應用EST-ILPs分子標記技術快速鑒定菟絲子屬種子
2012-09-28 01:44:06郭瓊霞黃可輝
植物保護 2012年6期
關鍵詞:檢測
郭瓊霞, 莊 蓉,2, 黃 振, 黃可輝,2*
(1.福建出入境檢驗檢疫局,福州 350001;2.福建農林大學植物保護學院,福州 350002)
近年來,分子標記已廣泛應用于遺傳圖譜構建、基因克隆、基因定位、遺傳多樣性分析、基因組比較、標記輔助育種等方面的研究[1-3]。由于EST來源于編碼區,內含子位置通常較為保守;而不同種或地理亞種間的內含子長度又存在差異,故用它建立的ILPs分子標記在種間鑒定上具有優越性。采用ILPs(intron length polymorphisms)分子標記在菟絲子屬的分類鑒定上國內外均未見報道。因此本研究利用旋花科(Convolvulaceae)番薯屬(Ipomoea)的EST(expressed sequence tags,Tags,表達序列標簽)序列,通過網站(http:∥ibi.zju.edu.cn/pgl/pip/index.html)查找內含子和外顯子,然后根據網站自動設計的引物,再對9個菟絲子的樣品進行引物的篩選,尋找種間特異性條帶從而達到分類鑒定的目的。
1 材料與方法
1.1 供試材料
本試驗選用菟絲子屬(Cuscuta)5個近似種(含不同的地理型)的種子作為研究材料。各個種的種子材料來源詳見表1。

表1 材料來源
1.2 總DNA的提取與凝膠電泳檢測
1.2.1 菟絲子總DNA的提取
選取菟絲子的種子為材料,采用SDS法提取總 DNA[4]。
1.2.2 凝膠電泳檢測
采用高必達等[5]設計的一對能檢測ITS序列的特異 性 引 物:A1(5′-TCGTAACAAGGTTTCCGTAG-3′)與 A2(5′-AGGAGGCCAGGATCTATT-3′)進行PCR擴增。擴增體系為:25μL(10×PCR buffer,25mmol/L MgCl2,10mmol/L dNTP,5 U/μL Taq DNA 聚合酶,10μmol/L 引物,模板DNA約為100ng/μL)。反應程序為:94℃5min;94℃1min,55℃1min,72℃1min,35個循環;72℃7min。擴增產物經1.5%瓊脂糖電泳分離,檢測400~500bp左右的擴增片段。從而根據電泳圖上此擴增片段的有無判斷是否提取出DNA。
1.3 ILPs標記的獲得和ILPs-PCR
ILPs標記是由浙江大學汪旭升、趙向前等根據已公布全基因組序列開發的一種內含子長度多態性標記[6]。從網站(http:∥ibi.zju.edu.cn/pgl/pip/index.html)上公布的ILPs標記中選擇引物。選取旋花科番薯屬已經設計好的引物10對(如表2所示),對菟絲子樣品進行PCR擴增,進而選擇特異性片段。
ILPs-PCR反應體系總體積為20μL,其中含100ng模 板 DNA;上、下 游 引 物 各 0.5μmol/L;200μmol/L dNTP;1.5mmoL/L MgCl2;1UTaq DNA聚合酶;2μL 10×PCR反應緩沖液。PCR擴增程序:94℃ 5min;94 ℃ 46s,52.5 ℃ 45s,72 ℃1min,35個循環;72℃10min。擴增產物經1.5%瓊脂糖電泳分離。對于擴增產物非特異性條帶多的引物進一步優化起始退火溫度,調整范圍為50~58℃。每對引物組合至少重復2次以確保擴增結果的可靠性。擴增產物上樣于6%的非變性聚丙烯酰胺凝膠,320V電泳1h45min。參照許紹斌等[7]的方法進行銀染。

表2 試驗所用的引物
2 結果與分析
2.1 DNA凝膠電泳的檢測
試驗結果表明,采用SDS法提取菟絲子屬5個近似種(含不同的地理型)種子的總DNA,經過凝膠電泳,其顯示在400~500bp得到一條明亮的特異條帶(如圖1所示)。從而證明所提取的菟絲子種子DNA有效,最后將已提取的DNA保存于-20℃備用。

圖1 利用引物A1和A2的rDNA-ITS PCR擴增圖譜
2.2 聚丙烯酰胺凝膠電泳檢測
因為退火溫度是影響PCR擴增結果的重要因素,所以對試驗條件進行調整與優化,經試驗得出結果:退火溫度為52.5℃可獲得滿意的分型效果。因此設定PCR擴增程序為:94℃5min;94℃46s,52.5℃45s,72℃1min,35個循環;72℃10min;于4℃保存。
根據引物IP9-1和IP9-2的聚丙烯酰胺凝膠電泳所檢測的結果(如圖2)所示,不同種的菟絲子呈現多態性的帶型。

圖2 IP9-1和IP9-2引物在材料1~9中的擴增產物(6%非變性聚丙烯酰胺凝膠電泳)
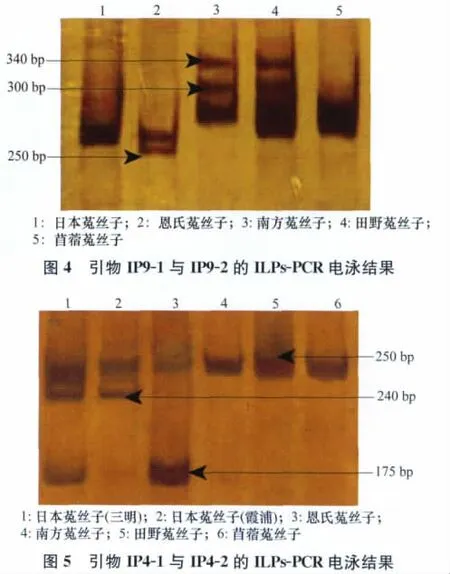
利用引物IP9-1和IP9-2擴增的試驗結果顯示:菟絲子屬5個近似種(含不同的地理型)9個種子樣品之間的內含子長度表現為5種基因型AB、A、ABC、BD、ABD。三明日本菟絲子與苜蓿菟絲子內含子長度的基因型均為AB;霞浦日本菟絲子內含子長度基因型為A;恩氏菟絲子內含子長度的基因型為ABC;南方菟絲子內含子長度基因型為BD;福州馬尾、霞浦、福州鼓樓、新疆這4個地理型的田野菟絲子內含子長度的基因型都是ABD。從圖2可以看出恩氏菟絲子在250bp左右具有一條特異性條帶,使其可以與其他的菟絲子區分開。南方菟絲子與田野菟絲子在內含子長度的基因型上也具有差異;因此可以通過內含子基因型的條帶多態性將田野菟絲子與南方菟絲子區分開。
用引物IP4-1和IP4-2擴增的結果(如圖3)顯示,菟絲子屬9個樣品之間的內含子表現為4種基因型EFG、EF、EG、E。三明日本菟絲子內含子長度的基因型為EFG;霞浦日本菟絲子內含子長度基因型為EF;恩氏菟絲子內含子長度基因型為EG;南方菟絲子、田野菟絲子、苜蓿菟絲子擴增結果相同,基因型均為E。從圖3可以得出以下結果:兩種地理型的日本菟絲子在240bp左右具有一條特征條帶,使其可以與其他種的菟絲子區分開;苜蓿菟絲子與日本菟絲子在內含子基因型上具有差異,可以通過條帶多態性將兩者區分開。從圖3還可以看出,不同地理型的日本菟絲子在基因型上也有所不同,三明日本菟絲子在175bp處有一條帶,但是霞浦日本菟絲子在175bp處卻沒有此條帶。
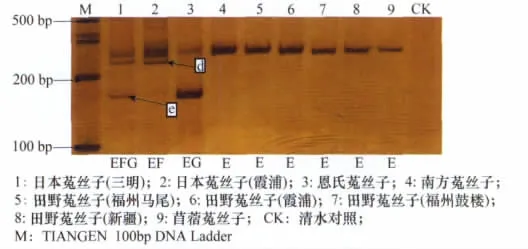
圖3 IP4-1和IP4-2引物在材料1~9中的擴增產物(6%非變性聚丙烯酰胺凝膠電泳)
通過圖4、圖5電泳結果的多態性綜合分析可以將本試驗所選取的菟絲子5個近似種逐一鑒別開。
2.3 聚類分析以及分子系統樹
根據選擇的10對引物(如表2)擴增出的穩定條帶,利用平均連鎖聚類方式(UPGMA)軟件進行聚類分析,生成9個供試材料的親緣關系樹狀圖(如圖6)。
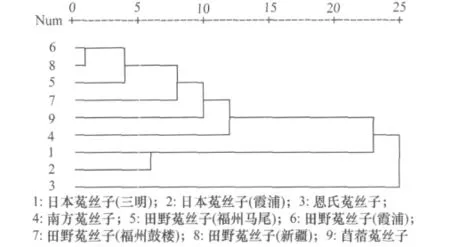
圖6 根據菟絲子近似種的ILPs分子標記構建的種間親緣關系圖
從圖6可以看出:田野菟絲子、苜蓿菟絲子、南方菟絲子的親緣關系比較接近;這3種與日本菟絲子、恩氏菟絲子之間的親緣關系較遠。
3 討論
本研究對菟絲子的5個近似種的內含子長度多態性進行了ILPs-PCR分析。從本試驗結果看出,a條帶可作為恩氏菟絲子的特異性條帶(如圖2示);d條帶可作為日本菟絲子特異性條帶(如圖3示)。利用ILPs分子標記檢測菟絲子近似種是菟絲子種間鑒定的新途徑。隨著ILPs分子標記檢測的深入研究與發展,該技術將為我國口岸雜草檢疫鑒定提供一種快速、準確的檢測方法,同時也為雜草檢疫科學理論的發展奠定良好的基礎。
通過本研究結果可知,不同地理型的日本菟絲子在內含子多態性的基因型上具有差異。因此可對不同地理區域的同種菟絲子進行進一步的比較研究,從而完善菟絲子分類鑒定的依據。還可以進一步從網站(http:∥ibi.zju.edu.cn/pgl/pip/index.html)上公布的ILPs標記中選擇引物進行篩選,進而選擇菟絲子種間特異性片段,完善并實現菟絲子種間的快速檢驗檢測。
[1] 吳海濱,朱汝財,李迪.一個內含子長度多態性標記與栽培稻F1花粉育性基因座連鎖[J].分子植物育種,2006,4(3):323-328.
[2] 曾華宗,鄭成木,朱穩,等.甘蔗種質間親緣關系及特異標記的RAPD分析[J].植物遺傳資源學報,2003,4(2):99-100.
[3] 盧泳全,汪旭升,黃偉素,等.基于水稻內含子長度多態性開發禾本科擴增共有序列遺傳標記[J].中國農業科學,2006,39(3):433-439.
[4] 王得元,鄧義才,李乃堅.RAPD標記檢測蔬菜種子純度中DNA提取方法研究[J].廣東農業科學,1999(3):13-14.
[5] 高必達,程毅,朱水芳,等.基于ITS序列的菟絲子PCR鑒定[J].湖南農業大學學報(自然科學版),2006,32(4):368-370.
[6] 汪旭升,趙向前,盧泳全,等.一種新型的分子標記:內含子長度多態性[C]∥2005植物分子育種國際學術研討會論文集,2005.
[7] 許紹斌,陶玉芬,楊昭慶,等.簡單快速的DNA銀染和膠保存方法[J].遺傳,2002,24(3):335-336.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
