肝纖維化病理過程中ZEB1和ZEB2的動態表達變化及意義*
2012-11-06 05:47:53謝汝佳張成俊程明亮
中國病理生理雜志 2012年9期
關鍵詞:模型
韓 冰, 謝汝佳, 洪 琴, 張成俊, 楊 勤, 程明亮
(貴陽醫學院病理生理教研室,貴州 貴陽 550004)
1000-4718(2012)09-1639-05
2012-03-23
2012-07-25
貴州省科技國際合作項目(No.黔科合G字【2011】7013號)
△通訊作者 Tel:0851-6908578;E-mail:hanbing791203@163.com
肝纖維化病理過程中ZEB1和ZEB2的動態表達變化及意義*
韓 冰△, 謝汝佳, 洪 琴, 張成俊, 楊 勤, 程明亮
(貴陽醫學院病理生理教研室,貴州 貴陽 550004)
目的觀察慢性肝損傷致肝纖維化過程中上皮-間質轉化(EMT)調節蛋白鋅指E盒結合同源盒蛋白(ZEB)1和ZEB2的動態表達變化并探討其調節機制。方法雄性Wistar大鼠40只,隨機分為正常對照組、2周、4周、6周和8周模型組,每組各8只,模型組按3 mL/kg體重的劑量皮下注射60%CCl4,每隔3 d注射1次,處死大鼠后測定肝臟指數、血清丙氨酸氨基轉移酶(ALT)和天冬氨酸氨基轉移酶(AST)活性,觀察肝組織病理改變;免疫組織化學法檢測肝組織中ZEB1、ZEB2、E-鈣黏蛋白(E-cadherin)和α-平滑肌肌動蛋白(α-SMA)的表達情況;real-time RT-PCR方法檢測肝臟組織中ZEB1及ZEB2 mRNA的表達變化。結果模型各組大鼠肝臟指數、血清ALT和AST活性顯著高于正常對照組(P<0.01),8周模型組肝纖維化明顯;隨著肝臟損傷逐漸加重,纖維化程度加深,E-cadherin蛋白的表達顯著降低,α-SMA蛋白表達顯著升高,ZEB1和ZEB2蛋白和mRNA表達量也逐漸增加,8周模型組肝組織中ZEB1和ZEB2蛋白(46.42±14.36和57.71±13.32)與mRNA(189.00±47.39和277.28±48.55)表達較正常組顯著增加(P<0.01)。結論ZEB1和ZEB2蛋白及mRNA表達量隨纖維化程度的加重而增加,提示EMT可能通過ZEB1和ZEB2參與肝纖維化的發生發展。
肝纖維化; 慢性肝損傷; 鋅指E盒結合同源盒蛋白
鋅指E盒結合同源盒蛋白(zinc finger E-box binding homeobox protein,ZEB)是一類具有鋅指結構的蛋白,包含2個成員ZEB1和ZEB2,ZEB2又稱Smad相互作用蛋白1(Smad-interacting protein 1,SIP1),由于都具有相同鋅指結構而能與含有CACCT的E-box序列和CACCT(G)的E-box相似序列結合,進而調控靶基因的表達。目前已知ZEB1和ZEB2能與上皮細胞內的E鈣黏蛋白(E-cadherin)基因的啟動子序列中的E-box(CACCTC/G)序列結合而抑制其轉錄,從而促進上皮-間質轉化(epithelial-mesenchymal transition,EMT)的發生[1]。EMT是指原本緊密排列的上皮細胞在某種因素的作用下轉化成間質細胞的過程。近年的研究發現,EMT除與肺纖維化和腎間質纖維化發病機制有關外[2-3],還參與了肝纖維化發生發展。但目前有關EMT通過其調控因子ZEB1和ZEB2參與調控慢性肝損傷及肝纖維化發病機制的研究鮮有報道。因此,本研究通過大鼠慢性肝損傷致肝纖維化模型,觀察慢性肝損傷發生致肝纖維化形成過程中,肝組織中ZEB1、ZEB2、E-cadherin和α-平滑肌肌動蛋白(α-smooth muscle actin,α-SMA)的動態表達變化,探討慢性肝損傷及肝纖維化發病過程中肝臟EMT變化特點及其通過ZEB1和ZEB2調控其發生發展的可能機制。
材 料 和 方 法
1材料
1.1動物與主要試劑 清潔級雄性Wistar大鼠40只,體重(200±20)g,購自貴陽醫學院動物實驗中心, ZEB1、ZEB2、E-cadherin和α-SMA多克隆抗體購自北京博奧森生物試劑公司;免疫組化EnvisionⅡ抗檢測試劑盒購自Gene;TRIzol購自Invitrogen,逆轉錄試劑盒購自Fermentas,SYBR Green購自Applied Biosystems,ZEB1和ZEB2的引物由TaKaRa公司代為設計合成。
1.2模型制備與分組 實驗動物適應性喂養1周后,隨機分成4組,每組8只,按3 mL/kg體重的劑量皮下注射60%CCl4,每隔3 d注射1次,同時正常飼料喂養,分別于喂養2周、4周、6周、8周后處死;另設正常對照組,共8只大鼠,給予正常飼料喂養,按3 mL/kg體重的劑量皮下注射生理鹽水每隔3d注射1次,分別于2周、4周、6周、8周各處死2只;所有處死大鼠收集血液及肝臟標本待測相關指標。
2方法
2.1計算肝臟指數 肝臟指數=肝臟重量(mg)/體重(g)。
2.2血清生化指標檢測 血清丙氨酸氨基轉移酶 (alanine aminotransferase,ALT)和天冬氨酸氨基轉移酶(aspartate aminotransferase,AST)檢測采用Siemens Advia 1650全自動生化分析儀測定。
2.3病理學觀察 取同一部位肝葉,常規石蠟包埋切片,行HE及Van Gieson(VG)染色,光鏡下觀察肝臟病理變化,參照纖維化分級法[4]進行膠原纖維增生程度半定量分析。
2.4肝組織ZEB1、ZEB2、E-cadherin和α-SMA蛋白表達檢測 采用Envision兩步法按照試劑盒說明操作。陰性對照以PBS液代替I抗。結果判定:ZEB1、ZEB2和α-SMA光鏡下觀察胞漿染成棕黃色的細胞為陽性,E-cadherin光鏡下觀察胞核染成棕黃色的細胞為陽性。400倍下觀察不少于5個高倍鏡視野,記數陽性細胞個數并計算陽性細胞所占百分比(使用Olympus Image-Pro Plus圖像分析軟件系統完成:采集400倍視野下5張圖片,然后通過軟件統計每張圖片上的陽性細胞數并計算陽性百分率)。
2.5肝組織ZEB1和ZEB2 mRNA表達量檢測 采用實時熒光定量RT-PCR的方法檢測,用TRIzol提取大鼠肝臟的總RNA,隨后采用紫外分光光度法進行質量控制及確定濃度。各取1μg總RNA為逆轉錄模板,使用ZEB1、ZEB2特異性逆轉錄引物進行逆轉錄反應,反應條件為:25 ℃ 10 min; 48 ℃ 60 min; 95 ℃ 5 min。實時熒光定量PCR反應條件為95 ℃ 10 min,95 ℃ 15 s,60 ℃ 60 s,40 個循環。PCR 反應在實時定量PCR 儀進行檢測, 以β-actin作為內參照,2-ΔΔCt為目的基因的相對表達強度,所有樣本均重復3孔。具體引物序列如下:β-actin上游引物5’-GAGAGGGAAATCGTGCGTGAC-3’,下游引物5’-CATCTGCTGGAAGGTGGACA-3’;ZEB1 上游引物5’-TGGCAAGACAACGTGAAAGA-3’,下游引物5’-AACTGGGAAAATGCATCTGG-3’;ZEB2 上游引物 5’-TAGCCGGTCCAGAAGAAATG-3’,下游引物5’-GGCCATCTCTTTCCTCCAGT-3’。
3統計學處理
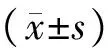
結 果
1各組大鼠肝臟指數、血清ALT和AST活性的變化
模型組2周組大鼠肝臟指數均高于正常組大鼠(P<0.05);4周組、6周組和8周組大鼠肝臟指數均顯著高于正常組大鼠(P<0.01);模型各組大鼠血清AST和ALT活性均較正常組大鼠顯著增加(P<0.01),見表1。
表1各組大鼠肝臟指數、血清ALT和AST活性

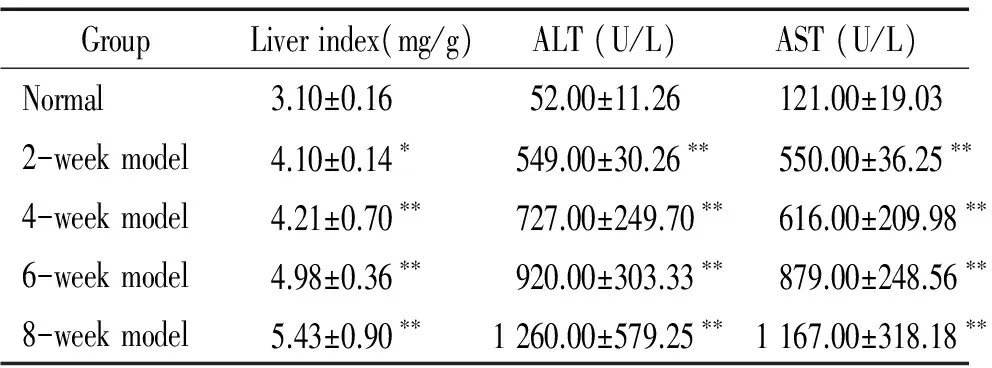
GroupLiverindex(mg/g)ALT(U/L)AST(U/L)Normal3.10±0.1652.00±11.26121.00±19.032-weekmodel4.10±0.14?549.00±30.26?? 550.00±36.25??4-weekmodel4.21±0.70??727.00±249.70?? 616.00±209.98??6-weekmodel4.98±0.36??920.00±303.33?? 879.00±248.56??8-weekmodel5.43±0.90??1260.00±579.25?? 1167.00±318.18??
*P<0.05,**P<0.01vsnormal group.
2肝臟病理變化
各組大鼠肝組織經HE及VG染色,光鏡下可見正常組大鼠肝細胞以中央靜脈為中心呈放射狀排列,無膠原纖維增生;模型2周組大鼠肝細胞內可見大小不一脂肪滴,部分氣球樣變,呈脂性肝炎表現;模型4周組大鼠肝細胞脂肪樣變明顯,可見部分空泡樣變,以微泡型為主,少數細胞為大泡型改變,少量匯管區可見纖維結締組織輕度增生;模型6周組大鼠肝細胞以大泡型脂肪變明顯,匯管區可見較多纖維結締組織增生,少量炎性細胞浸潤,假小葉形成不明顯;模型8周組大鼠肝細胞脂肪樣變嚴重,呈廣泛大泡型改變,肝小葉結構破壞,肝索排列紊亂,匯管區纖維結締組織大量增生,內有較多炎性細胞浸潤,膠原纖維形成纖維間隔,可見假小葉形成。病理學半定量結果顯示,模型2周組病理改變與正常組相比差異無統計學意義,模型4周組病理學改變與正常組相比差異有統計學意義(P<0.05),模型6周和8周組病理學改變與正常組相比有明顯差異(P<0.01),說明纖維化變明顯,見圖1、表2。

Figure 1. HE staining of rat hepatic tissues in each group (×400).A:normal group; B:2-week model group; C: 4-week model group; D: 6-week model group; E:8-week model group.
圖1各組大鼠肝組織HE染色結果
表2 各組大鼠肝纖維化程度分級
*P<0.05,**P<0.01vsnormal group.
3肝組織E-cadherin和α-SMA蛋白表達的變化
正常對照組大鼠肝組織未見α-SMA蛋白陽性表達,而E-cadherin蛋白則廣泛表達于正常對照組大鼠肝細胞,光鏡下可見細胞核染色棕黃;α-SMA蛋白隨著模型組大鼠肝臟肝損傷程度的加重及肝纖維化的發生而表達逐漸增加,在模型4周、6周、8周組大鼠肝組織中α-SMA蛋白表達于部分肝細胞及大量纖維間隔,較空白對照組表達明顯增加(P<0.01);E-cadherin蛋白在肝細胞胞核內隨著肝損傷及纖維化程度的加重而表達明顯減少,模型6周和8周組E-cadherin蛋白僅在組織部分結構相對完好區域的部分肝細胞內見少量表達,較正常對照組顯著降低(P<0.01),見圖2、表3。
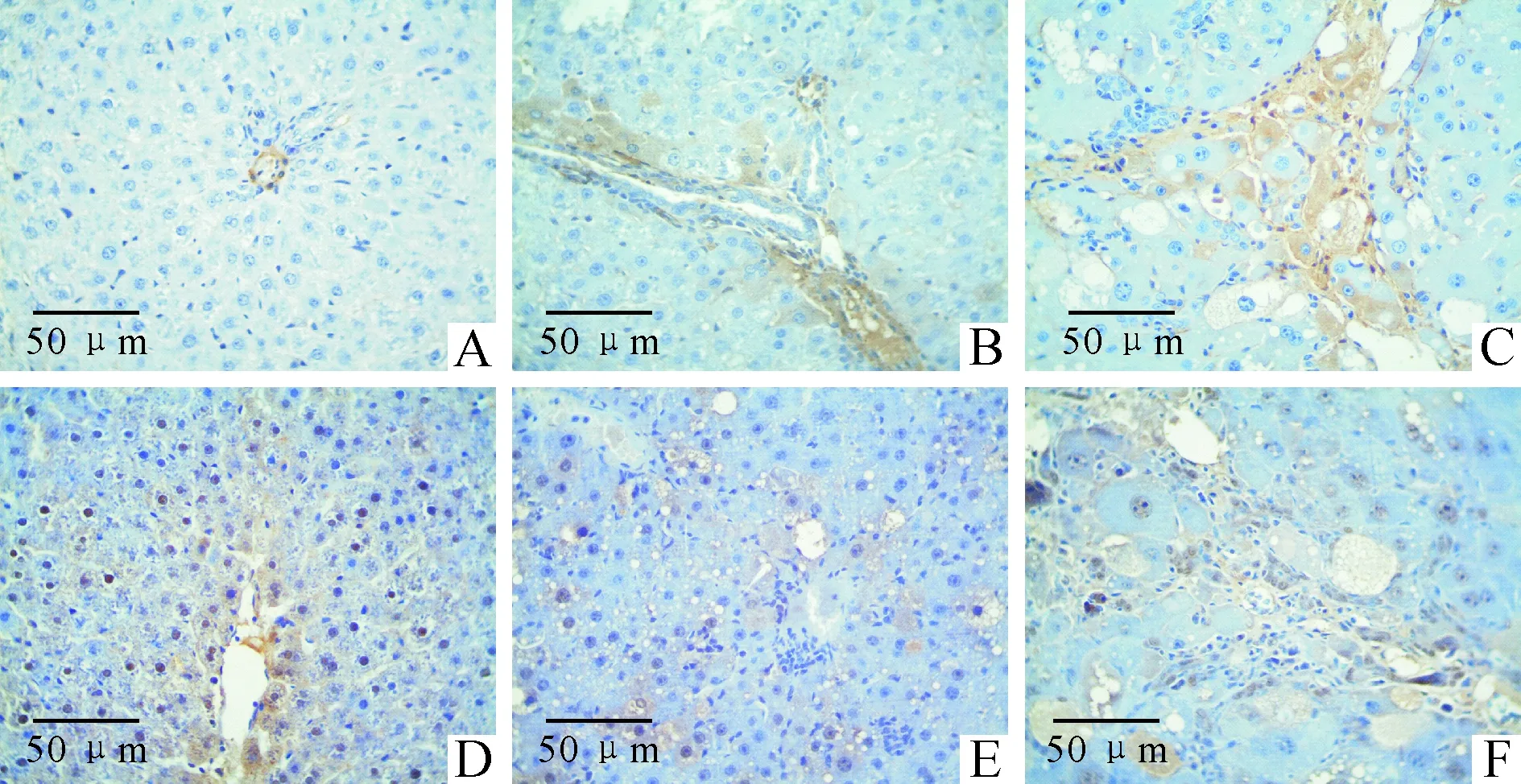
Figure 2. Expression of α-SMA and E-cadherin in rat hepatic tissue of each group.A:normal group(α-SMA); B:4-week model group (α-SMA); C: 8-week model group (α-SMA); D: normal group (E-cadherin); E: 4-week model group (E-cadherin); F: 8-week model group (E-cadherin).
圖2各組大鼠肝組織α-SMA和E-cadherin免疫組化結果
表3各組大鼠肝臟中α-SMA和E-cadherin的表達

Groupα-SMAE-cadherinNormal1.72±0.0690.48±4.922-weekmodel2.17±0.1886.28±6.354-weekmodel7.88±0.25??82.11±7.286-weekmodel19.78±4.28??24.37±3.25??8-weekmodel37.84±7.04??19.52±4.25??
**P<0.01vsnormal group.
4各組大鼠肝組織ZEB1和ZEB2蛋白及mRNA表達變化
正常對照組ZEB1/ZEB2蛋白表達較少,模型2周和4周組ZEB1和ZEB2蛋白及mRNA表達量較
正常對照組有所升高,但無顯著差異;而模型6周和8周組大鼠肝組織中可見ZEB1/ZEB2蛋白廣泛在小葉內氣球樣變的肝細胞及纖維化的肝細胞胞漿中,呈強陽性表達,色棕黃色,mRNA表達量也相應升高,與正常對照組表達量存在顯著差異(P<0.01),見圖3、表4。

Figure 3. Expression of ZEB1/ZEB2 in rat hepatic tissue of each group. A:normal group; B: 8-week model group (ZEB1); C: 8-week model group (ZEB2).
圖3各組大鼠肝組織ZEB1和ZEB2免疫組化結果
表4 各組大鼠肝臟中ZEB1/ZEB2蛋白和mRNA的表達
**P<0.01vsnormal group.
討 論
EMT是指上皮實質細胞向間質細胞的轉化,也就是上皮細胞失去其上皮表型特征而逐漸獲得間質細胞表型特征的過程[5]。纖維化被認為是器官受到慢性損傷時的一種創傷愈合過程,在正常生理情況,如果損傷愈合,由于炎癥逐漸減弱,EMT也會隨之停止,從而完成修復過程;但是如果炎癥不斷刺激,使EMT持續存在,則可生成過量肌成纖維細胞(MFLC),產生過多細胞外基質,便會導致器官纖維化[5-6]。本研究通過CCl4復制慢性肝損傷致肝纖維化模型,觀察病程演變過程中EMT動態變化。通過對上皮標志蛋白E-cadherin及間質標志蛋白α-SMA在各組大鼠肝臟組織中動態變化的觀察,發現隨著肝損傷及肝纖維化程度的加重,E-cadherin呈表達逐漸減少趨勢,α-SMA表達則呈明顯增加趨勢,特別是模型8周組大鼠肝臟組織中E-cadherin及α-SMA表達皆與正常對照組有顯著差異。因此實驗結果表明在肝纖維化形成過程中,上皮細胞標志蛋白表達逐漸減少而間質細胞特征蛋白表達逐漸增多,提示EMT參與了慢性肝損傷及肝纖維化的發生發展過程,并且隨著損傷程度的加重EMT的作用也相應加強。
雖然有研究表明,人轉化生長因子β(transforming growth factor β,TGF-β)和Wnt等信號蛋白可誘導EMT的發生,但是其介導EMT在纖維化中的作用卻不甚明了,考慮和具體的組織器官及其所處的微環境有關[7-8]。在肝纖維化研究中發現,靜息肝星狀細胞轉化為肌成纖維細胞的過程類似EMT[9]。有學者研究了不同來源的肌成纖維細胞樣肝星狀細胞,發現它們均共同表達間質及上皮祖細胞標志[10]。除了肝星狀細胞之外,成纖維細胞和上皮細胞的EMT以及骨髓來源的干細胞也發揮了重要作用[9]。Dooley等[11]研究表明肝細胞可被TGF-β1/Smad信號通路激活發生EMT,并表達I型膠原蛋白,提示肝細胞的EMT與肝纖維化密切相關。雖然目前肝纖維化中EMT的正向調控機制尚不清楚,但是由于E-cadherin表達的下降和缺失是EMT最重要的生物學特征,因此通過調解E-cadherin表達而參與EMT的調節機制成為研究熱點。ZEB1和ZEB2能通過與E-cadherin基因啟動子上的CACCT(G)序列結合抑制其表達,導致上皮細胞失去其表型特征轉化成為間質細胞,導致EMT的發生[12]。ZEB1可抑制多種重要的上皮分化和細胞黏附因子,包括細胞極性基因Crumbs3(Crb3)、HUGL2(human lethal giant larvae homologue 2)和Pals1(protein associated with Lin seven 1)相關的緊密連接蛋白,從而抑制E-cadherin,誘發EMT[13]。本研究在觀察慢性肝損傷致肝纖維化過程中EMT動態變化的同時,還觀察了在各組大鼠肝臟組織中ZEB1、ZEB2蛋白及mRNA表達量的變化,結果提示ZEB1、ZEB2在蛋白和基因水平隨著慢性肝損傷的發生及肝纖維化的形成,表達量也相應增加,表達變化趨勢與E-cadherin的變化趨勢相反,進一步驗證ZEB家族可通過抑制E-cadherin的表達而促進EMT的發生發展。本課題組前期的研究已發現CCl4可引發大鼠肝細胞TGF-β/Smad信號通路的激活,而ZEB家族已被證實與如TGF-β/ Smad、MAPK等多條參與肝纖維化發生發展的信號通路存在相互調節關系,因此我們推測在肝纖維化發生過程中,肝細胞EMT發生的可能機制是TGF-β/Smad信號通路激活ZEB1和ZEB2表達,ZEB1和ZEB2進一步抑制肝細胞表達E-cadherin,促進肝細胞發生EMT轉化為成纖維細胞,進而表達間質細胞特征蛋白α-SMA等,同時轉化產生的肌成纖維細胞表達大量的細胞外基質如Ⅰ型膠原蛋白,導致細胞外基質沉積最終導致肝纖維化發生。因此探明ZEB家族蛋白在肝纖維化發生機制中的作用,對闡明EMT與肝纖維化發生發展及相互調節關系有重要作用,其具體調節機制正在后續研究中。
[1] Naganuma S, Ohashi S, Kimura S,et al.ZEB1 and ZEB2 promote EMT and invasion in esophageal squamous cell carcinoma[J].Cancer Res, 2010, 70(8 Suppl 1): 2293.
[2] 徐西振, 鄭常龍, 凃 玲,等. 銀杏葉提取物對轉化生長因子β1誘導的人腎小管上皮細胞轉分化的影響及其機制[J].中國病理生理雜志,2008,24(11):2235-2238.
[3] Park SH,Choi MJ,Song IK,et al. Erythropoietin decreases renal fibrosis in mice with ureteral obstruction: role of inhibiting TGF-β-induced epithelial-to-mesenchymal transition[J].J Am Soc Nephrol, 2007,18(5): 1497-1507.
[4] 溫靜靜,謝汝佳,韓 冰,等.肝纖維化大鼠肝細胞內質網形態及GRP78表達的研究[J]. 中國病理生理雜志,2011,27(11):2210-2213.
[5] Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition[J].J Clin Invest, 2009, 119(6):1420-1428.
[6] Kalluri R.EMT:when epithelial cells decide to become mesenchymal-like cells[J].J Clin Invest,2009,119(6):1417-1419.
[7] Acloque H, Adarns MS, Fishwick K,et al. Epithelial-mesenchymal transitions:the importance of changing cell state in development and disease[J].J Clin Invest,2009,119(6):1438-1449.
[8] Zeisberg M,Neilson EG.Biomarkers for epithelial-mesenchymal transitions[J].J Clin Invest,2009,119(6):1429-1437.
[9] Choi SS,Omenetti A,Witek RP, et al.Hedgehog pathway activation and epithelial-mesenchymal transitions during myofibroblastic transformation of rat hepatic cells in culture and cirrhosis[J]. Am J Physiol Gastrointest Liver Physiol,2009,297(6):G1093-G1096.
[10]Sicklick JK,Choi SS,Bustamante M,et al.Evidence for epithelial-mesenchymal transitions in adult liver cells[J]. Am J Physiol Gastrointest Liver Physiol, 2006,291(4):G575-G583.
[11]Dooley S, Hamzavi J, Ciuclan L, et al. Hepatocytespecific Smad7 expression attenuates TGF-β-mediated fibrogenesis and protects against liver damage [J]. Gastroenterology, 2008, 135(2):642-659.
[13]Aigner K,Dampier B,Descovich L,et al.The transcription factor ZEBl(δEFl) promotes tumour cell dedifferentiation by repressing master regulators of epithelial polarity[J].Oncogene,2007,26(49):6979-6988.
DynamicchangesofZEB1andZEB2duringdevelopmentofliverfibrosisinducedbychronicliverinjuryinrats
HAN Bing, XIE Ru-jia, HONG Qin, ZHANG Cheng-jun, YANG Qin, CHENG Ming-liang
(DepartmentofPathophysiology,GuiyangMedicalCollege,Guiyang550004,China.E-mail:hanbing791203@163.com)
AIM: To observe the dynamic changes of zinc finger E-box binding homeobox 1 (ZEB1) and zinc finger E-box binding homeobox 2 (ZEB2) during the development of liver fibrosis induced by chronic liver injury in rats.METHODSForty male Wistar rats weighing 180~220 g were divided intonormal control group and 4 model groups. The rats in model groups were induced by hypodermic injection of CCl4at a dose of 3 mL/kg and once per 3 days for 2 weeks, 4 weeks, 6 weeks and 8 weeks, respectively. The liver index and serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) were analyzed. The liver fibrosis was observed under microscope. Additionally, the expression of ZEB1, ZEB2, E-cadherin and α-smooth muscle actin(α-SMA) SMA at protein level was determined by immunohistochemistry. The mRNA expression of ZEB1 and ZEB2 was detected by real-time RT-PCR.RESULTSThe liver index and the serum levels of ALT and AST in the 4 model groups were obviously higher than those in normal control group. The apparent liver fibrosis was observed in 8-week model group. The protein levels of α-SMA (37.84±7.04), ZEB1 (46.42±14.36) and ZEB2(57.71±13.32), and mRNA expression of ZEB1 (189.00±47.39) and ZEB2 (277.28±48.55) in the livers of 8-week model rats were obviously higher than those in the control rats (P<0.01). The protein level of E-cadherin in the liver of 8-week model rats was obviously lower than that in the control rats (P<0.01).CONCLUSIONIn the process of liver fibrosis induced by CCl4, the obvious changes of ZEB1/ZEB2 may play an important role in chronic liver injury and liver fibrosis.
Liver fibrosis; Chronic liver injury; Zinc finger E-box binding homeobox protein
R363
A
10.3969/j.issn.1000-4718.2012.09.018
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
