暗紫貝母的物候和鱗莖在海拔梯度上的變化
2012-08-20 04:07:12陳文年陳發軍謝玉華王淯段輝國齊澤民
草業學報 2012年5期
陳文年,陳發軍,謝玉華,王淯,段輝國,齊澤民
(1.內江師范學院生命科學系,四川 內江641112;2.四川省特色農業資源研究與利用重點實驗室,四川 內江641112)
暗紫貝母(Fritillaria unibracteata)是典型的高山植物,主要生長于青藏高原及其東部的高山地區,海拔一般為3 200~4 500m[1],分布范圍十分有限[2,3]。其藥用部分為干燥鱗莖,是中藥川貝的主要來源之一,屬于名貴中藥材。長期以來,由于藥材市場需求,暗紫貝母被大量采挖,導致其野生資源存量大幅下降,并逐漸趨于枯竭[4],從而又大大影響了藥材的市場供應[5]。因此,如何提高暗紫貝母的產量、滿足市場的需求就成了一個急待解決的問題。以前的研究主要集中于暗紫貝母的組織培養技術和鱗莖化學成分分析方面[6-9],以及暗紫貝母藥材的真偽鑒定方面[10-14]。在生態學領域,主要限于暗紫貝母群落生態學的研究,如暗紫貝母所在的群落類型與其品質之間關系[15],暗紫貝母群落的植物種類組成與區系分布[16]。而對暗紫貝母野生資源的生物學特性及不同生境條件下個體生長特性的比較研究報道很少[17,18]。筆者以生態因子中的海拔梯度作為研究對象,研究其與暗紫貝母鱗莖生長及物候的關系,旨在揭示高山地區海拔對暗紫貝母生長的影響,為暗紫貝母的引種栽培和生產提供了一定的參考,也為其野生資源的保護奠定基礎。
1 材料與方法
1.1 研究地概況
研究地位于青藏高原東部的四川省松潘縣章臘鄉卡卡山(32°59′N,103°40′E)[19,20]。該地區年均溫為2.8℃,1月均溫 -7.6℃,7月均溫9.7℃,年降水量為634.8mm,其中的72%發生在6-8月的夏季,土壤類型為黑氈土[21]。在卡卡山山腳,海拔為3 440m,山頂海拔為3 980m,垂直高度差近540m。
1.2 樣方設置及試驗設計
在卡卡山南坡,取3個海拔部位(3 500,3 750,3 950m),每個部位沿水平方向設置A、B兩個10m×10m調查樣方(水平距離為15m)。在每個樣方中隨機選取30株3年生的暗紫貝母個體(能抽莖但要等到下一年才開花)作為研究對象,以便于對比研究。所有選擇的植株都作了標記,并于2007年5月開始在每個樣方周圍設置鐵絲圍欄以防人畜干擾。
為了確定海拔對暗紫貝母的物候和鱗莖是否有確切的影響,在高、低海拔部位的B樣方之間進行植株的對換移植試驗。把高海拔B樣方的30株植株移植到低海拔部位的B樣方中;同樣,把低海拔部位B樣方中的30株植株移植到高海拔部位的B樣方中。采用連土移植的方法,即每株暗紫貝母連同其植株下面的土塊一起移植,土塊的長、寬、深大小尺寸為0.25m×0.25m×0.30m。整個移植試驗于2007年9月上旬進行,所有的移植植株都作了標記,A樣方作為對照。這樣,在高、低海拔部位就產生4種植株類型:高海拔部位A樣方的對照植株(HA)、高海拔部位B樣方的移植植株(HB)、低海拔部位A樣方的對照植株(LA)、低海拔部位B樣方移植植株(LB)。另外,中間海拔部位(3 750m)還有2個樣方,即中間部位A樣方植株(MA)和中間部位B樣方植株(MB)。
1.3 測定指標
1.3.1 各部位生態因子的測量 在2008,2009年的3—5月,測量各樣方中各選定植株從積雪中融化出來的時間(以1月1日開始的序日表示);同時在高、中、低各海拔部位,分別放置5個自動溫度記錄儀(型號:ZDR-01)以測定3—5月的氣溫變化,記錄儀離地高度為50cm,設定為每小時自動記錄1次;在5月中旬(此時積雪已融化完和凍土已經解凍)到8月底,用便攜式水分測量儀(型號:Mpkit-B)和pH計(型號:PHT-810)測量每個樣方中的土壤含水量及土壤酸堿度,每個樣方取中心和四周測量點5個,每周1次。
1.3.2 物候的觀察 于2008,2009年的3月下旬—9月上旬進行物候觀察,每3天觀察1次。物候期包括:1)開始生長期:以萌芽冒出地面1cm為標準,記錄每個萌芽達到1cm的日期,并轉化為從1月1日開始計算的序日[22](下同);2)始花期:每個樣方的30個植株中有1個開始開花為標準;3)盛花期:以每個樣方30個植株中有一半開始開花為標準;4)植株枯黃期:以每個個體葉片枯黃一半時的日期為標準。
1.3.3 鱗莖重量的測定 2009年9月下旬,將每個調查樣方中標記的30株暗紫貝母植株進行采挖,挖出鱗莖,洗凈,分樣方裝紙袋,帶回實驗室在65℃下烘48h,用精密電子天平(型號:PB303-E)稱量每粒鱗莖的干重。
1.4 統計方法
用SPSS 10.0對不同海拔部位間物候及鱗莖的差別進行方差分析(ANOVA)和多重比較。
2 結果與分析
2.1 不同海拔部位之間生態因子的比較
3個部位的融雪時間有明顯的差別,低海拔部位的積雪融化最早,在3月中下旬即可全部融化完,而高海拔部位則要在4月中下旬才可全部融化,高低部位之間相差可達30d左右(表1)。土壤含水量在3個部位之間差異顯著,高海拔部位比低海拔部位高43.5%。3個部位的pH相差不大,都呈現出弱酸性(表1)。另外從3—5月份的氣溫來看,高、中、低3個部位之間差異較顯著(表2),每個月都是以低海拔部位平均氣溫最高,以高海拔部位的平均氣溫最低,高、低部位之間的平均氣溫差異分別達到了2.7℃(3月)、3.0℃(4月)、5.3℃(5月)。

表1 3個部位環境因子的比較Table 1 Comparison of environmental factors among three portions

表2 3個部位3—5月的平均氣溫比較Table 2 Comparison of average air temperature at three portions in March,April and May ℃
2.2 不同海拔部位間物候的比較
各物候期都是在低海拔部位最先出現,中海拔部位次之,在高海拔部位最遲出現(表3)。就每一物候期來說,在各部位之間的差異都達到了顯著的程度,但同一海拔部位的2個樣方(A、B)之間的差異并不顯著。低海拔部位的暗紫貝母植株移植到高海拔部位之后,其物候表現出高海拔部位對照植株的特點;而高海拔部位的植株移植到低海拔部位之后也同樣表現出低海拔部位對照植株物候的特點。從不同的物候期來看,開始生長期在高、低海拔部位間的物候差異最大,達到25d左右;始花期和盛花期次之;而枯黃期在高、低海拔部位間的物候差異最小,只有14d。隨著植株的生長,越靠前的物候階段在高、低海拔部位之間的差異越大;而越靠后的則相反。各物候期在高、低海拔部位之間的差異可以從移植實驗的結果得到驗證(表3)。
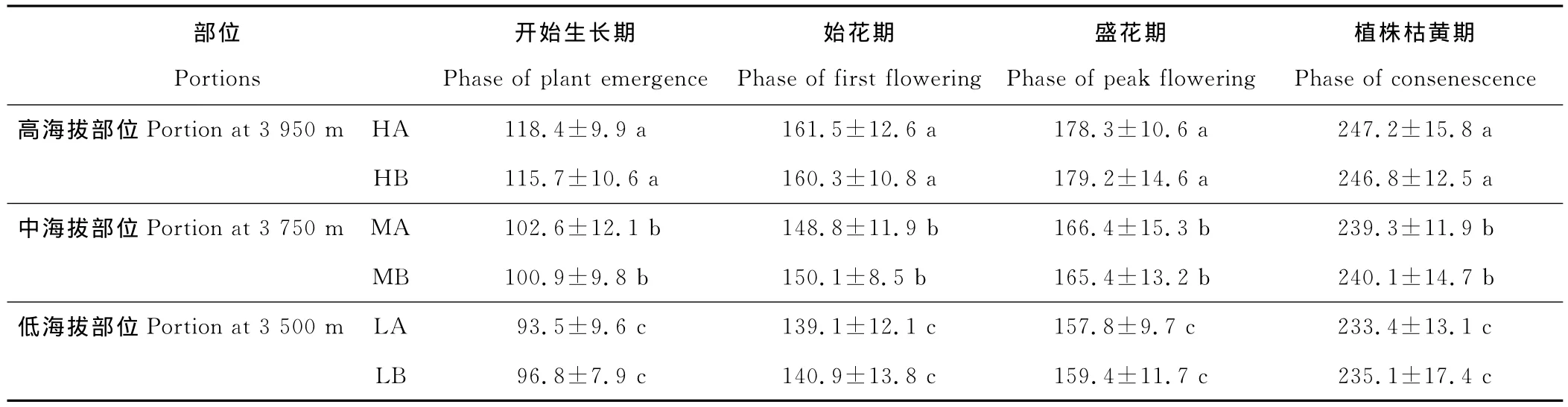
表3 3個部位物候期的比較Table 3 Comparison of phenological phases among three portions d
2.3 不同海拔部位對暗紫貝母鱗莖的影響
就對照樣方而言,低海拔的部位鱗莖平均單粒重量最高,隨著海拔的升高,鱗莖平均單粒重量降低(圖1)。和低海拔對照部位相比,高海拔對照部位的鱗莖重量減少了23.6%。從高海拔部位來看,移植植株(HB)和對照植株(HA)之間有明顯的差異,移植植株的單個鱗莖比對照植株大了13.2%;在低海拔部位,移植植株(LB)和對照植株(LA)之間也有明顯的差異,LA植株的鱗莖比LB植株大了9.7%;在中海拔部位沒有移植植株,A部位和B部位之間差別不明顯。顯然,低海拔部位的植株移植到高海拔后,其鱗莖有變小的趨勢;同樣,高海拔部位植株移植到低海拔后,其鱗莖有變大的趨勢。可見,鱗莖在高、低海拔部位之間的變化趨勢也可以從移植實驗得到證實。
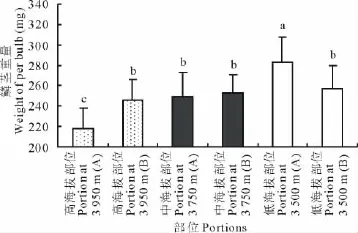
圖1 暗紫貝母鱗莖重量在海拔梯度上的變化Fig.1 Variation of bulbs’weight along the altitudinal gradient
3 討論
海拔的變化會引起融雪時間的變化,和高海拔部位相比,低海拔部位相對較高的氣溫會導致積雪的早融(表1),從而使得低海拔部位無雪期加長。有研究表明,在高山和脅迫環境中,無雪期長度是決定植物生長的主要因素[23,24]。生活在溫帶地區的高山植物,更多地受無雪期長度的影響[25]。在積雪晚融的高山區域,無雪期長度的縮短限制了植物積累更多的營養物質,而這些營養物質是來年生長必須的[26],從而影響植物的生長[27]。顯然,這些研究結果都認為無雪期長度對植物的生長有重大的影響。可見,在高海拔部位,暗紫貝母物候的推遲在很大程度上是由于融雪的推遲所引起的,其他學者對其他雪生植物的研究也得出了相似的結果[28,29]。另外,高、低海拔部位之間的平均氣溫(以3,4,5月為例)差值都在2.7℃以上(表2),而且在高海拔部位夜間溫度往往可達0℃以下[19]。Billings[27]認為,由于北極和高山植物的繁殖器官對低溫十分敏感,植物體暴露在夜間低溫中會增加凍害的風險。因此,在溫度相對較低的高海拔部位,由于夜間低溫的影響,植物開始生長的時間會相對較遲,這可能也是物候期推遲的原因之一。
由于低海拔部位物候期的提前,植物萌動生長開始較早,因此生長期相對較長;再加上低海拔處溫度相對較高,植物的生長相對較快,于是暗紫貝母可以積累較多的營養物質用于鱗莖的生長,故其鱗莖相對要大一些。也有一些研究認為,高山植物的個體生長要受積雪融化后夜間低溫的影響,這會導致早融部位的植株生長不好;而晚融部位植株所受影響較小,生長相對還要好一點[30,31]。這與本研究的結果不同,其原因可能是其他學者的研究主要集中于一個較小的海拔范圍內,不存在海拔梯度因素的影響,在這種情況下,融雪部位的夜間低溫成了決定植物生長的限制因子。而在本研究中,研究部位之間海拔相差很大,氣溫相差也很大,較大的氣溫差別使得不同海拔部位之間的融雪時間、生長期的長度以及植物生長的快慢都有顯著的不同,因此,在本研究中低海拔部位相對較高的氣溫和相對較長的無雪期對植物的生長起主要決定作用。
暗紫貝母鱗莖在海拔梯度上的變化可能與土壤含水量也有一定的關系。在高寒草甸群落中的研究表明,土壤含水量增加往往能使植物地下生物量減少而使地上生物量增加[29,32],干旱往往能使地下生物量增加[33]。在本研究中,低海拔部位由于土壤水分相對較低,其地下鱗莖相對較大,而在含水較多的高海拔部位則相反。
低海拔部位導致暗紫貝母物候期提前,還能導致其鱗莖產量有所提高。但不能據此認為,海拔越低對暗紫貝母的生長越好。因為暗紫貝母畢竟屬于高山雪生植物[2,3],在海拔過低的地方能否成活,以及其鱗莖產量會如何變化,這還有待于今后進一步的研究。
[1] 徐國鈞.中草藥彩色圖譜(修訂本)[M].福州:福建科學技術出版社,1997:210-213.
[2] 四川植物志編輯委員會.四川植物志(第七卷)[M].成都:四川民族出版社,1991:57-59.
[3] 青海植物志編輯委員會.青海植物志(第四卷)[M].西寧:青海人民出版社,1999:277-278.
[4] 桂鏡生,楊樹德.暗紫貝母與平貝母的資源狀況調查及市場供求分析[J].云南中醫學院學報,2008,31(6):36-39.
[5] 劉輝,陳士林,姚輝,等.暗紫貝母的資源學研究進展[J].中國中藥雜志,2008,33(14):1645-1648.
[6] 朱丹妮,蔣瑩,陳婷,等.組織培養川貝母化學成分和藥理作用的研究[J].中國藥科大學學報,1992,23(2):118-121.
[7] 李隆云,周裕書,代敏,等.暗紫貝母鱗莖再生組織培養技術研究[J].中國中藥雜志,1995,20(2):78-80.
[8] 韻海霞,陳志.暗紫貝母乙醚萃取物的GC-MS分析[J].青海醫學院學報,2009,30(2):133-135.
[9] 蔡朝暉,高山林,徐德然,等.不同培養條件及方法對組培暗紫貝母生長的影響[J].中國藥科大學學報,1991,23(6):367-369.
[10] 李雪松.暗紫貝母與其混淆品及偽品的鑒別[J].中國當代醫藥,2009,5:128-131.
[11] 潘莉,徐道華.暗紫貝母的真偽鑒別[J].中國藥業,2008,17(20):56-57.
[12] 劉明華.暗紫貝母與其常見偽品的性狀鑒別[J].首都醫藥,2007,14(12X):36.
[13] 劉紅巖.暗紫貝母及常見偽品性狀鑒別[J].時珍國醫國藥,2004,15(5):639.
[14] 鄭廣東.暗紫貝母及其混偽品的鑒別[J].海峽藥學,1998,10(4):49-51.
[15] 陳士林,肖小河,陳善墉.暗紫貝母的群落生態研究[J].中藥材,1989,12(11):5-8.
[16] 陳士林,肖小河,陳善墉.暗紫貝母植被分布格局的數值分析[J].西南師范大學學報(自然科學版),1997,22(4):416-420.
[17] 劉先齊.暗紫貝母與太白貝母的引種比較試驗[J].中國中藥雜志,1994,19(2):81-82.
[18] 李西文,陳士林.遮蔭下高原瀕危藥用植物川貝母(Fritillaria cirrhosa)光合作用和葉綠素熒光特征[J].生態學報,2008,28(7):3438-3446.
[19] Chen W N,Wu Y,Wu N,et al.Effect of snow-cover duration on plant species diversity of alpine meadows in the eastern Qinghai-Tibetan Plateau[J].Journal of Mountain Science,2008,5(4):327-339.
[20] Chen W N,Wu Y,Wu N,et al.Effect of snowmelt time on growth and reproduction of Pedicularis davidii var.pentodon in the eastern Tibetan Plateau[J].Plant Biosystems,2011,145(4):802-808.
[21] 馬德隆.松潘縣志[M].北京:民族出版社,1999:101-140.
[22] Galen C,Stanton M L.Consequences of emergence phenology for reproductive success in Ranunculus adoneus (Ranunculaceae)[J].American Journal of Botany,1991,78:978-988.
[23] Billings W D,Bliss L C.An alpine snowbank environment and its effects on vegetation,plant development,and productivity[J].E-cology,1959,40:388-397.
[24] Miller P C.Environmental and vegetation variation across a snow accumulation area in montane tundra in central Alaska[J].Holarctic Ecology,1982,5:85-98.
[25] Suzuki S,Kudo G.Short-term effects of simulated environmental change on phenology,leaf traits,and shoot growth of alpine plants on a temperature mountain,northern Japan[J].Global Change Biology,1997,3(Suppl.1):108-115.
[26] Wijk S.Performance of Salix herbaceain an alpine snow-bed gradient[J].Journal of Ecology,1986,74:675-684.
[27] Billings W D.Adaptations and origins of alpine plants[J].Arctic,Antarctic and Alpine Research,1974,6:129-142.
[28] Inouye D W,Morales M A,Dodge G J.Variation in timing and abundance of flowering by Delphinium barbeyi Huth(Ranunculaceae):the roles of snowpack,frost,and La Ni1a,in the context of climate change[J].Oecologia,2002,130:543-550.
[29] 李英年.高寒草甸植物地下生物量與氣象條件的關系及周轉值分析[J].中國農業氣象,1998,19(1):36-42.
[30] Kudo G.Effects of snow-free period on the phenology of alpine plants inhabiting snow patches[J].Arctic and Alpine Research,1991,23:436-443.
[31] Kudo G.Performance and phenology of alpine herbs along a snow-melting gradient[J].Ecological Research,1992,7:297-304.
[32] 劉偉,周華坤,周立.不同程度退化草地生物量的分布模式[J].中國草地,2005,27(2):9-15.
[33] Falk J H.The primary productivity of lawns in a temperate environment[J].Journal of Applied Ecology,1980,17:689-696.
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
