siRNA抑制PPARγ表達對絨癌細胞JEG-3侵襲力的影響
2012-11-13 07:57:10陳麗君
中國病理生理雜志 2012年5期
關鍵詞:實驗
許 華, 陳麗君
滋養(yǎng)細胞浸潤是人類胎盤及胚胎發(fā)育的關鍵過程,其結果為胚胎植入母體組織,間質(zhì)成分以及母胎間血管浸潤。母體螺旋動脈周圍血管平滑肌的退化以及內(nèi)皮細胞被滋養(yǎng)細胞取代,最終可導致血管腔的擴張,增加絨毛間血流供應[1-2]。妊娠期母兒間特殊的血管聯(lián)系,使胎兒從母體中得到了持續(xù)的養(yǎng)分及血氧供應。滋養(yǎng)細胞侵襲能力不足與各種不同的病理妊娠有關。底蛻膜的淺浸潤以及螺旋動脈重鑄受阻,可導致子癇前期、胎兒生長受限及自然流產(chǎn)等病理妊娠的發(fā)生[3-5]。過氧化物酶體增殖物激活受體γ(peroxisome proliferator-activated receptor gamma,PPARγ)屬配體依賴的核激素受體超家族成員,廣泛存在于各種組織中,PPARγ處于多種信號轉錄途徑的交叉點,被配體激活后在轉錄水平上抑制多種細胞的增殖、侵襲、分化和凋亡,在滋養(yǎng)細胞中廣泛存在[6],由于JEG-3細胞株的生物學特性與細胞滋養(yǎng)層細胞(cytotrophoblasts)的特性非常相似,本實驗將PPARγsiRNA轉染JEG-3細胞,觀察其對JEG-3細胞侵襲能力的影響,從而探討子癇前期、胎兒生長受限及習慣性流產(chǎn)等病理妊娠的發(fā)病機制。
材料和方法
1 主要試劑
人絨毛膜上皮癌細胞JEG-3購自中科院上海生命科學院。RPMI-1640培養(yǎng)液(HyClone),胎牛血清(上海復蒙),siRNA(廣州銳博生物),逆轉錄試劑盒(TaKaRa),SYBR Premix TaqTM(GeneCopoeia),Trizol(TaKaRa),Transwell小室(膜孔徑 8μm)(Costar),Matrigel基質(zhì)膠(BD),轉染試劑 Lipofectamine 2000(Invitrogen),其它試劑均為國產(chǎn)分析純。
2 主要方法
2.1 細胞培養(yǎng) JEG-3細胞采用含有10%胎牛血清的RPMI-1640培養(yǎng)基培養(yǎng),置于溫度37℃、濕度95%、5%CO2的培養(yǎng)箱中培養(yǎng)。細胞經(jīng)0.25%含EDTA胰酶消化后,以5×105cells/well的密度接種于6孔板中,用于轉染及測定轉染效率,提取細胞中mRNA以及觀察細胞的侵襲能力。
2.2 siRNA的合成與轉染 根據(jù) GenBank中PPARγ的序列號,由廣州銳博生物技術公司設計合成siRNA,產(chǎn)品為3條對 PPARγ高效沉默的雙鏈siRNA,序列分別為:S1正義鏈5'-AGAUAAAGCUUCUGGAUUUdTdT -3',反義鏈 3'- dTdTUCUAUUUCGAAGACCUAAA-5',其針對靶序列為 AGATAAAGCTTCTGGATTT;S2正義鏈5'-AGGAAAGACAACAGACAAAdTdT-3',反義鏈 3'-dTdTUCCUUUCUGUUGUCUGUUU-5',其針對靶序列為 AGGAAAGCAAA;S3正義鏈5'-GUACCAAAGUGCAAUCAAAdTdT -3',反義鏈 3'- dTdTCAUGGUUUCACGUUAGUUU-5',的針對靶序列為 GTACCAAAGTGCAATCAAA。非特異性陰性對照(negative control)siRNA的序列不靶向沉默人類目前已知的任何基因,并且與PPARγ基因不具有同源性。帶有熒光標記的siRNA用于檢測轉染效率,本實驗中的陰性對照siRNA帶有熒光標記Cy3,最大發(fā)射波長為570 nm,熒光顯微鏡下觀察綠色光下發(fā)紅光。siRNA產(chǎn)品按說明書用滅菌ddH2O配制成20μmol/L溶液,分裝保存于-20℃中。常規(guī)培養(yǎng)JEG-3細胞后,將細胞按5×105cells/well密度接種至6孔板中,培養(yǎng)箱中孵育約24 h后,細胞生長融合度約為30% ~50%時,進行轉染。具體步驟按提供的說明書進行。
2.3 電子熒光顯微鏡觀察轉染效率 根據(jù)推薦濃度,分別以100、50 nmol/L的終濃度將帶有熒光標記的陰性對照siRNA(濃度過低轉染效率可能會低,濃度過高可能產(chǎn)生脫靶效應及增加細胞毒性),2μL Lipofectamine 2000轉染6孔板中的JEG-3細胞,轉染后6 h置于電子熒光顯微鏡下觀察轉染效率,實驗重復3次。經(jīng)過觀察,以終濃度為100 nmol/L時進行轉染效率最高,備下面繼續(xù)研究。
2.4 實時定量PCR技術檢測PPARγ及黏蛋白-1(mucin-1,MUC1)mRNA的表達 廣州銳博生物技術公司提供的PPARγ特異性siRNA共有3條鏈:S1、S2及S3,為測定這3條鏈對PPARγ基因的抑制率,設實驗(1)為S1組(轉染PPARγ特異性siRNA中的S1鏈)、S2組(轉染PPARγ特異性siRNA中的S2鏈)、S3組(轉染 PPARγ特異性 siRNA中的 S3鏈)、S4組(轉染混勻的PPARγ特異性siRNA中的S1、S2、S3鏈)和陰性對照組(轉染陰性對照 siRNA)。每組設3個復孔,轉染24 h后提取各組JEG-3細胞的總RNA,按RT試劑盒說明書反轉錄為cDNA后,應用實時定量PCR技術檢測PPARγmRNA,引物由廣州復能基因有限公司設計合成,PPARγ的引物序列為上游5'-GAACAGATCCAGTAATTGCAG -3',下游5'-AGGCTCTTCATGAGGCTTATTG -3',擴增長度為137 bp;管家基因β-actin引物序列為:上游5'- CCAACCGCGAGAAGATGA -3',下游 5'-CCAGAGGCGTACAGGGATAG -3',擴增片段長度為97 bp。反應體系總體積25μL,在LightCycler 2.0 real-time PCR擴增儀上擴增,退火溫度50℃ 20 s,40個循環(huán)。采用2-ΔΔCt相對定量法計算各組PPARγ表達量與β-actin表達量的比值,置空白對照組的值為1,換算出各組PPARγmRNA的表達水平。實驗重復3次,取其平均值。實驗證明,S2鏈siRNA對PPARγ的抑制效率最高,用于實驗組,以備后續(xù)實驗。
另設實驗(2)為實驗組(轉染PPARγsiRNA)、陰性對照組(轉染陰性對照siRNA)和空白對照組(不轉染任何siRNA,余試劑與其它2組相同),每組設3復孔,檢測PPARγ及MUC1 mRNA的表達。MUC1引物序列:上游5'-GGAAGACCCAGCACT-3',下游5'-CACAGATCC TGGCCTGAAC -3',按上述步驟進行,實驗重復3次,取其平均值。
2.5 Matrigel侵襲實驗檢測轉染后細胞的侵襲能力
取30μL按1∶5稀釋的Matrigel基質(zhì)膠包被Transwell小室(8μm孔徑)的上面,37℃培養(yǎng)箱中放置4 h。將轉染后24 h的各組細胞收集后,分別用含10%胎牛血清的RPMI-1640培養(yǎng)液重懸細胞,將細胞密度調(diào)至4×105/L,取100μL細胞懸液加入小室的上室,另將600μL含20%胎牛血清的 RPMI-1640培養(yǎng)液加入小室的下室,置培養(yǎng)箱中繼續(xù)培養(yǎng)24 h取出Transwell小室,用棉簽小心拭去濾膜上室面的細胞,常規(guī)HE染色,取下濾膜,粘于載玻片上,于倒置顯微鏡的高倍顯微鏡(×400)下隨機選擇5個視野,分別計數(shù)細胞數(shù)并取其平均值,以穿膜細胞數(shù)表示細胞的侵襲能力。實驗重復3次,結果取其平均值。
3 統(tǒng)計學處理
采用SPSS 17.0軟件進行統(tǒng)計分析,數(shù)據(jù)用均數(shù)±標準差(ˉx±s)表示。組間差異用單因素方差分析進行統(tǒng)計,以P<0.05為差異有統(tǒng)計學意義。
結 果
1 JEG-3細胞的siRNA的轉染效率
熒光標記siRNA波長為470 nm,在綠色光下發(fā)紅色熒光,以100 nmol/L的siRNA進行轉染,其轉染效率最高,見圖1。
Figure 1.Results of JEG-3 cells transfected by siRNA labelled by fluorescence after 6 h by fluorescence microscopy( ×400).A:the final concentration of siRNA was 100 nmol/L;B:the final concentration of siRNA was 50 nmol/L.圖1 熒光顯微鏡下觀察熒光標記的siRNA轉染JEG-3細胞6 h
2 各組轉染后細胞中PPARγ及MUC1 mRNA的表達情況
轉染PPARγsiRNA后,S1組與空白對照組相比,PPARγmRNA 的表達水平下調(diào)了(40.88±1.05)%(P<0.05),S2組與空白對照組相比,下調(diào)了(71.92±0.71)%(P <0.05),S3 組與空白對照組相比,下調(diào)了(62.69±0.66)%(P <0.05),S4組與空白對照組相比,下調(diào)了(52.80±0.52)%(P<0.05)。其中,S2組siRNA抑制效率最高,見圖2。
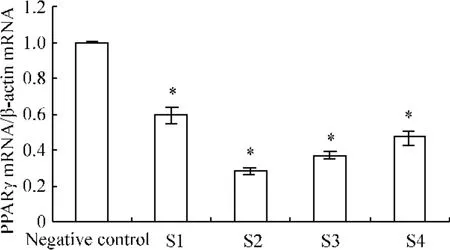
Figure 2.Relative expression of PPARγmRNA in JEG-3 cells after transfection.ˉx±s.n=15.*P<0.05 vs negative control.圖2 實時定量PCR測定各組細胞轉染后PPARγmRNA的相對表達量
轉染PPARγsiRNA后,實驗組細胞中 PPARγ mRNA的表達水平與空白對照組相比下調(diào)了(75.0±0.8)%(P<0.05),陰性對照組與空白對照組則無顯著差異(P>0.05),見圖3。實驗組細胞中MUC1 mRNA的表達水平與空白組相比下調(diào)了(65.0±1.3)%(P<0.05),陰性對照組與空白對照組則無顯著差異(P>0.05),見圖4。
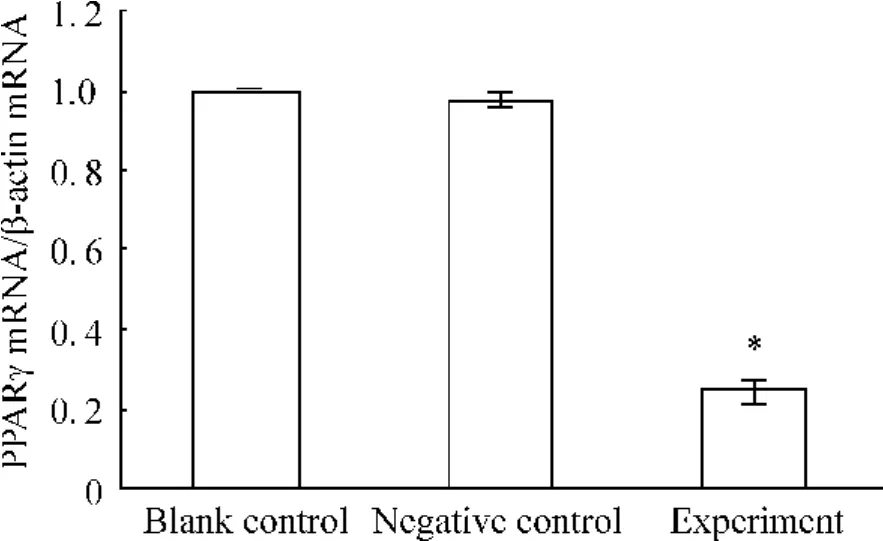
Figure 3.Relative expression of PPARγmRNA in JEG-3 cells after transfection.ˉx±s.n=9.*P<0.05 vs other groups.圖3 各組細胞轉染后PPARγmRNA相對表達量
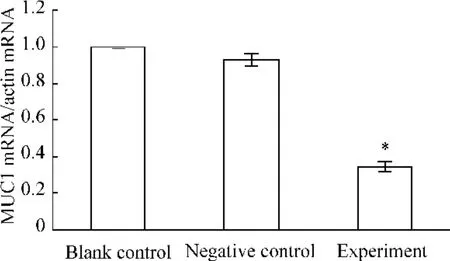
Figure 4.Relative expression of MUC1 mRNA in JEG-3 cells after transfection.ˉx±s.n=9.*P<0.05 vs other groups.圖4 各組細胞轉染后MUC1 mRNA相對表達量
3 各組轉染后JEG-3細胞侵襲力的比較
高倍顯微鏡(×400)下,實驗組穿膜細胞數(shù)為(47±4)個,陰性對照組的穿膜細胞數(shù)為(25±3)個,空白對照組的穿膜細胞數(shù)為(23±3)個,實驗組穿膜細胞數(shù)與空白對照組的穿膜細胞數(shù)有顯著差異(P<0.05),說明轉染PPARγsiRNA后,細胞的侵襲力增強。陰性對照組與空白對照組的穿膜細胞數(shù)相比無顯著差異(P <0.05),見圖5、6。
Figure 5.The invasion of JEG -3 cells24 h after transfection( ×400).A:blank control group;B:negative control group;C:experiment group.圖5 轉染24 h后各組JEG-3細胞的侵襲力
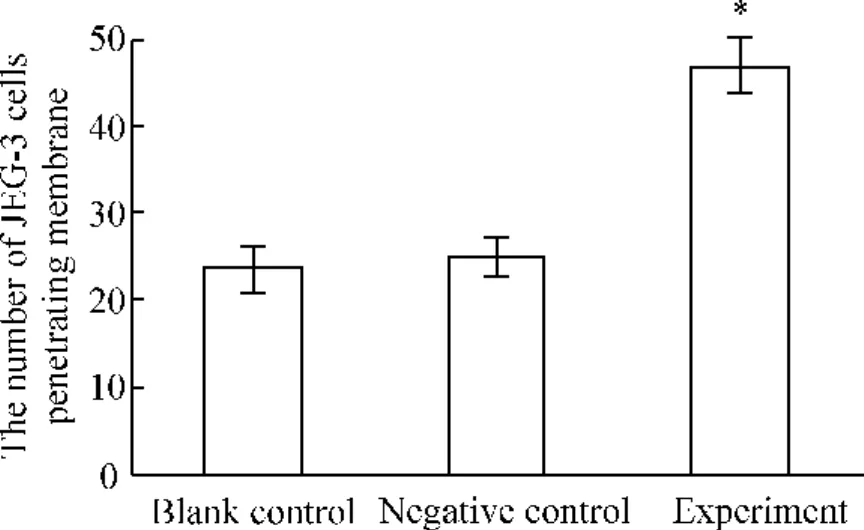
Figure 6.The number of JEG-3 cells penetrating membrane 24 h after transfection.ˉx±s.n=9.*P<0.05 vs other groups.圖6 轉染24 h后各組JEG-3細胞的穿膜細胞數(shù)
討 論
人類胚胎的著床、胎盤的形成以及胚胎的生長發(fā)育與滋養(yǎng)細胞的增殖、分化及侵襲功能有密切關系,受一系列復雜的、彼此聯(lián)系的事件精密、嚴格地調(diào)控,其中滋養(yǎng)細胞對子宮內(nèi)膜的合理侵入,以及著床后滋養(yǎng)細胞對子宮蛻膜層和肌層血管的侵襲,使子宮螺旋動脈發(fā)生血管重鑄等生理現(xiàn)象,是決定妊娠正常與否的關鍵,并維持妊娠的正常進行。滋養(yǎng)細胞的侵襲能力受多種因素調(diào)控[7]:滋養(yǎng)細胞分泌合成一些細胞因子及生長因子通過誘導內(nèi)皮細胞及血管平滑肌細胞的凋亡調(diào)節(jié)血管重鑄過程、子宮螺旋動脈血流動力學、細胞外基質(zhì)成分的改變、血管平滑肌細胞特性的變化以及局部免疫環(huán)境的變化。當這些因素平衡失調(diào)時,滋養(yǎng)細胞侵襲能力不足,子宮螺旋動脈血管重鑄受阻,可能導致子癇前期、胎兒生長受限及自然流產(chǎn)的發(fā)生。目前為止,已有許多研究發(fā)現(xiàn)多種影響滋養(yǎng)細胞侵襲的因素及其在子癇前期等病理妊娠中的發(fā)病機制,探討各種因子在滋養(yǎng)細胞侵襲能力的信號轉導途徑及其作用已成為目前病理妊娠研究的熱點。
PPARs屬于Ⅱ型核受體超家族成員,是近年來發(fā)現(xiàn)的一類核激素受體超家族成員。目前,已發(fā)現(xiàn)3種不同的PPARs亞型,分別為PPARα、PPARβ及PPARγ,這3種亞型在結構及功能上各有差異。其中,PPARγ有6個區(qū)域(A~F),4個功能結構域:N末端的A/B結構區(qū)為非配體依賴性的轉錄活化區(qū)域,可通過MAPK途徑被磷酸化,影響轉錄活性以及配基與受體結合;DNA結合結構域C區(qū)與啟動子和目的基因結合,調(diào)節(jié)基因轉錄;轉錄活性調(diào)節(jié)結構域D區(qū),是阻遏劑結合位點,許多核內(nèi)因子與該結構域結合影響PPARγ活性;配體結合域E區(qū);F為配體依賴性活化區(qū)。PPARγ須依賴于與其配體相結合,PPARγ被激活后與視磺酸受體(retinoic X receptor,RXR)結合形成異二聚體,再與過氧化物酶體增殖物反應元件(peroxisome proliferator responsive element,PPRE)結合,激活靶基因,從而調(diào)節(jié)基因轉錄、翻譯及各種生物學效應[8-9]。PPARγ 處于各種信號轉導途徑的交叉點,參與了糖、脂肪代謝、單核細胞激活、炎癥反應、細胞的分化、增生及凋亡等生理病理過程。其生物學功能復雜多樣,須與相應配體結合后才能活化,產(chǎn)生生物學效應。
迄今,已有很多關于PPARγ與滋養(yǎng)細胞侵襲力方面的研究。Barak等[10]最早發(fā)現(xiàn)PPARγ在胎盤發(fā)育中的作用,在其研究中發(fā)現(xiàn)PPARγ基因敲除影響了孕鼠胎盤滋養(yǎng)細胞的終末分化以及胎盤血管發(fā)生變化。Asami-Miyagishi等[11]采用 RT-PCR 方法檢測到在小鼠孕11 d時胎盤中即有PPARγmRNA的表達,在孕13 d時表達升高,之后則表達下降。免疫組化方法發(fā)現(xiàn)在孕鼠胎盤的母胎界面有PPARγ及其天然配體15-d-PGJ2的表達,說明PPARγ參與了胎盤的發(fā)育、發(fā)展過程。Fournier等[12]通過建立體外滋養(yǎng)細胞模型,用PPARγ的激活物抑制滋養(yǎng)細胞的侵襲力,并且檢測到轉化生長因子β(transforming growth factorβ,TGF-β)和妊娠相關血漿蛋白A(pregnancy-associated plasma protein-A,PAPP-A)的下調(diào)及白細胞介素-1β(interleukin-1β,IL-1β)的上調(diào),因為這些因子可能與滋養(yǎng)細胞的侵襲有關,推測PPARγ可能通過以上途徑影響滋養(yǎng)細胞侵襲能力。李淑娟等[13]利用早孕絨毛分離培養(yǎng)絨毛外滋養(yǎng)細胞模型,發(fā)現(xiàn)PPARγ的上調(diào)抑制了基質(zhì)金屬蛋白酶2、9(matrix metalloproteinase-2,9,MMP-2和 MMP-9)的表達,PPARγ抑制滋養(yǎng)細胞侵襲力可能與MMP-2、9有關。這些研究都表明PPARγ通過不同途徑影響滋養(yǎng)細胞的侵襲能力。Shalom-Barak等[14]通過實驗研究顯示MUC1基因是 PPARγ的靶基因,PPARγ缺失的胎盤組織中MUC1的表達也缺失,PPARγ激動劑羅格列酮(rosiglitazone)可上調(diào)MUC1基因的表達,推斷PPARγ可能通過調(diào)節(jié)MUC1調(diào)節(jié)胎盤的發(fā)育,另有研究發(fā)現(xiàn),Shyu等[15]通過體外培養(yǎng)滋養(yǎng)細胞模型,發(fā)現(xiàn)MUC1的過表達能夠下調(diào)MMP-9的表達,并抑制滋養(yǎng)細胞的侵襲能力。
因此本實驗設計采用siRNA技術,能夠特異性地抑制JEG-3細胞中PPARγ的表達,其mRNA抑制效率達(75.0±0.8)%,同時能夠下調(diào)黏蛋白 MUC1 mRNA水平的表達(65.0±1.3)%。Transwell侵襲實驗結果顯示PPARγ的下調(diào)能夠使滋養(yǎng)細胞的侵襲能力增強,這與之前的研究結果相一致,通過體外培養(yǎng)滋養(yǎng)細胞實驗再一次驗證了PPARγ能通過調(diào)節(jié)MUC1的表達影響滋養(yǎng)細胞的侵襲能力。
目前,國內(nèi)外還沒有關于通過抑制PPARγ下調(diào)滋養(yǎng)細胞中MUC1的表達影響滋養(yǎng)細胞侵襲力的研究,本研究驗證了這一點。但是PPARγ對MUC1的具體調(diào)節(jié)機制目前還不清楚,尚待進一步研究。這些研究將為探索滋養(yǎng)細胞侵襲能力不足引起的子癇前期、胎兒生長受限及自然流產(chǎn)等病理妊娠的發(fā)病機制提供依據(jù)。
[1] Harris LK,Keogh RJ,Wareing M,et al.Invasive trophoblasts stimulate vascular smooth muscle cell apoptosis by a Fas ligand-dependent mechanism[J].Am JPathol,2006,169(5):1863-1874.
[2] Pijnenborg R,Bland JM,Robertson WB,et al.Uteroplacental arterial changes related to interstitial trophoblast migration in early human pregnancy[J].Placenta,1983,4(4):397-413.
[3] Khong TY,De Wolf F,Robertson WB,et al.Inadequate maternal vascular response to placentation in pregnancies complicated by pre-eclampsia and by small-for-gestational age infants[J].Br J Obstet Gynaecol,1986,93(10):1049-1059.
[4] Zhou Y,Damsky CH,F(xiàn)isher SJ.Preeclampsia is associated with failure of human cytotrophoblasts to mimic a vascular adhesion phenotype.One cause of defective endovascular invasion in this syndrome? [J].JClin Invest,1997,99(9):2152-2164.
[5] Toth B,Bastug M,Mylonas I,et al.Peroxisome proliferator-activated receptor-gamma in normal human pregnancy and miscarriage[J].Acta Histochemica,2009,111(4):373-379.
[6] 李淑娟,尚 濤,李思揚,等.過氧化物酶體增殖型活化受體γ及其配體對早孕期細胞滋養(yǎng)細胞浸潤能力的影響及機制研究[J].中華醫(yī)學雜志,2007,87(3):174-178.
[7] Whitley GS,Cartwright JE.Cellular and molecular regulation of spiral artery remodelling:lessons from the cardiovascular field[J].Placenta,2010,31(6):465 -474.
[8] Vamecq J,Latruffe N.Medical significance of peroxisome proliferator- activated receptors[J].Lancet,1999,354(9173):141-148.
[9] Willson TM,Brown PJ,Sternbach DD,et al.The PPARs:from orphan receptors to drug discovery[J].J Med Chem,2000,43(4):527-550.
[10] Barak Y,Nelson MC,Ong ES,et al.PPARγ is required for placental ,cardiac,and adipose tissue development[J].Mol Cell,1999,4(4):585 -595.
[11] Asami-Miyagishi R,Iseki S,Usui M,et al.Expression and function of PPARγ in rat placental development[J].Biochem Biophys Res Commun,2004,315(2):497-501.
[12] Fournier T ,Handschuh K,Tsatsaris V,et al.Involvement of PPARγ in human trophoblast invasion[J].Placenta,2007,28(Suppl A):S76-S81.
[13] 李淑娟,尚 濤,李秋玲,等.過氧化物酶體增殖物激活型受體γ及其配體對早期細胞滋養(yǎng)細胞MMP-2、MMP-9表達的調(diào)控[J].生殖與避孕,2007,27(2):107-112.
[14] Shalom -Barak T,Nicholas JM,Wang Y,et al.Peroxisome proliferator-activated receptor gamma controls Muc1 transcription in trophoblasts[J].Mol Cell Biol,2004,24(24):10661-10669.
[15] Shyu MK,Lin MC,Liu CH,et al.MUC1 expression is increased during human placental development and suppresses trophoblast-like cell invasion in vitro[J].Biol Reprod,2008,79(2):233 -239.
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數(shù)理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發(fā)明與創(chuàng)新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
