瀕危樹種極東錦雞兒(Caragana fruticosa)種子的萌發(fā)特性
2012-08-02 00:13:04翟曉杰沈海龍
森林工程 2012年5期
關(guān)鍵詞:影響
楊 玲,翟曉杰,沈海龍
(東北林業(yè)大學(xué)林木遺傳育種國(guó)家重點(diǎn)實(shí)驗(yàn)室,哈爾濱150040)
錦雞兒 (Caragana spp.)根系強(qiáng)大,耐沙埋,耐寒,耐干旱,在防風(fēng)固沙、退耕還草、促進(jìn)生態(tài)平衡方面具有重要作用[1-2]。同時(shí),錦雞兒具有飼用、綠肥、蜜源、藥用、木質(zhì)纖維等豐富的資源價(jià)值[3]。極東錦雞兒 [Caragana fruticosa(Pall.)Besser.]是豆科 (Leguminosae)錦雞兒屬 (Caragana)多年生落葉小灌木[4],在我國(guó)主要分布于黑龍江省尚志縣帽兒山山頂,是黑龍江省特有種[5]。極東錦雞兒數(shù)量稀少,呈不規(guī)則小塊狀分布于帽兒山山頂約200 m2平臺(tái)四周及山頂南部坡度緩巖石裸露處,在山頂峭壁下也偶可觀察到單株分布,分布范圍狹小,已處于滅絕邊緣[5]。因此,建立極東錦雞兒苗木的擴(kuò)繁方法以保護(hù)其種質(zhì)資源、促進(jìn)和改善其繁育和苗木培育現(xiàn)狀成為亟待解決的問(wèn)題。
錦雞兒可以通過(guò)種子[6-10]、扦插[11]、根孽[12]和組織培養(yǎng)[13-16]來(lái)繁殖。最容易、最常用的繁殖方法仍是通過(guò)種子進(jìn)行繁殖。人們?cè)阱\雞兒種子萌發(fā)和苗木培育方法上積累了一定的經(jīng)驗(yàn)。如對(duì)白皮錦雞兒(C.leucophloea)、草原錦雞兒(C.pumila)和刺葉錦雞兒(C.acanthophylla)種子啟動(dòng)萌發(fā)所需最低土壤水分含量的研究[10],對(duì)小葉錦雞兒(C.microphylia)種子大小變異對(duì)種子萌發(fā)率和幼苗生長(zhǎng)影響的研究[9],對(duì)小葉錦雞兒1a生播種苗生長(zhǎng)規(guī)律的研究[6]以及對(duì)中間錦雞兒(C.intermedia)種子成熟后采摘時(shí)期和種子產(chǎn)量的研究[17]等。目前,尚未見到關(guān)于極東錦雞兒種子萌發(fā)和出苗影響的系統(tǒng)報(bào)道。本文以分布于黑龍江省帽兒山頂?shù)奶烊粯O東錦雞兒為研究對(duì)象,研究其種子物理性狀和不同溫度條件對(duì)其種子萌發(fā)和幼苗生長(zhǎng)的影響,旨在為建立極東錦雞兒播種育苗技術(shù)和確定合理播種期提供科學(xué)依據(jù),為改善極東錦雞兒苗木培育狀況、實(shí)現(xiàn)極東錦雞兒種質(zhì)資源保存和合理開發(fā)利用奠定基礎(chǔ)。
1 材料與方法
1.1 材料采集與處理
種子取自黑龍江省尚志市帽兒山山頂極東錦雞兒天然居群。2010年8月中旬采集成熟種莢,于室內(nèi)通風(fēng)處陰干,待種莢裂開后去雜質(zhì),收集種子,于4℃下密封儲(chǔ)存。
1.2 種莢結(jié)實(shí)率和種子形態(tài)特征調(diào)查
種子顏色、形狀通過(guò)目測(cè)觀察進(jìn)行描述;種子大小用游標(biāo)卡尺抽樣測(cè)量,取30粒種子測(cè)量種子的長(zhǎng)、寬、厚。選取500枚種莢統(tǒng)計(jì)種莢內(nèi)種子數(shù)和有蟲的種子數(shù),進(jìn)而計(jì)算出種莢結(jié)實(shí)率和種子蟲害率。

1.3 種子含水量和千粒重的測(cè)定
種子含水量用105℃恒重法 (標(biāo)準(zhǔn)法)測(cè)定,重復(fù)測(cè)定3次,每次50粒種子。計(jì)算公式為:

種子千粒重用百粒法測(cè)定,重復(fù)測(cè)定3次。從純凈種子中,隨機(jī)選取100粒種子為1組,共取8組,即為8個(gè)重復(fù),計(jì)算平均值、標(biāo)準(zhǔn)差及變異系數(shù),得出種子千粒重,單位為克。
1.4 種子生活力和吸水量測(cè)定
種子生活力測(cè)定用TTC染色法進(jìn)行,設(shè)3次重復(fù),每次50粒種子。種子于25℃下在蒸餾水中浸泡24 h,用醫(yī)用解剖刀沿種子的縱軸切開,在25℃下0.1%TTC溶液中染色24 h。以70%面積被染成紅色的種子為有活力的種子,計(jì)算有活力種子占總試驗(yàn)種子數(shù)的百分比。計(jì)算公式為:

種子吸水量測(cè)定采用羅靖德等 (2010)方法[18],選取100粒種子用蒸餾水浸沒(méi),重復(fù)3次,放在25℃恒溫箱中保溫。每隔1 h取出種子,吸干種子表面浮水稱重,直至恒重。計(jì)算種子吸水量。計(jì)算公式為:

1.5 種子萌發(fā)測(cè)驗(yàn)
種子在室溫下自來(lái)水中沖泡3 d。選取籽粒飽滿、質(zhì)地均勻的種子,用0.3%KMnO4溶液消毒15 min,后用清水沖洗干凈。每30粒種子置于鋪有2層濕潤(rùn)濾紙的培養(yǎng)皿內(nèi),在光照、不同溫度條件下進(jìn)行種子萌發(fā)試驗(yàn),每處理4個(gè)重復(fù)。溫度設(shè)置為15℃、20℃、25℃和30℃4種溫度梯度。每天12 h光照 (光照強(qiáng)度為100μmol·m-2·s-1),12 h黑暗。保持培養(yǎng)皿內(nèi)濕潤(rùn)和通氣。每天觀測(cè)記錄種子的發(fā)芽情況,當(dāng)胚根長(zhǎng)度達(dá)到與種子等長(zhǎng)、胚芽長(zhǎng)度達(dá)到種子一半時(shí),作為種子萌發(fā)標(biāo)準(zhǔn)。以發(fā)芽率和發(fā)芽勢(shì)為指標(biāo),在發(fā)芽種子數(shù)達(dá)到高峰時(shí)計(jì)算發(fā)芽勢(shì),在發(fā)芽末期連續(xù)5 d發(fā)芽粒數(shù)平均不足供試種子總數(shù)的1%時(shí)計(jì)算發(fā)芽率。發(fā)芽率和發(fā)芽勢(shì)計(jì)算公式如下:

1.6 幼苗生長(zhǎng)測(cè)量
發(fā)芽試驗(yàn)結(jié)束后,分別選取萌發(fā)一致的種子置于鋪有兩張濾紙的柱形玻璃管中 (1.0 cm ×9.5 cm),在氣候箱中培養(yǎng) (溫度為25℃和30℃2個(gè)溫度梯度,光照條件同1.5),定時(shí)補(bǔ)充蒸餾水。每個(gè)處理10株幼苗,重復(fù)3次,10 d后記錄幼苗的苗高、胚根長(zhǎng)和生物量。
1.7 數(shù)據(jù)處理和統(tǒng)計(jì)分析
表格和圖中數(shù)據(jù)均為各重復(fù)處理的平均數(shù)±標(biāo)準(zhǔn)差。百分比數(shù)據(jù)在統(tǒng)計(jì)分析前進(jìn)行了反正弦平方根轉(zhuǎn)換。應(yīng)用Excel 2003軟件進(jìn)行數(shù)據(jù)處理,應(yīng)用SigmaPlot 10.0軟件對(duì)數(shù)據(jù)進(jìn)行作圖,用SPSS17.0軟件對(duì)數(shù)據(jù)進(jìn)行方差分析和多重比較分析。
2 結(jié)果與分析
2.1 種子物理性狀觀測(cè)
極東錦雞兒種子呈腎形,少數(shù)為圓形,顏色有黃色、黃褐色和褐色,種皮革質(zhì),光滑,多數(shù)表面具紅褐色花紋。種子大小及其變異系數(shù)見表1。極東錦雞兒種子長(zhǎng)、寬、厚存在一定變異。其中種子長(zhǎng)/寬的變異系數(shù)最大,其次是種子厚度變異系數(shù),種子長(zhǎng)和寬的變異系數(shù)相近,均稍大于10%。種子結(jié)實(shí)率不高,平均每枚種莢結(jié)1.43粒種子。種子受蟲害嚴(yán)重,種子蟲害率達(dá)25.6%。成熟種子含水量為6.6%,千粒重為18.6 g。TTC染色法測(cè)定種子生活力為94%,說(shuō)明極東錦雞兒種子發(fā)育良好,4℃低溫儲(chǔ)存可以保持種子生活力。
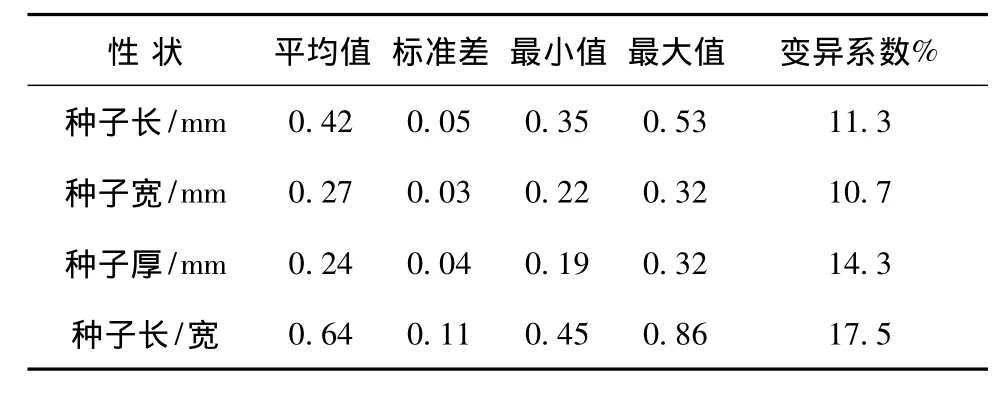
表1 極東錦雞兒種子大小及其變異系數(shù)Tab.1 The size and variation coefficient of Caragana fruticosa seeds
25℃下,極東錦雞兒種子的吸脹速度在1~2 h內(nèi)最快,吸脹4 h后種子的吸水量基本達(dá)到飽和,如圖1所示。種子最終吸水量達(dá)到53%左右。說(shuō)明極東錦雞兒種皮透水性良好,在水分充足時(shí)種子能迅速吸水為萌發(fā)做準(zhǔn)備。方差分析結(jié)果表明,不同吸脹時(shí)間的種子吸水量差異極顯著 (P=0.000 1)。不同吸脹時(shí)間下的種子吸水量多重比較結(jié)果如圖1所示。
2.2 溫度對(duì)極東錦雞兒種子萌發(fā)的影響
2.2.1 溫度對(duì)極東錦雞兒種子發(fā)芽率的影響
極東錦雞兒種子在恒溫15℃、20℃、25℃和30℃條件下的發(fā)芽情況如圖2所示。不同發(fā)芽溫度對(duì)種子發(fā)芽率和發(fā)芽勢(shì)影響的多重比較結(jié)果如圖3所示。不同溫度條件對(duì)種子發(fā)芽率具有極顯著影響(P=0.000 1)。種子在置床后第四天開始萌發(fā),在第十一天以后基本平穩(wěn)。在4種溫度梯度中,25℃條件下種子的發(fā)芽率最高,為38.33%。隨著溫度的升高或降低,種子發(fā)芽率降低。多重比較結(jié)果表明,30℃種子發(fā)芽率 (31.67%)與25℃種子發(fā)芽率相比差異不顯著;20℃條件下種子發(fā)芽率 (3.33%)與30℃、25℃相比差異極顯著;15℃條件下種子發(fā)芽率最低 (0.83%),與20℃相比差異不顯著,但與30℃、25℃相比差異極顯著,如圖3所示。

圖1 25℃下極東錦雞兒種子吸水量曲線圖Fig.1 The seed soakage curve of Caragana fruticosa at 25℃(Values represented the mean and standard error,means within a data point followed by different lowercase letters are highly significantly different from each other at P=0.01 level of significance using the Duncan's Multiple Range Test.)
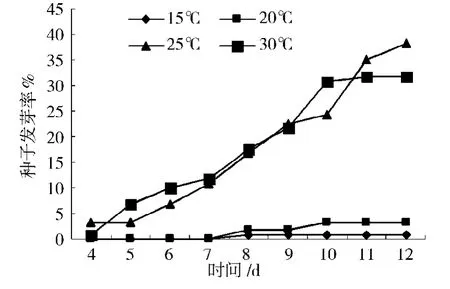
圖2 發(fā)芽溫度對(duì)極東錦雞兒種子萌發(fā)的影響Fig.2 Effects of temperatures on seeds germination of Caragana Fruticosa
2.2.2 溫度對(duì)極東錦雞兒種子發(fā)芽勢(shì)的影響
極東錦雞兒種子在恒溫15℃、20℃、25℃、30℃條件下種子發(fā)芽勢(shì)及其多重比較結(jié)果如圖3所示。不同溫度條件對(duì)種子發(fā)芽勢(shì)具有顯著的影響(P=0.03)。發(fā)芽試驗(yàn)的第八天時(shí),25℃條件下種子發(fā)芽勢(shì)為 16.67%,達(dá)到最大發(fā)芽率的43.49%;30℃條件下種子發(fā)芽勢(shì)為17.50%,為最大發(fā)芽率的55.26%;20℃條件下種子發(fā)芽勢(shì)為1.67%,為最大發(fā)芽率的50.15%;15℃條件下,種子發(fā)芽勢(shì)最低,為0.83%,此后種子發(fā)芽數(shù)量沒(méi)再增加。不同溫度下的種子發(fā)芽勢(shì)均在發(fā)芽試驗(yàn)的第八天達(dá)到50%以上,說(shuō)明發(fā)芽試驗(yàn)的第八天是計(jì)算極東錦雞兒種子發(fā)芽勢(shì)的適宜時(shí)期。多重比較結(jié)果表明,30℃條件下種子發(fā)芽勢(shì)與25℃、20℃的相比差異均不顯著,但與15℃的相比差異顯著。25℃條件下種子發(fā)芽勢(shì)與20℃、15℃相比差異均為顯著,15℃條件下種子發(fā)芽勢(shì)與20℃相比差異不顯著,如圖3所示。

圖3 極東錦雞兒種子發(fā)芽率和發(fā)芽勢(shì)的多重比較Fig.3 Multiple comparison of germination percentage and germination energy of Caragana fruticosa seeds.(Means within a data point of germination energy followed by different lowercase letters are significantly different from each other at P=0.05 level of significance,and the means within a data point of germination percentage followed by different capital letters are highly significantly different from each other at P=0.01 level of significance using the Duncan's Multiple Range Test.)
2.3 溫度對(duì)極東錦雞兒幼苗生長(zhǎng)的影響
2.3.1 溫度對(duì)極東錦雞兒幼苗高度的影響
2種不同溫度 (25℃和30℃)條件下極東錦雞兒幼苗高度及其多重比較結(jié)果如圖4所示。方差分析結(jié)果表明,不同溫度條件對(duì)極東錦雞兒幼苗高度的影響極顯著 (P=0.001)。萌發(fā)后培養(yǎng)10 d時(shí),30℃條件下極東錦雞兒幼苗高度僅為0.98 cm,而25℃條件下幼苗高度為3.4 cm,是30℃條件下的3.5倍。
2.3.2 溫度對(duì)極東錦雞兒幼苗胚根長(zhǎng)的影響
2種不同溫度 (25℃和30℃)條件下極東錦雞兒幼苗胚根長(zhǎng)度及其多重比較結(jié)果如圖5所示。方差分析結(jié)果表明,不同溫度條件對(duì)極東錦雞兒幼苗胚根長(zhǎng)的影響極顯著 (P=0.004)。萌發(fā)后培養(yǎng)10 d時(shí),30℃條件下的幼苗胚根長(zhǎng)僅為1.76 cm,而25℃條件下的幼苗胚根長(zhǎng)為3.1 cm,是30℃條件下的1.8倍。
2.3.3 溫度對(duì)極東錦雞兒幼苗生物量的影響
2種溫度 (25℃和30℃)條件下極東錦雞兒幼苗生物量及其多重比較結(jié)果如圖6所示。方差分析結(jié)果表明,不同溫度條件對(duì)極東錦雞兒幼苗生物量的影響不顯著 (P=0.25)。萌發(fā)后培養(yǎng)10 d時(shí),30℃條件下的幼苗生物量為0.16 g,而25℃條件下的極東錦雞兒幼苗生物量為0.07 g,30℃條件下的幼苗鮮重是25℃條件下的2.3倍。說(shuō)明較高的溫度有利于極東錦雞兒幼苗生物量的增加。
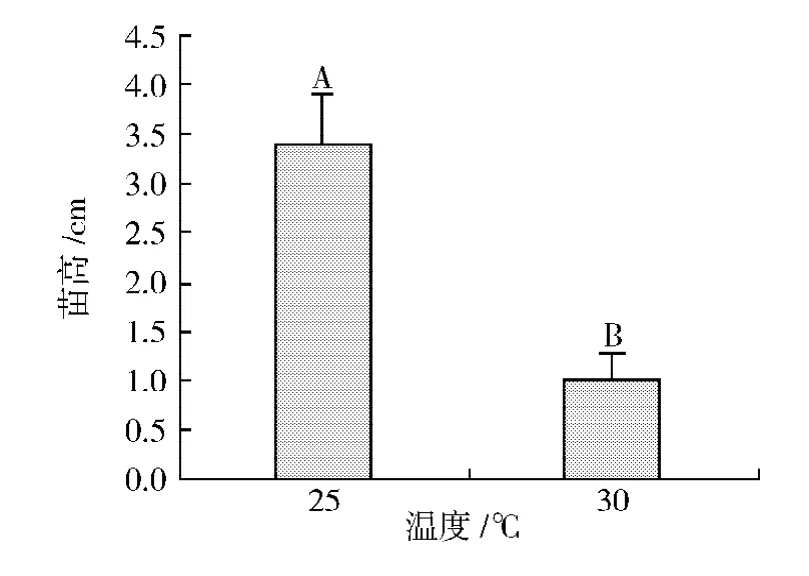
圖4 溫度對(duì)極東錦雞兒幼苗高度的影響Fig.4 Effects of temperatures on height of Caragana fruticosa seedlings.(Means within a data point of seedling height followed by different capital letters are highly significantly different from each other at P=0.01 level of significance using the Duncan's Multiple Range Test.)
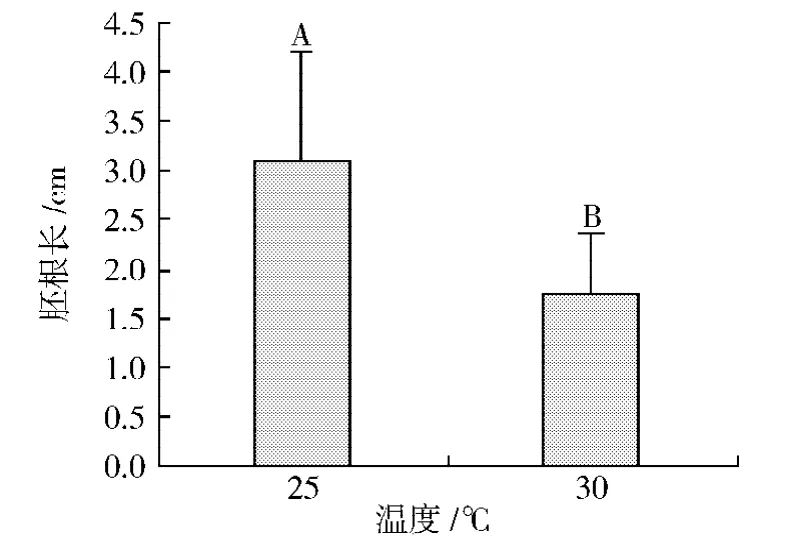
圖5 溫度對(duì)極東錦雞兒幼苗根胚根長(zhǎng)度的影響Fig.5 Effects of temperatures on seedling length of Caragana fruticosa.(Means within a data point of radicle length followed by different capital letters are highly significantly different from each other at P=0.01 level of significance using the Duncan's Multiple Range Test.)
3 討論
危害錦雞兒樹種的害蟲種類很多[19]。多數(shù)害蟲在植株開花時(shí)將卵產(chǎn)在果實(shí)子房?jī)?nèi)或子房上,蟲卵在種子內(nèi)部發(fā)育,并蠶食、危害種子[17]。極東錦雞兒未成熟種莢表面無(wú)明顯的蟲子侵入孔,而在成熟種子表面有羽化孔,推測(cè)是在開花期害蟲產(chǎn)下蟲卵,幼蟲在種子內(nèi)部發(fā)育變成成蟲,羽化后咬破種皮和種莢飛出導(dǎo)致的。極東錦雞兒種子的蟲害率達(dá)到1/4,嚴(yán)重影響了種子產(chǎn)量和實(shí)生苗數(shù)量。
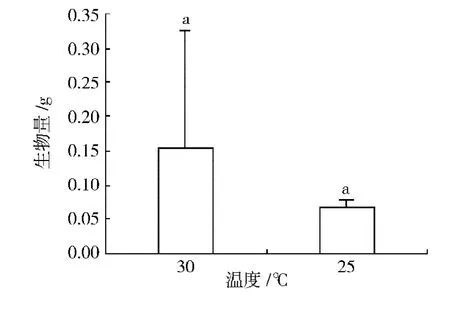
圖6 溫度對(duì)極東錦雞兒幼苗生物量的影響Fig.6 Effects of temperatures on total biomass of Caragana fruticosa seedlings.(Means within a data point of biomass followed by same capital letters are not significantly different from each other at P=0.05 level of significance using the Duncan's Multiple Range Test.)
植物種子大小變異既受物種自身內(nèi)部遺傳基因的控制,又受外在變化的環(huán)境因素的影響[20],同時(shí)種實(shí)在植株上的位置也對(duì)種子大小有一定影響[21]。種子大小對(duì)植物早期生長(zhǎng)發(fā)育影響很大,主要表現(xiàn)在種子萌發(fā)率[22-23]、幼苗出土深度[24-25]、幼苗大小[26-27]和幼苗競(jìng)爭(zhēng)能力[22,28]等方面。極東錦雞兒種子千粒重為18.6 g,屬于小粒種子。種子較小的物種比較大種子物種對(duì)光照的需求更大,但是在受到高溫脅迫時(shí),較小種子物種比較大種子物種具有更大的萌發(fā)能力和抵抗脅迫的能力[29]。極東錦雞兒種子表型存在變異,這種差異對(duì)于極東錦雞兒適應(yīng)環(huán)境繁衍后代具有積極的意義。自然群體中保持大的變異貯存對(duì)群體是有利的,群體內(nèi)多種基因型所對(duì)應(yīng)的表型范圍很多,從而使群體在整體上適應(yīng)可能遇到的大多數(shù)環(huán)境條件[30]。
植物對(duì)生境的適應(yīng)與否主要表現(xiàn)在種子的萌發(fā)速度上[31]。極東錦雞兒種子萌發(fā)從開始到結(jié)束需要10 d時(shí)間。在試驗(yàn)過(guò)程中極東錦雞兒除個(gè)別種子腐爛外,多數(shù)種子在短時(shí)間內(nèi)能萌發(fā),這是植物為物種延續(xù)和進(jìn)一步的地域擴(kuò)張而形成的大部分種子在適宜條件下萌發(fā),以提高幼苗定植幾率的一種生態(tài)適應(yīng)。但在帽兒山山頂?shù)膼毫由姝h(huán)境下,這種生態(tài)適應(yīng)增加了極東錦雞兒實(shí)生幼苗在后續(xù)干旱少雨氣候中死亡的風(fēng)險(xiǎn),這也許是極東錦雞兒在帽兒山山頂群落中少有實(shí)生苗的原因之一。
光照和溫度對(duì)植物種子萌發(fā)具有重要的作用[32-33]。對(duì)于不同的植物來(lái)說(shuō),種子萌發(fā)的最適溫度不同[32-35]。在不同的溫度條件下,極東錦雞兒種子萌發(fā)進(jìn)程差異較大。不同的溫度條件對(duì)最終的發(fā)芽率和發(fā)芽勢(shì)具有顯著的影響。本研究結(jié)果表明,25℃條件更適合極東錦雞兒種子的萌發(fā),其發(fā)芽率和發(fā)芽勢(shì)最高。發(fā)芽勢(shì)可以作為指示種子發(fā)芽整齊度和發(fā)芽速度的指標(biāo),一般以整個(gè)發(fā)芽實(shí)驗(yàn)進(jìn)程的1/3或1/2時(shí)間點(diǎn)作為發(fā)芽勢(shì)的統(tǒng)計(jì)時(shí)期。本研究結(jié)果表明,在發(fā)芽試驗(yàn)的第八天,不同溫度條件下的種子發(fā)芽率基本達(dá)到種子最大發(fā)芽率的50%。說(shuō)明發(fā)芽試驗(yàn)的第八天是計(jì)算極東錦雞兒種子發(fā)芽勢(shì)的適宜時(shí)期。
溫度影響種子萌發(fā)后的幼苗的生長(zhǎng)發(fā)育[35-36]。本研究結(jié)果表明,不同溫度條件對(duì)幼苗高度和胚根長(zhǎng)度影響差異極顯著,但對(duì)總生物量的積累影響不顯著。25℃時(shí)極東錦雞兒的幼苗高度和胚根生長(zhǎng)量分別與30℃時(shí)的差異達(dá)到極顯著水平,但生物量的積累與30℃相比無(wú)顯著差異。因此,在直播培育極東錦雞兒苗木時(shí),除了需要有適當(dāng)?shù)墓庹蘸蜐穸葪l件,建議首選25℃溫度條件進(jìn)行播種育苗。
[1]Cao CY,Jiang D M,Teng X H,et al.Soil chemical and microbiological properties along a chronosequence of Caragana microphylla Lam.plantations in the Horqin sandy land of Northeast China[J].Applied Soil Ecology,2008,40(1):78-85.
[2]秦樹高,吳 斌,張宇清,等.錦雞兒林草帶狀復(fù)合系統(tǒng)細(xì)根空間分異特征[J].林業(yè)科學(xué),2011,47(7):42-49.
[3]賈 麗,曲式曾.豆科錦雞兒屬植物研究進(jìn)展[J].植物研究,2001,21(4):515-518.
[4]周以良.黑龍江省樹木志[M].哈爾濱:東北林業(yè)大學(xué)出版社,1986.
[5]秦瑞明,王 迪,遲福昌.黑龍江省稀有瀕危植物[M].哈爾濱:東北林業(yè)大學(xué)出版社,1993.
[6]孫志蓉,翟明普,王文全,等.小葉錦雞兒1年生播種苗的生長(zhǎng)規(guī)律[J].林業(yè)科技,2004,29(5):5-8.
[7]趙庭花.甘蒙錦雞兒容器育苗和苗期高生長(zhǎng)觀測(cè)[J].青海農(nóng)林科技,2006,(4):71.
[8]冷肖荀.甘蒙錦雞兒容器苗培育基質(zhì)研究[J].福建林業(yè)科技,2007,34(2):137-139.
[9]何玉惠,趙哈林,劉新平,等.小葉錦雞兒種子大小變異對(duì)萌發(fā)和幼苗生長(zhǎng)的影響[J].種子,2008,27(8):10-13.
[10]李曉梅,趙曉英,趙 法,等.新疆3種錦雞兒屬植物種子萌發(fā)對(duì)溫度和土壤水分的響應(yīng)[J].草業(yè)科學(xué),2009,26(10):140-145.
[11]季 蒙,楊文斌,梁海榮,等.中間錦雞兒嫩枝扦插初步研究[J].內(nèi)蒙古林業(yè)科技,2006,(1):5-8.
[12]張建華,馬成倉(cāng),劉志宏,等.干旱荒漠區(qū)狹葉錦雞兒灌叢擴(kuò)展對(duì)策[J].生態(tài)學(xué)報(bào),2011,31(8):2132-2138.
[13]胡鈉梅,韓素英,梁國(guó)魯,等.TDZ誘導(dǎo)中間錦雞兒植株再生[J].北方園藝,2009,(8):204-205.
[14]李光道,白生才,張秀志,等.檸條錦雞兒組培研究[J].甘肅農(nóng)業(yè)科技,2010,(11):18-20.
[15]Zhai X J,Yang L,Shen H.Shoot multiplication and plant regeneration in Caragana fruticosa(Pall.)Besser[J].Journal of Forestry Research,2011,22(4):561-567.
[16]沈海龍,翟曉杰,楊 玲.極東錦雞兒幼胚子葉體細(xì)胞胚發(fā)生和植株再生[J].植物生理學(xué)報(bào),2011,47(4):399-405.
[17]陳河龍,王 贊,孫桂枝,等.中間錦雞兒結(jié)實(shí)習(xí)性[J].草業(yè)科學(xué),2007,24(2):20-25.
[18]羅靖德,甘小洪,賈曉娟,等.瀕危植物水青樹種子的生物學(xué)特性[J].云南植物研究,2010,32(3):204-210.
[19]楊彩霞,高立原.寧夏固沙植物檸條昆蟲資源的調(diào)查[J].中國(guó)沙漠,2000,20(4):61-63.
[20]張世挺,杜國(guó)禎,陳家寬.種子大小變異的進(jìn)化生態(tài)學(xué)研究現(xiàn)狀與展望[J].生態(tài)學(xué)報(bào),2003,23(2):353-364.
[21]Cavers P B,Harper J L.Germination polymorphism in Rumex xrispus and Rumex obtusifoltus[J].The Journal of Ecology,1996,54(2):367-382.
[22]Gross K L.Effects of seed size and growth form on seedling establishment of six monocarpic perennials[J].The Journal of Ecology,1984,72(2):369-376.
[23]Greipsson SA,Davy J.Seed mass and germination behavior in populations of the dune-building grass Leymus arenarius[J].Annals of Botany,1995,76:493-501.
[24]Reader R J,Best B J.Variation in competition along an environmental gradient:Hieracium floribundumin an abandoned pasture[J].The Journal of Ecology,1989,77(3):673-684.
[25]Gòmez JM.Bigger is not always better:Conflicting selective pressures on seed size in Quercus ilex[J].Evolution,2004,58(1):71-80.
[26]Marshall D.Effects of seed size on seedling success in three species of Sesbania(Fabaceae)[J].America Journal of Botany,1986,73(4):457-464.
[27]Seiwa K.Effects of seed size and emergence time on tree seedling establishment:Importance of developmental constraints[J].Oecologia,2000,123:208-215.
[28]Lloret F,Casanovas C,Pe?uelas J.Seedling survival of Mediterranean shrubland species in relation to root:shoot ratio,seed size and water and nitrogen use[J].Functional Ecology,1999,13:210-216.
[29]武高林,杜國(guó)禎.植物種子大小與幼苗生長(zhǎng)策略研究進(jìn)展[J].應(yīng)用生態(tài)學(xué)報(bào),2008,19(1):191-197.
[30]李文英,顧萬(wàn)春.蒙古櫟天然群體表型多樣性研究[J].林業(yè)科學(xué),2005,41(1):49-56.
[31]Proberts R J.The role of temperature in germination ecphysiology[A].In:Fenner M Ed.Seeds:The ecology of regeneration in plant communities[C].Wallingford:C.A.B.International,2000,285-325.
[32]吳 彥,劉 慶,何 海,等.光照與溫度對(duì)云杉和紅樺種子萌發(fā)的影響[J].應(yīng)用生態(tài)學(xué)報(bào),2004,15(12):2229-2232.
[33]張光飛,王定康,翟書華,等.光照和溫度對(duì)青陽(yáng)參種子萌發(fā)的影響[J].種子,2008,27(2):80-81.
[34]Taylor C W,Gerrie W A.Effects of temperature on seed germination and seed dormancy in Sorbus glabrescens Cardot[J].Acta Horticulture,1987,215:185 ~192.
[35]李有志,黃繼山,朱杰輝.光照和溫度對(duì)小葉章種子萌發(fā)及其幼苗生長(zhǎng)的影響[J].湖南農(nóng)業(yè)大學(xué)學(xué)報(bào)(自然科學(xué)版),2007,33(2):187-190.
[36]楊立學(xué).俄羅斯大果沙棘(Hippophae rhamnoides L.)種子萌發(fā)特性[J].生態(tài)學(xué)報(bào),2007,27(6):2215-2222.
[37]王 峰,王志英,李丹雷,等.小興安嶺代表性菌物系統(tǒng)進(jìn)化初步研究[J].森林工程,2012,28(2):1-3.
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級(jí)版)(2011年3期)2011-01-01 00:00:00
