不同種質翅果油樹表型及抗旱性比較研究
2012-07-31 03:30:48宮慧芳
浙江農業科學 2012年3期
宮慧芳,陳 惠
(山西師范大學 生命科學學院,山西 臨汾 041004)
翅果油樹(Elaeagnus mollis Diels.)屬胡頹子科、胡頹子屬的木本油料植物,目前被列為國家重點保護野生植物名錄,屬國家二級保護植物。自然分布狹窄僅限于山西南部的五縣和陜西的戶縣[1-2]。翅果油樹為早春蜜源植物,種仁中含有較高含量的不飽和脂肪酸亞油酸和高含量的維生素E,且出油率高、油質好,營養價值豐富[3];其根系發達,根部具根瘤菌,可改良土壤、增加土壤養分[4],因而具有較高的經濟價值和生態效益,開發和利用前景廣闊。
前人根據翅果油樹果形分為3個生態類型,即長果型、大宮燈和小宮燈。然而,近年來山西琪爾康生物公司每年秋季從山西南部5縣收集翅果油樹果實,根據中果皮顏色、果實大小、形狀等的差異共鑒定出222份資源品種,陳惠等[5]對其中產地為翼城的2個翅果油樹種質的果實表型特征以及幼苗生長發育進行過初步研究,關于其他產地不同種質翅果油樹果實表型特征、幼苗表型差異及種質間抗旱生理特性的比較研究還鮮見報道。
本研究選取產地為鄉寧的果實形態不同的3個翅果油樹種質,除對其果實和幼苗的表型進行觀察比較外,還對干旱脅迫下3個種質的葉片保水力、質膜透性、葉綠素含量、超氧化物歧化酶(SOD)、過氧化物酶(POD)、游離脯氨酸進行分析比較。對了解翅果油樹種質之間的表型差異,幼苗早期的品種分類、鑒定、選育、栽培留種有一定的借鑒。
1 材料與方法
1.1 材料
2009年9月在山西省鄉寧縣采集了3個翅果油樹種質的果實,去掉最外層的膜質果皮,剩下帶有中果皮的果實常溫下保存備用。2010年5月20日,在山西師范大學生命學院溫室進行種子萌發實驗,6月20日3個種質分別選取生長一致的幼苗在組織培養實驗室進行生理指標的測定。
1.2 播種與處理
將采集的3個翅果油樹種質的果實分別人工去除木質化的中果皮和胚部的革質種皮,露胚胚部朝下插入6 cm×6 cm的營養缽中(栽培基質:花卉營養土與蛭石1∶3)每缽4粒種子,播種的深度為種子與土壤表面平齊,放置在23~25℃的露天環境,以后每2 d向淺盤中加水1次,保持基質表面濕潤。待幼苗生長15 d之后采用盆栽控水法進行干旱處理。實驗分3個水分脅迫梯度處理,即正常澆水(2 d澆水1次)、輕度干旱脅迫(5 d澆水1次)和重度干旱脅迫(7 d澆水1次),干旱處理2周,每個處理設3組重復,共54缽。取翅果油樹自下而上第2片葉子,作為供試材料進行生理指標的測定。
1.3 相關指標測定
每個種質隨機選取30個果實在電子天平上稱其重量(精度為0.001 g),重復3次;用游標卡尺對3個種質的果實進行長度、寬度測量。
幼苗葉片保水力和相對電導率的測定參照馬引利[6]的方法;葉綠素含量測定采用張志良等[7]的分光光度法;SOD的測定采用氮藍四唑(NBT)法[8];POD 酶活性的測定根據 Jasdanwala 等[9]方法;脯氨酸含量的測定采用磺基水楊酸法[10]。
1.4 數據處理
實驗數據用SPSS13.0軟件進行統計分析。
2 結果與分析
2.1 不同種質果實、幼苗表型的觀察比較
2.1.1 不同種質翅果油樹果實形態觀察
根據帶有中果皮果實的大小、形狀、果皮上溝和脊的花紋顏色等的表型觀察,將本實驗選取的3個供試種質表型特征(圖1)描述如下。
種質1:小宮燈型,果實顆粒小,呈燈籠形,溝和脊的分界明顯,溝的顏色比脊的顏色淺,溝的顏色呈灰白色,脊的顏色呈灰色。
種質2:大宮燈型,果實顆粒較大,呈燈籠形,溝和脊的分界明顯,脊的顏色比溝的顏色要淺,溝的顏色呈深棕色,脊的顏色呈紅棕色。
種質3:長果型,果實的顆粒大,呈梭形,溝和脊的分界明顯,脊的顏色比溝的顏色深,脊的顏色呈棕色,溝的顏色呈灰白色。

圖1 3個翅果油樹種質的果實形態
2.1.2 不同種質翅果油樹果實大小、質量的比較
種子的重量是衡量種子活力的重要指標之一,粒大、飽滿的種子、內部貯藏的營養物質多,有利于種子萌發和幼苗的生長。由圖1和表1可以看出,3個種質在質量、長度和寬度上有極顯著差異。種質3的質量和長度要明顯大于種質1和2,種質2的寬度要大于種質1和3。
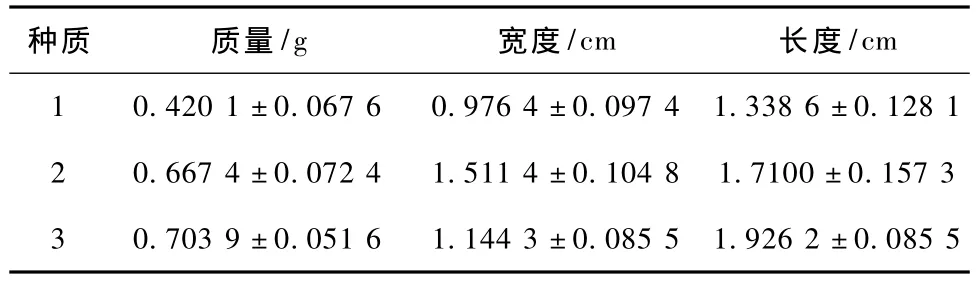
表1 3個種質果實大小、質量的比較
2.1.3 不同資源種質間幼苗的生長發育和表型差異
幼苗生長的第18天,種質1的葉片沒有完全展開,為淡綠色,株高為0.7~1.1 cm;種質2的葉片狹長,呈現黃綠色,株高為1.3~1.8 cm;種質3的葉片肥厚,深綠色,株高為2.0~2.2 cm。幼苗生長的第26天,種質3的生長情況明顯要比種質1和2良好(圖2)。
觀察3個種質表型有明顯差異,3個種質的葉片面積不斷增大,株高增加,測量各種質株高分別為2.2~2.8 cm,2.3~3.0 cm和3.2~3.5 cm。在幼苗生長的第29天,觀察到3個資源種質的株高明顯增加,葉片面積明顯增大,種質1葉片呈現長橢圓形,種質2葉片為卵圓形,種質3葉片呈現長卵圓形。種質3的葉面積明顯大于種質1和種質2,幼苗葉片表型差異更加明顯。測量各種質的株高分別為 3.0~3.3 cm,3.2~3.5 cm和 3.7~4.2 cm。

圖2 翅果油樹3個種質26 d苗齡的幼苗表型
2.2 不同翅果油樹種質幼苗抗旱性比較
2.2.1 干旱脅迫對離體葉片保水力的影響
葉片保水力指葉片在離體條件下,保持原有水分的能力。保水力的高低與植物遺傳特性有關,與細胞特性,特別是與原生質膠體特性有關,反應干旱條件下葉片抗脫水的能力,可以作為衡量植物抗旱品種篩選的簡易指標[6,11]。本實驗測定了輕度脅迫下離體葉片的保水力,發現不同種質葉片間葉片保水力差異極顯著,種質3的葉片保水力高于種質1和2為86%,表明種質3有較強的抗脫水能力,種質2次之,種質1最差(圖 3)。
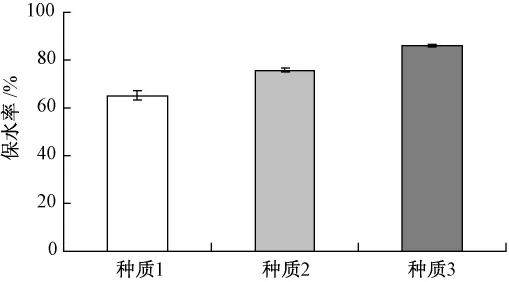
圖3 干旱脅迫下葉片保水力的比較
2.2.2 干旱脅迫對葉片葉綠素含量的的影響
15日苗齡的3個翅果油樹種質分別進行正常澆水、輕度干旱和重度干旱處理15 d后,各取同一葉位的葉片進行葉綠素含量的測定(圖4)。
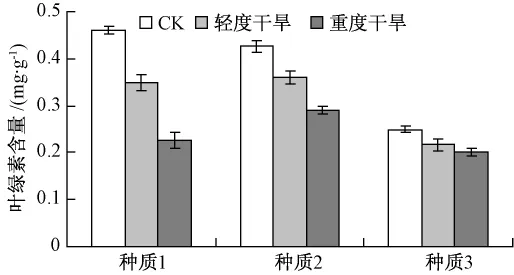
圖4 干旱脅迫下葉綠素含量的變化
圖4 結果表明,隨著干旱程度的加重,各種質葉綠素含量均呈現下降的趨勢,但不同種質間下降的幅度各有差異。種質1下降的幅度最大,種質2次之,種質3下降的幅度最小。說明3個翅果油樹種質的抗旱性強弱為種質3>種質2>種質1。
2.2.3 干旱脅迫對葉片質膜透性的影響
由圖5可知,隨著干旱程度的加重,3個供試種質的相對電導率均呈現不同程度的上升,在重度干旱脅迫處理時,種質1,2和3的相對電導率比對照分別增加了56.63%,43.12%,27.39%。因此在干旱脅迫下,種質3的相對電導率變化較小,說明干旱脅迫對種質3的傷害最小,其抗旱能力要大于種質1和種質2。
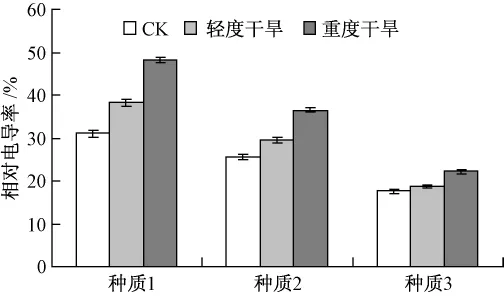
圖5 干旱脅迫對細胞膜透性的影響
2.2.4 干旱脅迫對葉片保護酶活性的影響
由圖6和圖7可知,在干旱脅迫條件下,3個種質的SOD、POD活性均呈上升趨勢,但上升的幅度有所差異。種質3的 SOD、POD活性迅速升高,且含量明顯高于種質1和種質2。由此可知,種質3具有明顯的清除體內活性氧的能力,而種質1和種質2抵御活性氧的能力較弱。這說明在干旱脅迫下,種質3有強的自我修復能力和生理調節能力,對逆境的抗旱能力較強。

圖6 干旱脅迫下POD活性的比較
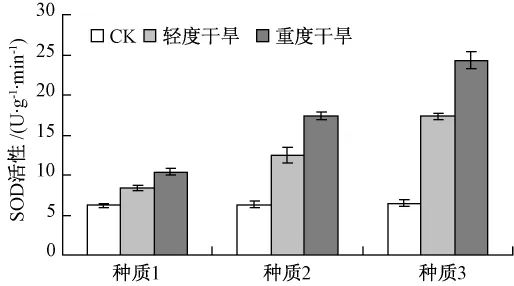
圖7 干旱脅迫下SOD活性的比較
2.2.5 干旱脅迫對葉片脯氨酸含量的影響
從圖8可知,在干旱脅迫下,各種質葉片脯氨酸含量均有不同程度的增長,不同種質間差異極顯著,與POD和SOD酶活性變化趨勢有一定的相似性。在重度干旱處理時,種質1,2和3的脯氨酸含量比對照分別增加了0.527,1.277和2.201倍。種質3葉片內脯氨酸含量增幅最大,則說明種質3在干旱脅迫下滲透調節能力最強,抗旱能力也最強。
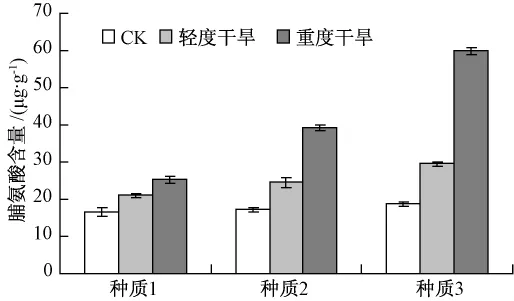
圖8 干旱脅迫下脯氨酸含量的變化
3 小結和討論
3.1 不同翅果油樹種質之間果實與幼苗之間的表型差異
選取鄉寧縣3個翅果油樹種質為材料,研究不同翅果油樹種質資源品種之間的果實、幼苗的表型差異和形態的多樣性及不同種質間的抗旱生理特性差異,結果表明,3個種質在果型、堅果花紋、干果的大小、重量以及幼苗生長發育表型等方面有很大差異,而且各種質幼苗葉片面積大小與果實重量大小之間存在相關性,果型與幼苗葉形之間也有一定的相關性,與陳惠等[5]用翼城產地的2個翅果油樹種質為材料研究的結果是一致的。因此,中果皮花紋、顏色、干果大小等可作為翅果油樹品種鑒定的可靠指標。
3.2 不同翅果油樹種質之間的抗旱生理特性差異
水分脅迫是各種環境脅迫中最為普遍的逆境因子之一。植物對干旱脅迫的響應包含著形態結構和一系列復雜的生理生化變化,并形成了植物各自遺傳性機制的適應能力[12-13]。為了研究不同翅果油樹種質對干旱脅迫的抵抗能力,本研究對3個種質進行了正常澆水、輕度干旱脅迫和重度干旱脅迫3種梯度處理,通過測定其離體葉片的保水力、葉綠素含量、質膜透性、POD、SOD保護酶活性和脯氨酸含量,對其抗旱性的差異進行了比較。
任慶成等研究顯示:在相同的水分脅迫條件下,抗旱性強的烤煙品種葉片相對含水量下降速度較慢且葉片保水能力強[11]。本研究采用的3個翅果油樹種質表現出相似的規律,種質3的葉片保水能力最強,抗旱能力最高,可以有效緩解干旱對植物細胞結構的破壞,維持較長時間的正常生命活動。
干旱脅迫引起植物光合能力下降,可能由于干旱使植物葉綠體中的光合作用片層結構受損,光合磷酸化和電子傳遞受阻[14]。在重度脅迫下,不同種質的翅果油樹葉片葉綠素含量均有下降,但是幼苗仍可以維持正常的生命活動,這說明翅果油樹具有一定的抗旱性,與劉仁濤等對翅果油樹抗旱性研究的結果一致[15]。但3個種質之間存在差異,種質3葉綠素含量下降的幅度最小最抗旱,種質2次之。
保護酶系統的POD、SOD、CAT是植物細胞抵御清除活性氧,降低膜脂過氧化水平重要的保護系統[16]。在干旱脅迫下,各種質翅果油樹葉片的SOD和POD酶活性均有增高,表明其自身的保護能力加強,翅果油樹具有一定的抗旱潛能,也說明在本研究設計的干旱脅迫梯度下,并未對翅果油樹幼苗產生嚴重傷害。但各種質保護酶增加的程度不同,種質3增加最多,因此抵御干旱的能力最強最為抗旱,種質2次之,種質1最弱。脯氨酸是一種滲透調節物質,可以降低因滲透失水造成的對細胞膜、酶及蛋白質結構與功能的傷害,在提高植物對各種脅迫因子抗性等方面起著十分重要的作用[17]。在干旱脅迫下,種質3幼苗葉片內的脯氨酸含量迅速增加且增加幅度最大,因此干旱適應性最強。
綜上所述,根據多種生理指標的綜合分析可以確定3個翅果油樹種質的抗旱性順序依次為:種質3、種質2、種質1。本研究再次證明翅果油樹在形態、生理生化特性等方面具有豐富的遺傳多樣性,為新優品種的選育,優良種質資源的發現等方面提供了一定的方法,具有一定的參考價值。
[1]宋朝樞,徐榮章,張清華.中國珍稀瀕危保護植物[M].北京:中國林業出版社,1989.
[2]賈良智,周俊.中國油脂植物[M].北京:科學出版社,1987.
[3]馮寶英,楊坪榮.翅果油樹種仁化學成分分析研究[J].山西林業科技,1989(4):4-9.
[4]王毅巖.一種新的共生固氮植物:翅果油樹[J].植物學報,1981,23(5):389-391.
[5]陳惠,盧英梅,申峻沛.不同栽培基質對翅果油樹種子萌發的影響及不同種質幼苗生長發育的表型研究[J].山西師范大學學報:自然科學版,2010,24(3):63-66.
[6]馬引利,楊純,閆桂琴,等.土壤水分對翅果油樹幼苗生理生化特性的影響[J].西北植物學報,2008,28(7):1397-1403.
[7]張志良.植物生理學實驗指導[M].北京:高等教育出版社,2003.
[8]GiannopolitisC N,RiesS K.Superoxide dismutase I:Occurrence in higher plants[J].Plant Physiolgy,1977,59:309-314.
[9]Jasdanwala R T,Singh Y D,Chiony J J.Auxin metabolism in developing cotton hairs[J].Journal of Experimental Botany,1997,28:1111-1116.
[10]李霞,閻秀峰,于濤.水分脅迫對黃檗幼苗保護酶活性及脂質過氧化作用的影響[J].應用生態學報,2005,16(12):2353-2356.
[11]任慶成,陳秀華,張生杰,等.不同烤煙品種抗旱生理特征比較研究[J].西北植物學報,2009,29(10):2019-2025.
[12]Media BV.Regulation of gene expression during water deficit stress[J].Plant Growth Regulation,1999,29:22-33.
[13]Egert M,Tevini M.Influence of drought on some physiological parameters symptomatic for oxidative stress in leaves of chives(Allium schoenoprasum)[J].Envrionmental and Experimental Botany,2002,48(1):43-49.
[14]蔣龍,尹俊,孫振中.4種畫眉草抗旱性比較[J].草業科學,2009,11(26):66-70.
[15]劉任濤,畢潤成,任佳.翅果油樹幼苗抗旱性[J].生態學雜志,2006,25(12):1528-1531.
[16]Scandalios J G.Oxygen stress and superoxide dismutase[J].Plant Physiol,1993,101(1):7 - 12.
[17]胡化廣,劉建秀,周志芳,等.結縷草屬植物抗旱生理機理[J].草地學報,2008,16(2):143-144.
