轉DsRed熒光蛋白的新月彎孢Curvularia lunata菌株構建
2012-09-28 01:43:56陳茂功韓志群林小虎王曉鳴周印富李洪杰
植物保護 2012年6期
陳茂功, 韓志群, 林小虎, 王曉鳴*, 周印富*, 李洪杰
(1.河北科技師范學院生命科技學院,秦皇島 066004;2.中國農業科學院作物科學研究所/農作物基因資源與基因改良國家重大科學工程,北京 100081)
在中國,玉米彎孢葉斑病是由新月彎孢[Curvularia lunata(Wakker)Boedijn]引起的一種葉部病害,目前幾乎在各玉米種植地區都有發生,一度成為北方地區主要葉部病害之一,并在遼寧西部地區引起嚴重的生產損失[1]。目前對病菌的研究主要集中在致病性分化、變異與致病機制等方面[2],但對其在玉米葉片上的侵染及發育過程尚不十分清楚。
農桿菌介導的遺傳轉化(Agrobacteriumtume faciens-mediated transformation,ATMT)是近年來被廣泛應用的一種真菌轉化技術,其免除了聚二乙醇(polyethylene glycol,PEG)介導和限制性內切酶介導整合(restriction enzyme-mediated integration,REMI)等轉化方法中原生質體制備的復雜過程,具有轉化效率高、易獲得穩定轉化子和T-DNA在真菌基因組中以單拷貝插入為主等優點[3]。自1998年de Groot等首次將ATMT法應用于絲狀真菌的遺傳轉化以來,已有大量的絲狀真菌通過ATMT法成功實現了遺傳轉化[4]。目前,ATMT技術也被用于彎孢菌的遺傳轉化、基因定點突變和功能研究[5]。紅色熒光蛋白(DsRed)來源于海葵體內的非常穩定的生物發光蛋白。DsRed和綠色熒光蛋白(GFP)具有相似的產生熒光機理,但DsRed的熒光信號能夠在組織中更加充分地轉換,其波長比GFP的長并具較高的噪音比,更易觀察。DsRed作為新一代報告基因,已經被用于果蠅、細菌、真菌、黏菌的標記研究[6-15]。在我國,DsRed 基因也被用于魚、酵母、病毒以及植物致病真菌等的研究中[16-20]。
本研究以hygB抗性基因為篩選標記,采用ATMT法將DsRed熒光蛋白基因整合入新月彎孢基因組中,實現新月彎孢的DsRed標記,并對TDNA插入突變體的主要表型特征進行鑒定,檢測基因的插入位點,為進一步的新月彎孢研究進行技術準備。
1 材料和方法
1.1 供試菌株、質粒和植物材料
分離自玉米的新月彎孢(C.lunata)由中國農業科學院作物科學研究所保存;攜帶DsRed基因的質粒pCAMDsRed由 Marina Franceschetti(John Inner Center,UK)惠贈。DsRed基因受構巢曲霉3-磷酸甘油醛脫氫酶基因gpdA的啟動子PgpdA調控,以潮霉素B抗性基因hyg作為選擇標記基因[21];高感彎孢葉斑病的玉米自交系‘黃早四’用于病菌致病力檢測。
1.2 農桿菌介導的遺傳轉化
通過對含潮霉素B濃度為0、30、50、75、100μg/mL和150μg/mL的PDA平板上新月彎孢生長狀況的檢測,確定野生型C.lunata對潮霉素B的耐受濃度為50μg/mL。
參照Mullins等的方法[22],進行農桿菌介導下DsRed基因導入C.lunata的操作。獲得轉化子菌落后,挑取陽性轉化子至含抗生素(頭孢霉素200μg/mL和潮霉素B 50μg/mL)的PDA平板上,待菌株長滿全皿且產孢后,用無菌水洗下孢子,并稀釋至1.0×102個/mL,取100μL孢子懸浮液均勻涂布到含潮霉素B 50μg/mL的PDA平板上,25℃培養2d后挑取單菌落,獲得單孢純化菌株。
1.3 轉化子熒光觀察及標記檢測
在熒光顯微鏡(Olympus BX41,日本)下觀察轉化子的紅色熒光表達。檢測DsRed基因和hyg基因整合結果時,采用CTAB法提取菌株DNA,以DsRed 基因引物(DsRed-F:5′-ACTCCTCCGAGGACGTCATCAA-3′;DsRed-R:5′-ACGTAGTGTAGTAGCCGGGCAGCT-3′)和潮霉素B抗性hyg基 因 引 物 (Hyg-8F:5′-GCAGACAGGAACGAGGACAT-3′; Hyg-8R: 5′-GCTCCATACAAGCCAACCAC-3′)進行擴增,產物以0.8%瓊脂糖凝膠電泳鑒定。潮霉素B抗性基因的Southern blotting分析根據試劑盒操作說明進行。
1.4 轉化子致病性相關性狀測定
將轉化子和野生型菌株用直徑4mm的打孔器打菌餅接于PDA平板上,25℃恒溫黑暗培養5d,測定菌絲生長速率;產孢量測定參照賀春萍等人的方法[23]。將轉化子及野生型菌株接種液體培養基(參照 Marcus等配制[24])中,25℃恒溫培養15d,每天120r/min振蕩1h。將培養液經雙層紗布過濾,過濾液轉入離心管,在4℃,10 000r/min高速離心,上清液即為酶提取液。纖維素酶系中的內切β-1,4-葡聚糖酶(Cx酶)活性測定參照 Lee等方法[25];果膠酶中的聚甲基半乳糖醛酸酶(polymethylgalacturonase,PMG)活性測定參照 Murata等方法[26];粗毒素的提取參照唐樹戈等方法[27],毒素生物測定所用離體葉片針刺法參照康紹蘭等[28],毒素致病性分析設3次重復,測定獨立進行2次。
1.5 轉化子的致病性測定
在自交系‘黃早四’4葉期時,采用毛筆涂抹法接種葉片,轉化子接種懸浮液中分生孢子濃度為1.0×106個/mL,接種后黑暗保濕36h,然后放置溫室(25℃)中正常管理。分別以無病菌接種液和野生型孢子接種液為陰性對照和陽性對照。接種5d后觀察葉片發病率并計算病情指數,方法參照李金堂等[29]。玉米植株、營養土及所用儀器用完后進行滅菌處理。
1.6 轉化子的初步應用
將Cl-DsRed 4轉化子轉接至含有50μg/mL潮霉素B的PDA平板,25℃培養7d,用無菌水洗下孢子鏡檢并將菌液稀釋至1.0×106個/mL,取400μL置于凹形載玻片的凹槽中,25℃黑暗保濕培養12h后在熒光顯微鏡下觀察孢子萌發的形態;將該孢子懸浮液接種‘黃早四’的葉片,黑暗保濕36h后放置溫室正常管理,4d后取葉片病斑處觀察菌絲侵染情況。
1.7 DsRed基因插入位點側翼序列分析
突變體T-DNA插入位點側翼序列的擴增采用TAIL-PCR,參照 Liu等[30]和徐榮旗等[31]的方法進行。膠回收PCR擴增產物,連接到pGM-T載體中轉入大腸桿菌感受態,篩選陽性克隆子進行測序。引物合成(表1)和測序工作由上海生工生物工程有限公司完成。

表1 hiTAIL-PCR擴增T-DNA插入位點側翼序列引物列表1)
2 結果與分析
2.1 新月彎孢的遺傳轉化與熒光表達特點
采用農桿菌介導法將DsRed基因整合至新月彎孢基因組中,獲得紅色熒光蛋白基因標記的菌株Cl-DsRed 1、Cl-DsRed 2、Cl-DsRed 3和 Cl-DsRed 4。利用DsRed和hyg基因特異性引物從4個轉化子基因組DNA中均擴增獲得579bp和800bp目的片段,質粒pCAMDsRed中也能擴增出相同片段,而野生型菌株中無擴增(圖1a,b),表明DsRed基因已轉入新月彎孢基因組中。Southern blotting檢測發現,4個轉化子分別出現位置不同的單一條帶,表明轉化子為T-DNA單拷貝插入,插入位點不同,無明顯的位點插入偏好(圖1c)。

圖1 轉化子中DsRed基因(a)和hyg基因(b)的PCR檢測與Southern雜交分析(c)
雖然轉化子外觀形態與野生型相同,但在熒光顯微鏡紫外光激發下,轉化子菌絲、分生孢子梗和分生孢子能發出明亮的紅色熒光(圖2a,b),而野生型無熒光表達(圖2c,d)。DsRed基因只在轉化子分生孢子原生質中表達而在孢子壁和隔膜中無表達(圖2e),分生孢子成熟后由于孢子壁加厚導致熒光減弱。對4個轉化子在普通PDA培養基上繼代培養5代后,重新轉接至含潮霉素B的培養基上,轉化子仍能夠正常生長,PCR檢測能夠擴增出目的片段,表明DsRed基因在轉化子中穩定遺傳。
2.2 轉化子致病相關性狀的改變
差異顯著性分析表明,轉化子Cl-DsRed 2的生長速度極顯著低于野生型菌株,其他轉化子無顯著差異;轉化子Cl-DsRed 3產孢量極顯著減少,Cl-DsRed 1的產孢量也受到一定程度的影響。各轉化子Cx酶均略有下降,但差異不顯著。而菌株間PMG 酶活性差異明顯,轉化子 Cl-DsRed 1、Cl-DsRed 2和Cl-DsRed 4的活性顯著高于野生型菌株,而Cl-DsRed 3則表現為下降。轉化子Cl-DsRed 3粗毒素的致病力極顯著下降,其他轉化子粗毒素的致病力也受到一定影響(表2)。
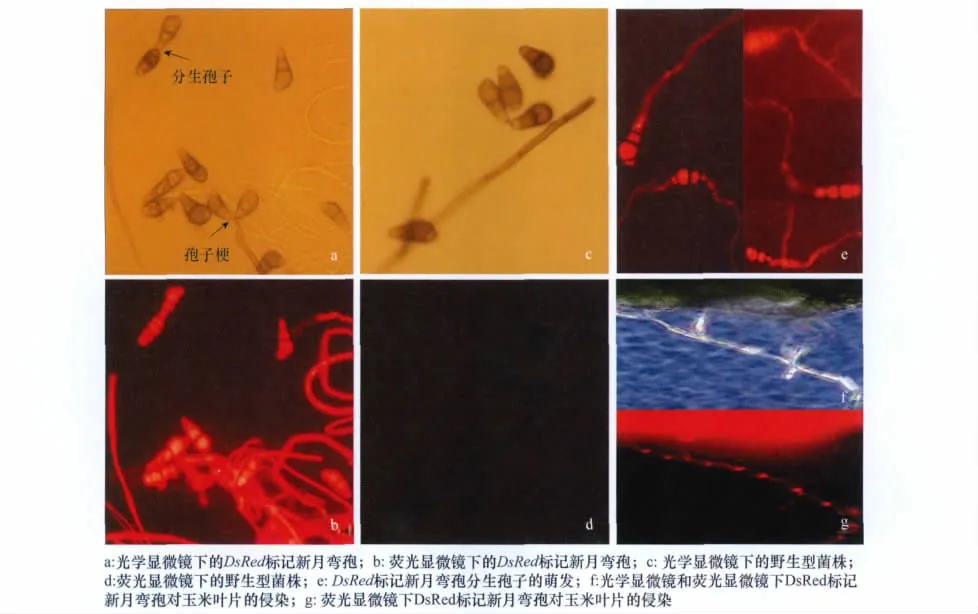
圖2 轉化子Cl-DsRed 4菌株的熒光表達

表2 轉化子致病相關性狀的比較分析1)
2.3 轉化子對玉米的致病性
Cl-DsRed 1、Cl-DsRed 2和Cl-DsRed 4與野生型菌株一樣,在感病自交系‘黃早四’的葉片上表現出較強的毒性和侵襲力,在接種后3d即可產生典型病斑;Cl-DsRed 3在接種5d后才出現典型病斑且病斑面積小,表明其侵襲力和致病力有所減弱。
2.4 轉化子的初步應用
新月彎孢為半知菌亞門絲孢綱絲孢目暗色絲孢科彎孢屬的真菌,分生孢子具3個隔橫4細胞,中隔常位于中央偏下處,從莖部數第3個細胞膨大;在黑暗培養12h后,孢子萌發率達到95%以上,一般從兩端顏色較淡的細胞長出芽管,少許孢子單端萌發;接種Cl-DsRed 4后5d,自交系‘黃早四’葉片上形成梭形褪綠透明的病斑(圖2f)。在熒光顯微鏡下,能夠看到紅色熒光蛋白在病菌侵染玉米葉片菌絲中的表達(圖2g)。
2.5 DsRed基因轉化子插入位點分析
在7個簡并引物中,AD5最適合彎孢菌的TAIL-PCR擴增,4個轉化子中有3個獲得特異性條帶,致病力表現下降的Cl-DsRed 3未獲得側翼序列,其他簡并引物未產生特異性擴增。將所獲得的序列提交NCBI后采用blastn比對,未發現高度同源的基因序列。初步比對結果(表3)表明,在Cl-DsRed 1轉化子中,DsRed蛋白編碼基因插入在一個未知功能的假定蛋白(939bp)中,但有效比對區域僅為216bp,插入位點為該假定蛋白基因的575和576位堿基之間。Cl-DsRed 2轉化子的DsRed蛋白編碼基因插入在ATP核糖磷酸轉移酶編碼基因(1 252bp)中,有效比對區域為473~583bp大小,插入位點為該基因的227和228位堿基之間。Cl-DsRed 2轉化子在菌絲生長速度和PMG酶活性方面表現出與野生型菌株的極顯著差異,可能與ATP核糖磷酸轉移酶活性受到干擾有關。Cl-DsRed 4轉化子的側翼序列未發現有登錄的相似基因序列。

表3 側翼序列基因的比對
3 討論
作為生物學研究工具,DsRed已對許多真菌成功實現了標記,但對玉米彎孢葉斑病菌的遺傳標記國內外尚未見報道。本研究采用根癌農桿菌介導的轉化系統,將紅色熒光蛋白標記基因插入新月彎孢基因組DNA中,獲得了DsRed基因穩定遺傳的轉化子,為該病菌在寄主植物中的侵染循環和致病機制研究提供了重要的材料。
對轉化子表型測定發現,轉化子與野生型相比,在一些性狀方面出現改變,可能是外源DNA片段的隨機插入位點影響了相關基因的表達,產生的突變體可用以研究彎孢菌生長發育相關基因和致病基因。在獲得的4個轉化子中,Cl-DsRed 4與野生型在致病性和其他性狀方面相近,適用于彎孢菌與玉米互作的研究。但由于玉米葉片自身紅色熒光較強,直接在葉表觀察彎孢菌絲的擴展、入侵等特征較為困難,只有在葉片橫切面才能有效觀察到發熒光菌絲。因此需要其他技術手段消除葉片自身熒光而不影響病菌熒光的觀察,以便能夠檢測到彎孢菌對玉米葉片的完整侵染循環過程。
hiTAIL-PCR技術可以通過對插入位點側翼序列的分析確定基因插入的位置,從而分析該基因功能與表型突變之間的關系[31]。本試驗還對其他20個突變體的插入位點側翼序列進行了擴增,獲得了其中13個序列,從本文的側翼序列比對結果中發現,具有相似性的登錄基因序列均來自與新月彎孢相近分類學地位的子囊菌格孢腔菌科(Pleosporace-ae)真菌:偃麥草核腔菌[Pyrenophora tritici-repentis(Died.)Drechsler]、圓核腔菌(Pyrenophora teres f.teres)、穎枯暗球腔菌(Phaeosphaeria nodorum)(無性態:斑污小球腔菌Leptosphaeria maculans),表明格孢腔菌科真菌在基因組成方面具有相似性。
[1] 王曉鳴,石潔,晉齊鳴,等.玉米病蟲害田間手冊-病蟲害鑒別與抗性鑒定[M].北京:中國農業科學技術出版,2010.
[2] Liu T,Liu L X,Hou J M,et al.Expression of green fluorescent protein in Curvularia lunata causing maize leaf spot[J].Canadian Journal of Plant Pathology,2010,32(2):225-228.
[3] Degefu Y,Hanif M.Agrobacterium tumefaciens-mediated transformation of Helminthos poriumturcicum,the maize leaf-blight fungus[J].Archives of Microbiology,2003,180:279-284.
[4] Mullins E D,Chen P X,Romaine P,et al.Agrobacterium transformation of Fusarium oxysporum:an efficient tool for insertional mutagenesis and gene transfer[J].Phytopathology,2001,58:2043-2052.
[5] Liu T,Liu L X,Jiang X,et al.Agrobacterium-mediated transformation as a useful tool for the molecular genetics study of the phytopathogen Curvularia lunata[J].European Journal of Plant Pathology,2010,126:363-371.
[6] Verkhusha V V,Otsuna H,Awasaki T,et al.An enhanced mutant of red fluorescent protein DsRed for double labeling and developmental timer of neural fiber bundle formation[J].The Journal of Biological Chemistry,2001,276:29621-29624.
[7] Nancharaiah Y V,Wattiau P,Wuertz S,et al.Dual labeling of Pseudomonas putida with fluorescent proteins for in situ monitoring of conjugal transfer of the TOL plasmid[J].Applied and Environmental Microbiology,2003,69(8):4846-4852.
[8] Mikkelsen L,Sarrocco S,Lübeck M,et al.Expression of the red fluorescent protein DsRed-Express in filamentous ascomycete fungi[J].FEMS Microbiology Letters,2003,223(1):135-139.
[9] Eckert M,Maguire K,Urban M,et al.Agrobacterium tumefaciens-mediated transformation of Leptosphaeriaspp.and Oculimaculaspp.with the reef coral gene DsRedand the jellyfish gene gfp[J].FEMS Microbiology Letters,2005,252:67-74.
[10] Fischera M,Haasea I,Wiesnerb S,et al.Visualizing cytoskeleton dynamics in mammalian cells using a humanized variant of monomeric red fluorescent protein[J].FEBS Letters,2006,580:2495-2502.
[11] Janus D,Hoff B,Hofmann E,et al.An efficient fungal RNA-silencing system using the DsRed reporter gene[J].Applied and Environmental Microbiology,2007,73(3):962-970.
[12] Elleuche S,P?ggeler S.Visualization of peroxisomes via SKL-tagged DsRed protein in Sordaria macrospora[J].Fungal Genetics Reports,2008,55:9-12.
[13] Helber N,Requena N.Expression of the fluorescence markers DsRed and GFP fused to a nuclear localization signal in the arbuscular mycorrhizal fungus Glomus intraradices[J].New Phytologist,2008,177(2):537-548.
[14] Nizam S,Singh K,Verma P K.Expression of the fluorescent proteins DsRed and EGFP to visualize early events of colonization of the chickpea blight fungus Ascochyta rabiei[J].Current Genetics,2010,56(4):391-399.
[15] Monteiro R A,Schmidt M A,de Baura V A,et al.Early colonization pattern of maize roots by Herbaspirillum seropedicae[J].Genetics and Molecular Biology,2008,31:932-937.
[16] 姜鵬,陳敏,白俊杰,等.外源性紅色熒光蛋白基因(RFP)在轉基因唐魚中的整合分析[J].農業生物技術學報,2010,18(5):968-974.
[17] 楊坤宇,劉如石,林鑒,等.紅色熒光蛋白在畢赤酵母中的高效表達[J].廈門大學學報(自然科學版),2005,44(3):410-415.
[18] 羅園媛,李慧娟,李東野,等.紅色熒光蛋白標記蛋白激酶B的慢病毒構建和表達[J].中華實驗外科雜志,2011,28(5):765-767.
[19] 楊峰,施李鳴,范素華,等.板栗疫病菌綠色熒光與紅色熒光蛋白共定位載體的構建[J].基因組學與應用生物學,2011,30(3):308-315.
[20] 吳磊,王曉鳴,徐榮旗,等.利用紅色熒光蛋白標記的輪枝鐮孢研究病原菌對與玉米根系的系統侵染和定殖[J].作物學報,2011,37(5):793-802.
[21] Eckert M,Maguire K,Urban M,et al.Agrobacterium tumefaciens-mediated transformation of the Leptosphaeriaspp.and Oculimaculaspp.with the reef coral gene DsRedand the jellyfish gene gfp[J].FEMS Microbiology Letters,2005,252:67-74.
[22] Mullins E D,Chen X,Romaine P,et al.Agrobacterium-mediated transformation of Fusarium oxysporum:an efficient tool for insertional mutagenesis and gene transfer[J].Phytopathology,2000,91:173-180.
[23] 賀春萍,邱世文.稻瘟菌T-DNA插入突變體的培養基篩選試驗[J].熱帶農業科學,2005,25(6):24-27.
[24] Marcus L,Schejter A.Single step chromatographic purification and characterization of the endopolygalacturonases and pectinesterases of the fungus,Botrytis cinerea Pers.[J].Physiological Plant Pathology,1983,23(1):1-13.
[25] Lee B H,Blackburn T H.Cellulase production by a thermophilic Clostridiumspecies[J].Applied and Environmental Microbiology,1975,30(3):346-353.
[26] Murata H,Chatterjee A,Liu Y,et al.Regulation of the production of extra cellular pectinase,cellulose,and protease in the soft rot bacterium Erwinia carotovora subsp.carotovora[J].Applied and Environmental Microbiology,1994,60(9):3150-3159.
[27] 唐樹戈,鄭其格,牟林,等.玉米彎孢葉斑病菌粗毒素的制備[J].西北農業學報,2010,19(1):57-59.
[28] 康紹蘭,張浩,黃悟芳.誘發玉米抗小斑病突變體的研究Ⅲ:影響玉米小斑病菌致病毒素作用的因素和生物測定[J].河北農業大學學報,1986,9(3):7-16.
[29] 李金堂,傅俊范.玉米彎孢菌葉斑病發病率與嚴重度的關系[J].植物保護學報,2009,36(6):569-570.
[30] Liu Y G,Chen Y.High-efficiency thermal asymmetric interlaced PCR for amplification of unknown flanking sequences[J].Biotechniques,2007,43:649-656.
[31] 徐榮旗,汪佳妮,陳捷胤,等.棉花黃萎病菌T-DNA插入突變體表型特征和側翼序列分析[J].中國農業科學,2010,43(3):489-496.
