚糖酶酶活基本趨于穩定,因此在以麩皮與豆粕粉質量為10 g為標準時,魔芋粉添加量為0.1 g,此時酶活力可以達到862 IU/g。

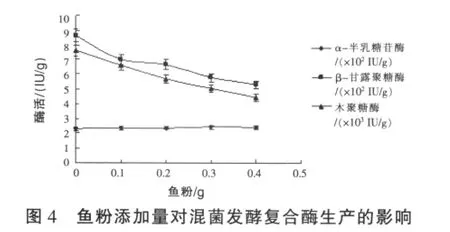
2.1.4 魚粉添加量對固態混菌發酵復合酶生產的影響 黑曲霉α-半乳糖苷酶的純菌發酵中,由于發酵結束之后,孢子數量多,在進行磨粉處理時會造成孢子在空氣中揮散。經過試驗,在發酵培養基中添加適量的魚粉,可以在一定程度上抑制黑曲霉孢子的生長,但又不會造成α-半乳糖苷酶酶活的降低。因此,進一步對混菌發酵培養基中魚粉的添加量對復合酶生產的影響進行研究,結果見圖4。由圖4可見,混菌發酵培養基中添加魚粉,對α-半乳糖苷酶酶活沒有影響,但卻強烈抑制了β-甘露聚糖酶和木聚糖酶的酶活,雖然發酵產品烘干后孢子數量減少,但也相應降低了復合酶的酶活,由此可見,在黑曲霉混菌發酵進行復合酶生產時,混菌發酵培養基中不需要添加魚粉。
2.1.5 無機氮源(NH4)2SO4和NaNO3對固態混菌發酵復合酶生產的影響 少量的NaNO3和(NH4)2SO4對黑曲霉木聚糖酶生產都有激活作用,而且NaNO3的誘導效果及其顯著;NaNO3和(NH4)2SO4對黑曲霉β-甘露聚糖酶的生產都有輕微的抑制作用;NaNO3對黑曲霉α-半乳糖苷酶的生產有抑制作用,而(NH4)2SO4對α-半乳糖苷酶的生產卻有激活作用 (許堯興等,2009)。通過試驗,確定 NaNO3和(NH4)2SO4的最適添加濃度,使混菌發酵的復合酶酶活達到最優化,結果見圖5和圖6。綜合考慮α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶三種酶活力,當(NH4)2SO4添加量為
0.2 g,NaNO3添加量為0.1 g時,三種酶的酶活力可以得到最優化,此時三種飼用酶活力分別可以達到 216、772、9244 IU/g。

綜合分析,黑曲霉混菌發酵生產飼用復合酶制劑的優化培養基配方為 (以麩皮和豆粕總量為10 g)計算:麩皮 7.0 g,豆粕 3.0 g,玉米芯 1.0 g,魔 芋 粉 0.1 g, 葡 萄 糖 0.5 g, (NH4)2SO40.2 g,NaNO30.1 g,MgSO40.1g,KH2PO40.2 g,H2O 11 mL。以下的培養條件試驗就是以優化好的培養基配方為基礎。

2.2 培養條件對黑曲霉固態混菌發酵生產復合酶的影響 前期研究表明,黑曲霉菌株純菌發酵生產α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶時,培養溫度、培養基含水量和裝載量條件是一致的,在培養溫度30℃時,當固形物與加水比1∶1,300 mL三角瓶中裝量8 g培養基時各酶活達到最大值 (許堯興等,2009; 李艷麗等,2009;Xu等,2008)。因此,在研究培養條件對黑曲霉混菌發酵生產復合酶的影響時不予改變。
2.2.1 接種比例對固態混菌發酵復合酶生產的影響 純菌發酵時接種量的大小影響發酵的產量和發酵周期,接種量小,延長菌體延遲期,發酵周期長,易染菌;但接種量過高往往又會使菌體生長過快,代謝產物積累過多,反而會抑制后期菌體的生長。混菌發酵進行飼用復合酶生產時,不同的接種比例也會對復合酶制劑當中單一酶的酶活產生影響,結果見圖7。由圖7可知,不同接種比例對復合酶中α-半乳糖苷酶的酶活基本無影響;當α∶β∶X 為 4∶5∶5時,β-甘露聚糖酶的酶活達到最高值883 IU/g,其次當 α∶β∶X 為 5∶6∶6 時,β-甘露聚糖酶的酶活可以達到 813 IU/g; 當 α∶β∶X 為 1∶2∶2時,木聚糖酶的酶活達到最高值9900 IU/g,其次當 α∶β∶X 為 5∶6∶6 時,木聚糖酶的酶活可以達到9767 IU/g。綜合考慮α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶三種酶活力,當 α∶β∶X 為 5∶6∶6 時,可以達到酶活最優化,此時,三種酶制劑的活力分別可以達到 201、813、9767 IU/g。

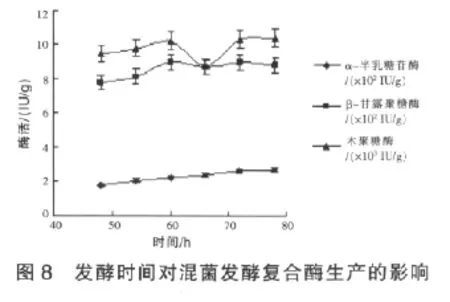
2.2.2 發酵時間對混菌發酵復合酶生產的影響在獲得了混菌發酵復合酶生產菌株接種比例的基礎上,研究了不同培養時間對菌株合成飼用復合酶的影響。接種后從培養48 h起,每6 h取樣,測定不同培養時間的各單酶產量,結果見圖8。由圖8可見,在試驗的時間范圍內,隨著發酵時間的延長,α-半乳糖苷酶的酶活呈不斷上升趨勢,其中78 h時的酶活比60 h時的酶活提高了20%;β-甘露聚糖酶酶活在60 h之后達到了穩定;木聚糖酶則在60 h時達到第一個酶活高峰,此后隨著時間延長,木聚糖酶酶活先是下降,66 h之后酶活又呈上升趨勢,在72 h后酶活也達到了穩定。雖然木聚糖酶酶活出現兩個產酶高峰,但第二個產酶高峰比第一個產酶高峰時間上延長了12 h以上,但單位酶活僅提高2%。綜合分析發酵效率和發酵產量,將菌株復合發酵生產復合酶的時間定為60 h,此時三種酶制劑的活力分別可以達到221、894、10188 IU/g。
綜合分析,黑曲霉混菌發酵生產飼用復合酶制劑的產酶最適培養條件為:培養溫度30℃,固形物與加水比為 1∶1,α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶接種比例為5∶6∶6,接種混合孢子懸浮液2.5 mL(以一支菌種斜面加30 mL無菌水為標準),300 mL三角瓶中裝量8 g培養基,發酵60 h時,復合酶產量達到最優,α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶三種酶制劑的活力分別可以達到 221、894、10188 IU/g。
3 討論
混菌發酵指采用兩種或多種微生物的協同作用共同完成某發酵過程的一種新型發酵技術,是純種發酵技術的新發展,也是一種不需要進行復雜的DNA體外重組就可取得類似效果的新型發酵技術,其優點是可提高發酵效率甚至可形成新產品,因此,得到了廣泛的應用(方浩等,2009;胡奎娟等,2007)。本試驗中的三株黑曲霉菌株,雖然具有相似的遺傳背景但卻具有不同的生產性狀,分別為α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶高產菌株,酶活分別可以達到245、1101、9478 IU/g。通過混菌發酵營養條件和培養條件研究,通過單次混菌發酵,發酵產物中α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶的酶活分別可以達到221、894、10188 IU/g。 與純菌發酵相比,混菌發酵產品中相對單一酶來講,只有木聚糖酶酶活提高了7.50%,而α-半乳糖苷酶和β-甘露聚糖酶酶活分別比純菌發酵降低了9.80%和18.80%。目前,在飼料中添加復合酶制劑成為一個趨勢,但現在的復合酶一般是由單一酶復配而來的,提高了應用的成本。從表觀上看,本次研究混菌發酵對試驗的三種酶活的提高并沒有起到作用,但從復合酶角度來講,若想得到含有α-半乳糖苷酶、β-甘露聚糖酶和木聚糖酶的復合酶制劑產品,勢必要對三種純菌發酵的酶制劑進行復配,這樣復配產品的獲得一是需要進行三次純菌發酵過程,二是通過復配之后(假設按 1∶1∶1進行配制),復配產品中單一酶制劑比純菌發酵的酶制劑產品酶活要降低。對混菌發酵而言,復合酶產品只需要進行一次發酵過程,而且發酵產品中不但木聚糖酶酶活比純菌發酵要提高,而且α-半乳糖苷酶和β-甘露聚糖酶酶活也只比純菌發酵降低了9.80%和18.77%,優勢明顯。
[1]方浩,宋向陽,趙晨,等.里氏木霉與黑曲霉混合發酵產纖維素酶的研究[J].林產化學與工業,2009,29(122):15 ~ 19.
[2]胡奎娟,吳克,潘仁瑞,等.固態混合發酵提高木聚糖酶和纖維素酶活力的研究[J].菌物學報,2007,26(2):273 ~ 278.
[3]李吉祥,夏先林,郁建生.飼用酶制劑對早期斷奶仔豬作用研究進展[J].銅仁職業技術學院學報,2007,5(3):34 ~ 37.
[4]李艷麗,許少春,柳永.黑曲霉MA-56 β-甘露聚糖酶的生產條件研究[J].中國飼料,2009,16:11 ~ 14.
[5]許堯興,姚曉紅,許少春.兩種測定程序對飼用α-半乳糖苷酶活性檢測結果的比較[J].浙江農業學報,2004,16(6):349 ~ 353.
[6]許堯興,李艷麗,許少春,等.培養基組成及發酵條件對黑曲霉變種產α-半乳糖苷酶的影響[J].浙江大學學報(農業與生命科學版),2009,35(4):401~408.
[7]Xu Y X,Li Y L,Xu S C.et al.Improvement of xylanase production by Aspergillus niger XY-1 using response surface methodology for optimizing the medium composition[J].Journal of Zhejiang University Science B,2008,9(7):558~566.
猜你喜歡
江蘇安全生產(2022年9期)2022-11-02 07:01:24
中國化肥信息(2022年7期)2022-08-31 01:28:54
山東冶金(2022年2期)2022-08-08 01:50:42
小學科學(學生版)(2020年10期)2020-10-28 07:52:12
中國化肥信息(2020年7期)2020-03-19 01:54:02
中國軍轉民(2017年6期)2018-01-31 02:22:28
消費導刊(2017年24期)2018-01-31 01:29:23
中國制筆(2017年2期)2017-07-18 10:53:09
現代企業(2015年4期)2015-02-28 18:48:06
汽車零部件(2014年11期)2014-09-18 11:57:16
